α-Lipoic Acid Reduces Hypertension and Increases Baroreflex Sensitivity in Renovascular Hypertensive Rats

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
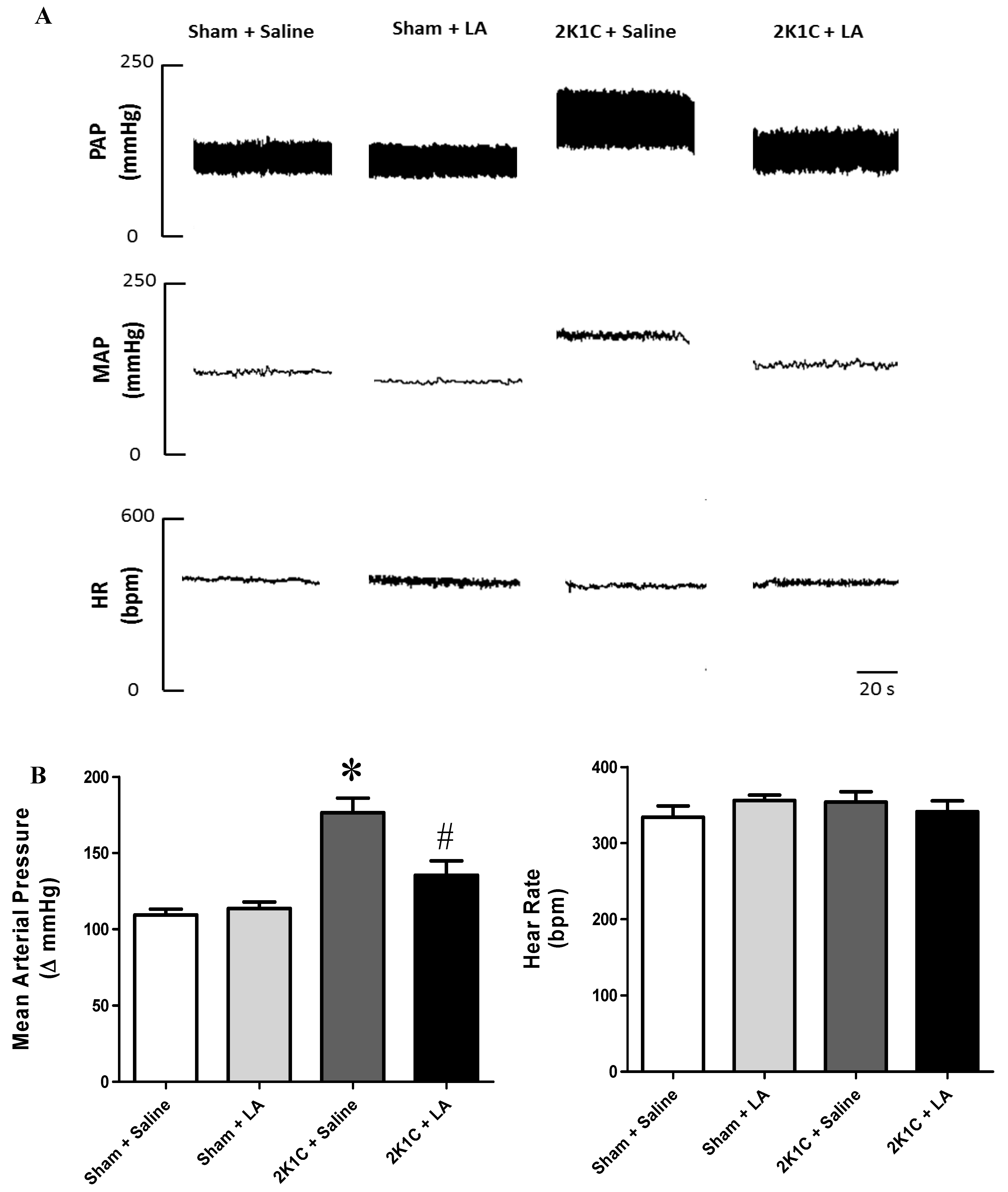
2.1. Renal Artery Clipping Induces Hypertension without Affecting Heart Rate

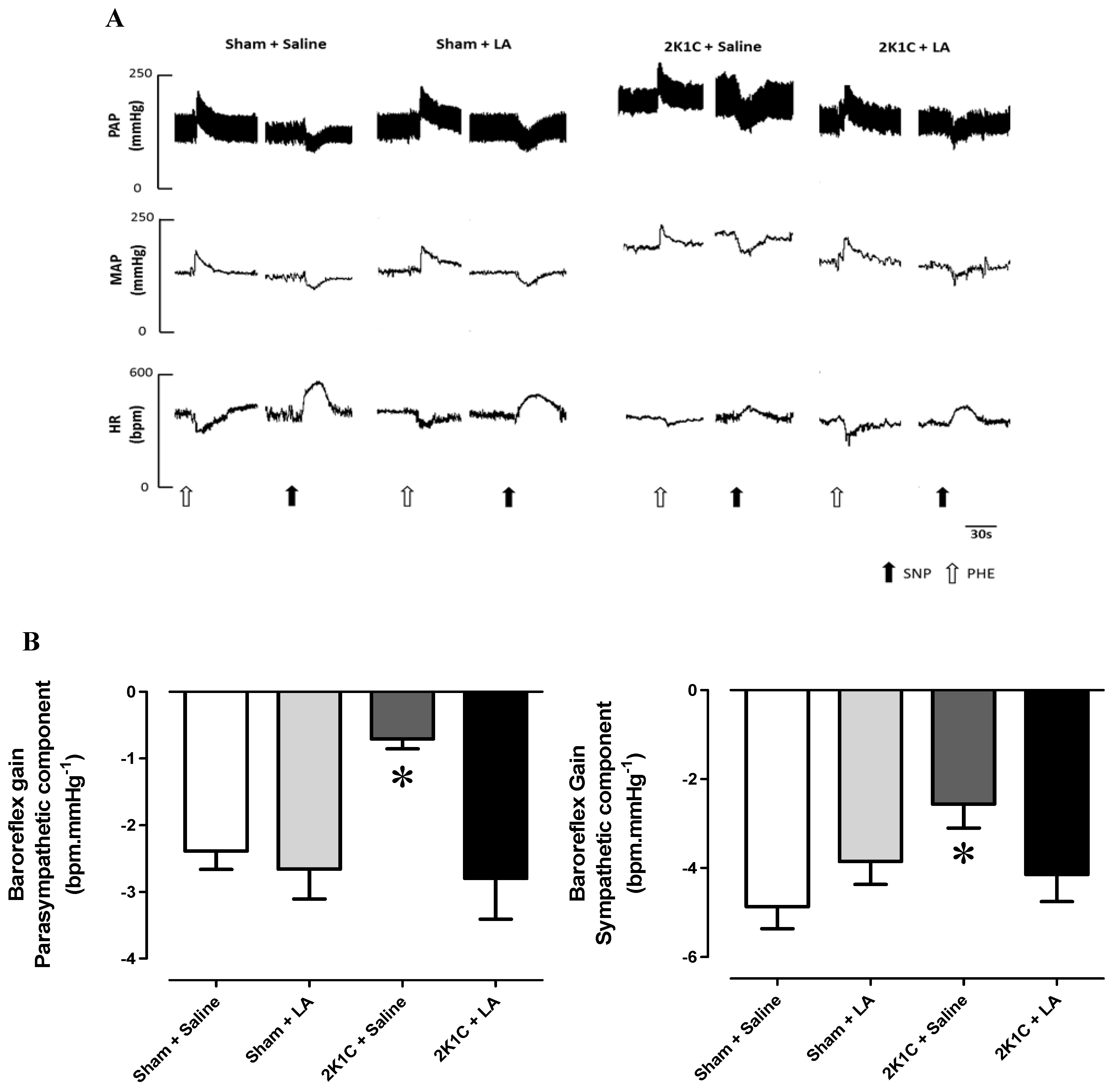
2.2. Chronic Administration of LA Restores Baroreflex Sensitivity in Renovascular Hypertension

3. Experimental
3.1. Animals
3.2. Surgical Procedures
3.3. Blood Pressure and Heart Rate Recordings
3.4. Baroreflex Sensitivity Test
3.5. Statistical Analyses
4. Conclusions
Acknowledgments
- Sample Availability: Samples of the compounds are available from the authors.
