Antioxidant, Antitubercular and Cytotoxic Activities of Piper imperiale

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolic Content

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. imperiale | Phenols (mg GAE/g DW) |
|---|---|
| Leaf | 6.94 ± 0.4 |
| Flower | 2.86 ± 0.2 |
| Wood | 1.98 ± 0.2 |
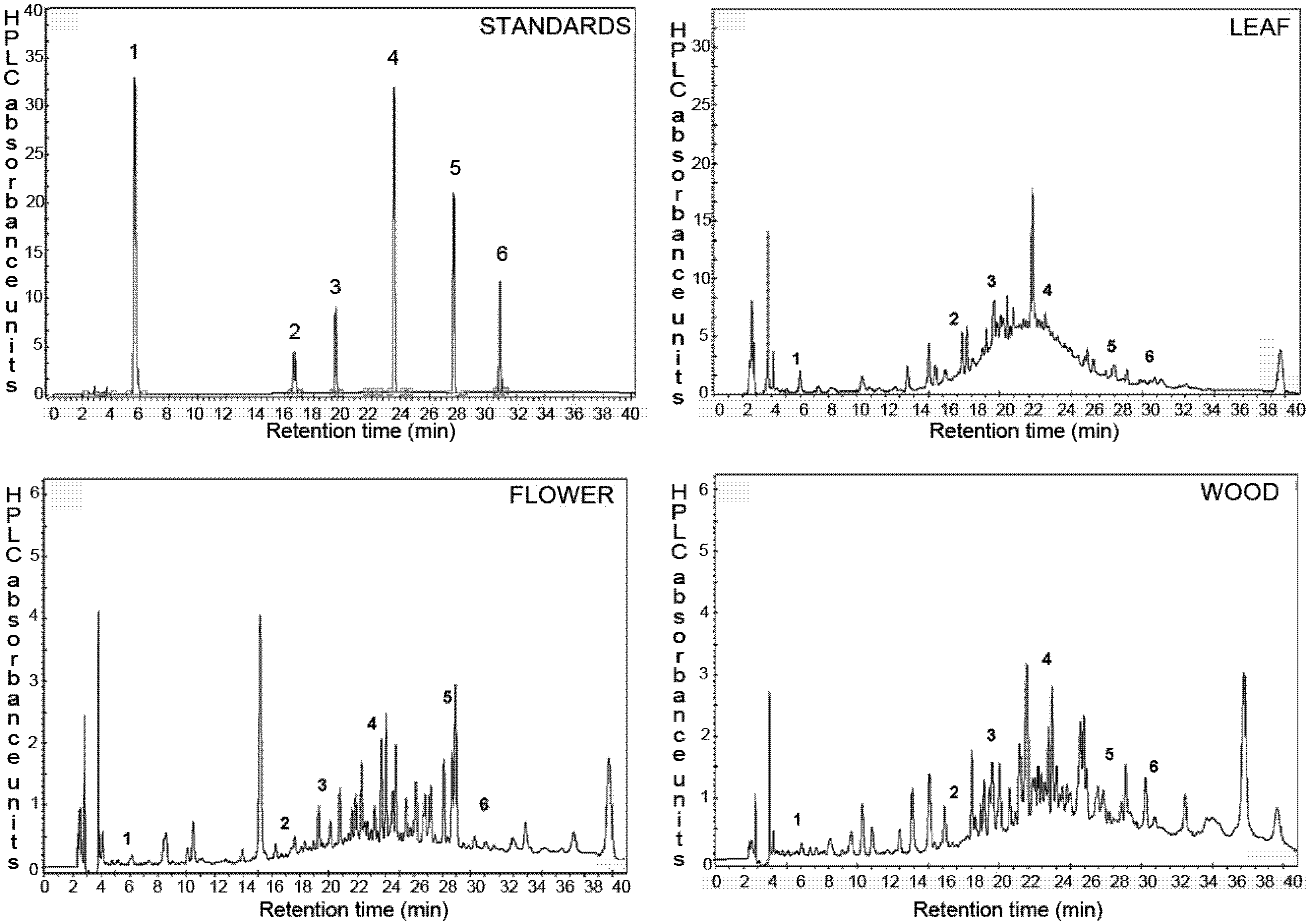
2.2. Identification and Quantification of Phenolic Compounds by HPLC
| No. | Compound | Natural Product Class | Content in Extracts (µg/100 g DW) | ||
|---|---|---|---|---|---|
| Leaves | Flowers | Wood | |||
| 1 | Gallic acid | Phenolic acid | 939.2 ± 74.2 | 13.1 ± 0.17 | 33.5 ± 0.43 |
| 2 | Catechin | Flavanol | 13613 ± 254 | 379.4 ± 15.4 | 457.1 ± 55.1 |
| 3 | Epicatechin | Flavanol | 2378 ± 41.7 | 1124 ± 5.30 | 1950 ± 17.7 |
| 4 | Ferulic acid | Hydroxycinnamic acid | 267.6 ± 8.48 | 75.0 ± 0.07 | 1629 ± 2.94 |
| 5 | Resveratrol | Stilbene | 1133 ± 28.3 | 1076 ± 201 | 97.8 ± 4.60 |
| 6 | Quercetin | Flavonol | 1954 ± 42.4 | 3661 ± 299 | 572.1 ± 16.8 |

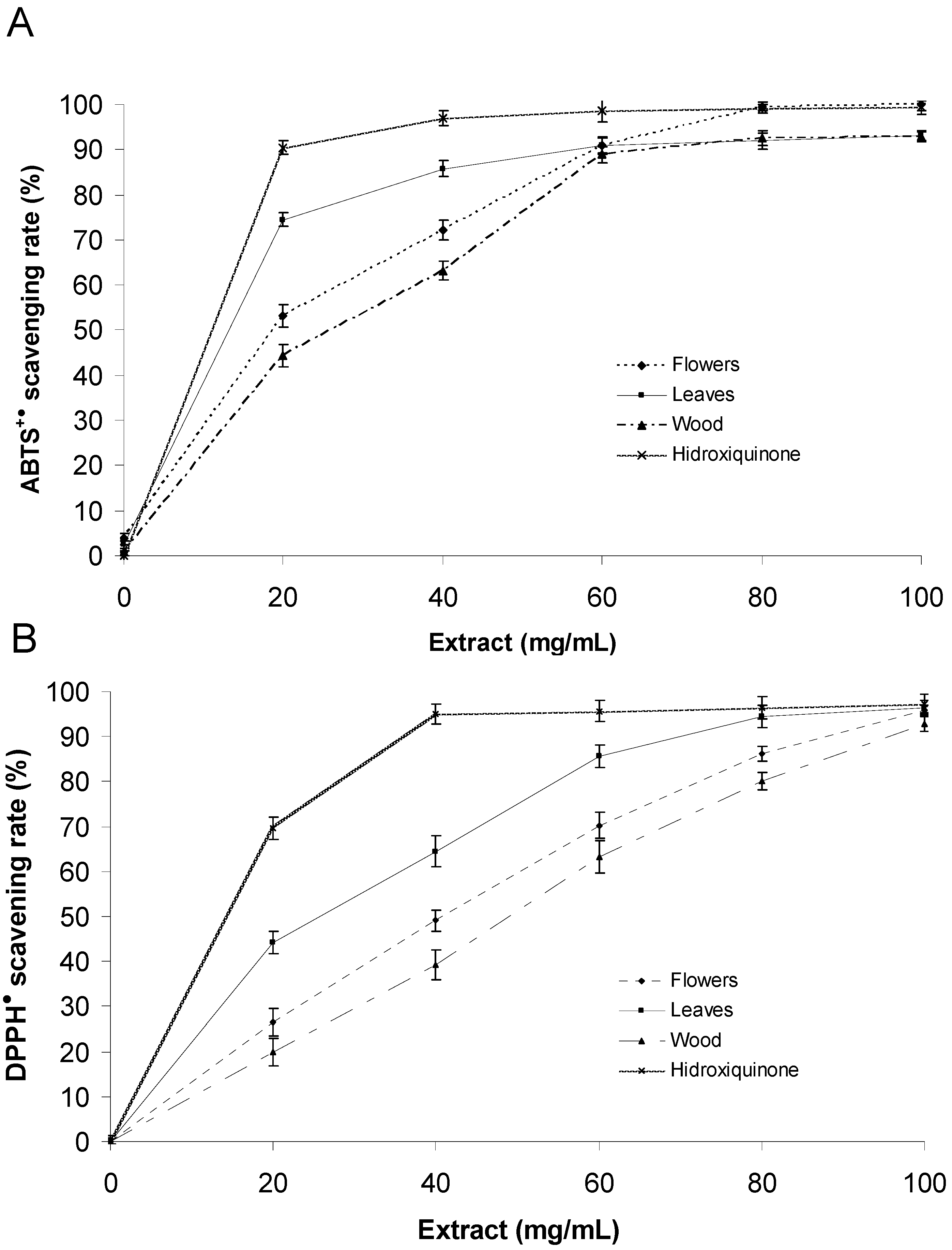
2.3. Evaluation of Antioxidant Activity

| Antioxidant activity | EC50 (µg/mL) | |||
|---|---|---|---|---|
| Flower | Leaf | Wood | Hydroxyquinone | |
| ABTS | 20.5 ± 1.2 | 15.6 ± 0.8 | 28.5 ± 0.9 | 10.2 ± 0.6 |
| DPPH | 39.7 ± 1.3 | 27.3 ± 1.1 | 47.8 ± 1.2 | 15.7 ± 0.9 |
2.4. Anti-Tubercular Activity
| Piper imperiale | MIC (µg/mL)
Mycobacterium tuberculosis H37Rv |
|---|---|
| Leaves | >500 |
| Flowers | 75 |
| Wood | >500 |
| Isoniazid | 0.1 |
2.5. Evaluation of Cytotoxicity of Natural Extract

| Extracts/Control | IC50 (µg/mL) * |
|---|---|
| Leaves | 18.6 ± 1.2 |
| Flowers | 24.5 ± 1.5 |
| Wood | 30.7 ± 1.7 |
| Curcumin | 8.2 ± 0.7 |

3. Experimental
3.1. Reagents
3.2. Plant Material and Extracts
3.3. Total Phenolic Content
3.4. HPLC Analysis
3.5. DPPH Radical Scavenging Assay

3.6. ABTS Radical Scavenging Test
3.7. Antitubercular Activity
3.8. Cytotoxicity of the Plant Extracts on MCF-7 Cell Line