References
- Reed, L. Multienzyme complexes. Acc. Chem. Res. 1974, 7, 40–46. [Google Scholar] [CrossRef]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar]
- Takaoka, M.; Kobayashi, Y.; Yuba, M.; Ohkita, M.; Matsumura, Y. Effects of α-lipoic acid on deoxycorticosterone acetate-salt-induced hypertension in rats. Eur. J. Pharm. 2001, 424, 121–129. [Google Scholar] [CrossRef]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Effects of α-lipoic acid on biomarkers of oxidative stress in streptozotocin-induced diabetic rats. J. Nutr. Biochem. 2003, 14, 288–294. [Google Scholar] [CrossRef]
- Biewenga, G.P.; Haenen, G.R.M.; Bast, A. The pharmacology of the antioxidant lipoic acid. Gen. Pharm. 1997, 29, 315–331. [Google Scholar] [CrossRef]
- Heinisch, B.B.; Francesconi, M.; Mittermayer, F.; Schaller, G.; Gouya, G.; Wolzt, M.; Pleiner, J. Alpha-lipoic acid improves vascular endothelial function in patients with type 2 diabetes: A placebo-controlled randomized trial. Eur. J. Clin. Investig. 2010, 40, 148–154. [Google Scholar] [CrossRef]
- Braga, V.A.; Antunes, V.R.; Machado, B.H. Autonomic and respiratory responses to microinjection of L-glutamate into the commissural subnucleus of the NTS in the working heart-brainstem preparation of the rat. Brain Res. 2006, 1093, 150–160. [Google Scholar] [CrossRef]
- Simic, D.V.; Mimic-Oka, J.; Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Opacic, M.; Matic, D.; Ianovic, B.; Simic, T. Byproducts of oxidative protein damage and antioxidant enzyme activities in plasma of patients with different degrees of essential hypertension. J. Hum. Hypertens. 2006, 20, 149–155. [Google Scholar] [CrossRef]
- Matteucci, E.; Giampietro, O. Thiol Signalling Network with an Eye to Diabetes. Molecules 2010, 15, 8890–8903. [Google Scholar] [CrossRef]
- Kitiyakara, C.; Wilcox, C.S. Antioxidants for hypertension. Curr. Opin. Nephrol. Hypertens. 1998, 7, 531–538. [Google Scholar] [CrossRef]
- Houston, M.C. Nutraceuticals, vitamins, antioxidants, and minerals in the prevention and treatment of hypertension. Prog. Cardiovasc. Dis. 2005, 47, 396–449. [Google Scholar] [CrossRef]
- Braga, V.A.; Burmeister, M.A.; Zhou, Y.; Sharma, R.V.; Davisson, R.L. Selective ablation of AT1a receptors in rostral ventrolateral medulla (RVLM) prevents chronic angiotensin-II-dependent hypertension in part by reducing oxidant stress in this region. Hypertension 2008, 52, e36. [Google Scholar]
- Botelho-Ono, M.S.; Pina, H.V.; Sousa, K.H.F.; Nunes, F.C.; Medeiros, I.A.; Braga, V.A. Acute superoxide scavenging restores depressed baroreflex sensitivity in renovascular hypertensive rats. Auton. Neurosci. 2011, 159, 38–44. [Google Scholar] [CrossRef]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef]
- Rajagopalan, S.; Kurz, S.; Munzel, T.; Tarpey, M.; Freeman, B.A.; Griendling, K.K.; Harrison, D.G. Angiotensin II-mediated hypertension in the rat increases vascular superoxide production via membrane NADH/NAD(P)H oxidase activation: Contribution to alterations of vasomotor tone. J. Clin. Investig. 1996, 97, 1916–1923. [Google Scholar] [CrossRef]