3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Dyer, L.; Palmer, A. Piper: A Model Genus for Studies of Phytochemistry, Ecology, and Evolution; Springer: New York, NY, USA, 2004; pp. 156–174. [Google Scholar]
- Kramer, E.; Dorit, R.; Irish, V. Molecular evolution of the petal and stamen identity genes, APETALA3 and PISTILLATA, after petal loss in the Piperales. Genetics 1998, 149, 765–783. [Google Scholar]
- Pino-Benitez, N.; Melendez, E.; Stashenko, E. Chemical composition and antibacterial activity of the essential oil from the leaves of Piper lanceaefolium a plant traditionally used in Colombia. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 2009, 8, 301–304. [Google Scholar]
- Scott, I.; Jensen, H.; Philogène, B.; Arnason, J. A review of Piper spp. (Piperaceae) phytochemistry, insecticidal activity and mode of action. Phytochem. Rev. 2008, 7, 65–75. [Google Scholar]
- Parmar, V.; Jain, S.; Bisht, K.; Jain, R.; Taneja, P.; Jha, A.; Tyagi, O.; Prasad, A.; Wengel, J.; Olsen, C.; et al. Phytochemistry of the genus Piper. Phytochemistry 1997, 46, 597–673. [Google Scholar] [CrossRef]
- Leopoldini, M.; Russo, N.; Toscano, M. Gas and liquid phase acidity of natural antioxidants. J. Agric. Food Chem. 2006, 54, 3078–3085. [Google Scholar]
- Lavid, N.; Schwartz, A.; Yarden, O.; Tel-Or, E. The involvement of polyphenols and peroxidase activities in heavy-metal accumulation by epidermal glands of the waterlily (Nymphaeaceaceae). Planta 2001, 212, 323–331. [Google Scholar]
- Erdemoglu, S.; Gucer, S. Selective determination of aluminium bound with tannin in tea infusion. Anal. Sci. 2005, 21, 1005–1008. [Google Scholar]
- Zhu, M.; Phillipson, J.; Greengrass, P.; Bowery, N.; Cai, Y. Plant polyphenols: Biologically active compounds or nonselective binders to protein? Phytochemistry 1997, 44, 441–447. [Google Scholar] [CrossRef]
- Charlton, A.; Baxter, N.; Khan, M.; Moir, A.; Haslam, E.; Davies, A.; Williamson, M. Polyphenol/peptide binding and precipitation. J. Agric. Food Chem. 2002, 50, 1593–1601. [Google Scholar]
- Kagan, V.; Tyurina, Y. Recycling and redox cycling of phenolic antioxidants. Ann. NY Acad. Sci. 1998, 854, 425–434. [Google Scholar]
- Boudet, A. Evolution and current status of research in phenolic compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar]
- Bondia-Pons, I.; Aura, A.; Vuorela, S.; Kolehmainen, M.; Mykkanen, H.; Poutanen, K. Rye phenolics in nutrition and health. J. Cereal Sci. 2009, 49, 323–336. [Google Scholar]