- American Heart Association. Heart disease and stroke statistics—2008 update. In Circulation; 2008; 117, pp. 125–146. [Google Scholar]
- Lefkowits, R.J.; Willerson, J. Prospects for cardiovascular research. JAMA 2001, 285, 581–587. [Google Scholar] [CrossRef]
- Queiroz, T.M.; Machado, N.T.; Furtado, F.F.; Oliveira-Filho, A.A.; Alustau, M.C.; Figueiredo, C.S.; Miranda, G.E.C.; Barbosa-Filho, J.M.; Braga, V.A.; Medeiros, I.A. Vasorelaxation induced by Dictyota pulchella (Dictyotaceae), a brown alga, is mediated via inhibition of calcium influx in rats. Mar. Drugs 2011, 9, 2075–2088. [Google Scholar] [CrossRef]
- Chen, X.; Touyz, R.M.; Park, J.B.; Schiffrin, E.L. Antioxidant effects of vitamins C and E are associated with altered activation of vascular NADPH oxidase and superoxide dismutase in stroke-prone SHR. Hypertension 2001, 38, 606–611. [Google Scholar] [CrossRef]
- Hong, H.J.; Hsiao, G.; Cheng, T.H.; Yen, M.H. Supplementation with tetrahydrobiopterin suppresses the development of hypertension in spontaneously hypertensive rats. Hypertension 2001, 38, 1044–1048. [Google Scholar] [CrossRef]
- Costa, C.A.; Amaral, T.A.; Carvalho, L.C.; Ognibene, D.T.; da Silva, A.F.; Moss, M.B.; Valença, S.S.; de Moura, R.S.; Resende, A.C. Antioxidant treatment with tempol and apocynin prevents endothelial dysfunction and development of renovascular hypertension. Am. J. Hypertens. 2009, 22, 1242–1249. [Google Scholar] [CrossRef]
- Landmesser, U.; Dikalov, S.; Price, S.R.; Mc-Cann, L.; Fukai, T.; Holland, S.M.; Mitch, W.E.; Harrison, D.G. Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J. Clin. Investig. 2003, 111, 1201–1209. [Google Scholar]
- Grassi, G.; Cattaneo, B.M.; Seravalle, G.; Lanfranchi, A.; Mancia, G. Baroreflex control of sympathetic nerve activity in essential and secondary hypertension. Hypertension 1998, 31, 68–72. [Google Scholar] [CrossRef]
- Salgado, M.C.; Justo, S.V.; Joaquim, L.F.; Fazan, R., Jr.; Salgado, H.C. Role of nitric oxide and prostanoids in attenuation of rapid baroreceptor resetting. Am. J. Physiol. Heart Circ. Physiol. 2009, 290, 1059–1063. [Google Scholar]
- Pechánová, O.; Zichaj, J.; Kojsová, S.; Dobesová, Z.; Jendekovál, L.; Kunes, J. Effect of chronic N-acetylcysteine treatment on the development of spontaneous hypertension. Clin. Sci. 2006, 110, 235–242. [Google Scholar] [CrossRef]
- Contreras, F.; de La Parte, M.A.; Cabrera, J.; Ospino, N.; Israili, Z.H.; Velasco, M. Role of angiotensin II AT1 receptor blockers in the treatment of arterial hypertension. Am. J. Ther. 2003, 10, 401–408. [Google Scholar] [CrossRef]
- Berk, B.C.; Vekshtein, V.; Gordon, H.M.; Tsuda, T. Angiotensin II-stimulated protein synthesis in cultured vascular smooth muscle cells. Hypertension 1989, 13, 305–314. [Google Scholar] [CrossRef]
- Geisterfer, A.A.; Peach, M.J.; Owens, G.K. Angiotensin II induces hypertrophy, not hyperplasia, of cultured rat aortic smooth muscle cells. Circ. Res. 1988, 62, 749–756. [Google Scholar] [CrossRef]
- Naftilan, A.J.; Gilliland, G.K.; Eldridge, C.S.; Kraft, A.S. Induction of the proto oncogene c-jun by angiotensin II. Mol. Cell. Biol. 1990, 10, 5536–5540. [Google Scholar]
- Neyses, L.; Nouskas, J.; Luyken, J.; Fronhoffs, S.; Oberdorf, S.; Pfiefer, U.; Williams, R.S.; Sukhatme, V.P.; Vetter, H. Induction of immediateearly genes by angiotensin II and endothelin-1 in adult rat cardiomyocytes. J. Hypertens. 1993, 11, 927–934. [Google Scholar] [CrossRef]
- Taubman, M.B.; Berk, B.C.; Izumo, S.; Tsuda, T.; Alexander, R.W.; Nadal-Ginard, B. Angiotensin II induces c-fos mRNA in aortic smooth muscle. Role of Ca2+ mobilization and protein kinase C activation. J. Biol. Chem. 1989, 264, 526–530. [Google Scholar]
- Everett, A.D.; Tufro-McReddie, A.; Fisher, A.; Gomez, R.A. Angiotensin receptor regulates cardiac hypertrophy and transforming growth factor-beta 1 expression. Hypertension 1994, 23, 587–592. [Google Scholar] [CrossRef]
- Fernandez-Alfonso, M.S.; Ganten, D.; Paul, M. Mechanisms of cardiac growth. The role of the renin-angiotensin system. Basic Res. Cardiol. 1992, 87, 173–181. [Google Scholar] [CrossRef]
- Himeno, H.; Crawford, D.C.; Hosoi, M.; Chobanian, A.V.; Brecher, P. Angiotensin II alters aortic fibronectin independently of hypertension. Hypertension 1994, 23, 823–826. [Google Scholar] [CrossRef]
- Crawford, D.C.; Chobanian, A.V.; Brecher, P. Angiotensin II induces fibronectin expression associated with cardiac fibrosis in the rat. Circ. Res. 1994, 74, 727–739. [Google Scholar] [CrossRef]
- Berry, C.; Hamilton, C.A.; Brosnan, M.J.; Magill, F.G.; Berg, G.A.; McMurray, J.J.; Dominiczak, A.F. Investigation into the sources of superoxide in human blood vessels: Angiotensin II increases superoxide production in human internal mammary arteries. Circulation 2000, 101, 2206–2212. [Google Scholar] [CrossRef]
- Zimmerman, M.C.; Lazartigues, E.; Sharma, R.V.; Davisson, R.L. Hypertension caused by angiotensin II infusion involves increased superoxide production in the central nervous system. Circ. Res. 2004, 95, 210–216. [Google Scholar] [CrossRef]
- Lassegue, B.; Clempus, R.E. Vascular NAD(P)H oxidases: Specific features, expression, and regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, 277–297. [Google Scholar]
- Hirooka, Y. Role of reactive oxygen species in brainstem in neural mechanisms of hypertension. Auton. Neurosci. 2008, 142, 20–24. [Google Scholar] [CrossRef]
- Monahan, K.D.; Eskurza, I.; Seals, D.R. Ascorbic acid increases cardiovagal baroreflex sensitivity in healthy older men. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, 2113–2117. [Google Scholar] [CrossRef]
- Nishi, E.E.; Oliveira-Sales, E.B.; Bergamaschi, C.T.; Oliveira, T.G.; Boim, M.A.; Campos, R.R. Chronic antioxidant treatment improves arterial renovascular hypertension and oxidative stress markers in the kidney in Wistar rats. Am. J. Hypertens. 2010, 23, 473–480. [Google Scholar] [CrossRef]
- Li, Z.; Mao, H.Z.; Abboudm, F.M.; Chapleau, M.W. Oxygen-derived free radicals contribute to baroreceptor dysfunction in atherosclerotic rabbits. Circ. Res. 1996, 79, 802–811. [Google Scholar] [CrossRef]
- Nightingale, A.K.; Blackman, D.J.; Field, R.; Glover, N.J.; Pegge, N.; Mumford, C.; Schmitt, M.; Ellis, G.R.; Morris-Thurgood, J.A.; Frenneaux, M.P. Role of nitric oxide and oxidative stress in baroreceptor dysfunction in patients with chronic heart failure. Clin. Sci. 2003, 104, 529–535. [Google Scholar] [CrossRef]
- Chapleau, M.W.; Cunningham, J.T.; Sullivan, M.J.; Wachtel, R.E.; Abboud, F.M. Structural versus functional modulation of the arterial baroreflex. Hypertension 1995, 26, 341–347. [Google Scholar] [CrossRef]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Is alpha-lipoic acid a scavenger of reactive oxygen species in vivo? Evidence for its initiation of stress signaling pathways that promote endogenous antioxidant capacity. IUBMB Life 2008, 60, 362–367. [Google Scholar] [CrossRef]
- San Jose, G.; Fortuno, A.; Beloqui, O.; Diez, J.; Zalba, G. NADPH oxidase CYBA polymorphisms oxidative stress and cardiovascular diseases. Clin. Sci. (Lond.) 2008, 114, 173–182. [Google Scholar] [CrossRef]
- Judge, E.P.; Phelan, D.; O’Shea, D. Beyond statin therapy: A review of the management of residualrisk in diabetes mellitus. J. R. Soc. Med. 2010, 103, 357–362. [Google Scholar] [CrossRef]
- Galan, C.; Jardín, I.; Dionisio, N.; Salido, G.; Rosado, J.A. Role of Oxidant Scavengers in the Prevention of Ca2+ Homeostasis Disorders. Molecules 2010, 15, 7167–7187. [Google Scholar] [CrossRef]
- Braga, V.A. Dietary salt enhances angiotensin-II-induced superoxide formation in the rostral ventrolateral medulla. Auton. Neurosci. Basic Clin. 2010, 155, 14–18. [Google Scholar] [CrossRef]
- Braga, V.A.; Burmeister, M.A.; Sharma, R.V.; Davisson, R.L. Cardiovascular responses to peripheral chemoreflex activation and comparison of different methods to evaluate baroreflex gain in conscious mice using telemetry. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, 1168–1174. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Queiroz, T.M.; Guimarães, D.D.; Mendes-Junior, L.G.; Braga, V.A. α-Lipoic Acid Reduces Hypertension and Increases Baroreflex Sensitivity in Renovascular Hypertensive Rats. Molecules 2012, 17, 13357-13367. https://doi.org/10.3390/molecules171113357
Queiroz TM, Guimarães DD, Mendes-Junior LG, Braga VA. α-Lipoic Acid Reduces Hypertension and Increases Baroreflex Sensitivity in Renovascular Hypertensive Rats. Molecules. 2012; 17(11):13357-13367. https://doi.org/10.3390/molecules171113357
Chicago/Turabian StyleQueiroz, Thyago M., Drielle D. Guimarães, Leônidas G. Mendes-Junior, and Valdir A. Braga. 2012. "α-Lipoic Acid Reduces Hypertension and Increases Baroreflex Sensitivity in Renovascular Hypertensive Rats" Molecules 17, no. 11: 13357-13367. https://doi.org/10.3390/molecules171113357
APA StyleQueiroz, T. M., Guimarães, D. D., Mendes-Junior, L. G., & Braga, V. A. (2012). α-Lipoic Acid Reduces Hypertension and Increases Baroreflex Sensitivity in Renovascular Hypertensive Rats. Molecules, 17(11), 13357-13367. https://doi.org/10.3390/molecules171113357