- Montilla, P.; Espejo, I.; Muñoz, M.; Bujalance, I.; Muñoz-Castañeda, J.; Túnez, I. Effect of red wine on oxidative stress and hypercholesterolemia induced by feeding a high-cholesterol diet in rat. J. Physiol. Biochem. 2004, 60, 259–264. [Google Scholar]
- Brune, M.; Hallberg, L.; Skanberg, A. Determination of iron-binding phenolic groups in foods. J. Food Sci. 1991, 56, 128–131. [Google Scholar]
- Cornwell, T.; Cohick, W.; Raskin, I. Dietary phytoestrogens and health. Phytochemistry 2004, 65, 995–1016. [Google Scholar]
- Stephen, H. Green tea and the skin. J. Am. Acad. Dermatol. 2005, 52, 1049–1059. [Google Scholar]
- Duthie, G.; Duthie, S.; Kyle, J. Plant polyphenols in cancer and heart disease: Implications as nutritional antioxidants. Nutr. Res. Rev. 2000, 13, 79–106. [Google Scholar]
- Schijlen, E.; Ric de Vos, C.; van Tunen, A.; Bovy, A. Modification of flavonoid biosynthesis in crop plants. Phytochemistry 2004, 65, 2631–2648. [Google Scholar]
- Wootton-Beard, P.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin-Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Fincher, R.; Dodson, C.; Richards, J.; Tobler, M.; Searcy, J.; Mather, J.; Reid, A.; Rolig, J.; Pidcock, W. Inter- and intraspecific comparisons of antiherbivore defenses in three species of rainforest understory shrubs. J. Chem. Ecol. 2008, 34, 558–574. [Google Scholar]
- Richards, L.; Dyer, L.; Smilanich, A.; Dodson, C. Synergistic effects of amides from two Piper species on generalist and specialist herbivores. J. Chem. Ecol. 2010, 36, 1105–1113. [Google Scholar] [CrossRef]
- García-Macías, P.; Ordidge, M.; Vysini, E.; Waroonphan, S.; Battey, N.; Gordon, M.; Hadley, P.; John, P.; Lovegrove, J.; Wagstaffe, A. Changes in the flavonoid and phenolic acid contents and antioxidant activity of red leaf lettuce (Lollo Rosso) due to cultivation under plastic films varying in ultraviolet transparency. J. Agric. Food Chem. 2007, 55, 10168–10172. [Google Scholar]
- Wu, H.; Luo, J.; Raza, W.; Liu, Y.; Gu, M.; Chen, G.; Hu, X.; Wang, J.; Mao, Z.; Shen, Q. Effect of exogenously added ferulic acid on in vitro Fusarium oxysporum f. sp. niveum. Sci. Hortic. 2010, 124, 448–453. [Google Scholar] [CrossRef]
- Ghassempour, A.; Mollayi, S.; Farzaneh, M.; Sharifi-Tehrani, A.; Aboul-Enein, H. Variation of Catechin, epicatechin and their enantiomers concentrations before and after wheat cultivar—Puccinia triticina infection. Food Chem. 2011, 125, 1287–1290. [Google Scholar] [CrossRef]
- de Quirós, A.; Lage-Yusty, M.; López-Hernández, J. HPLC-analysis of polyphenolic compounds in Spanish white wines and determination of their antioxidant activity by radical scavenging assay. Food Res. Int. 2009, 42, 1018–1022. [Google Scholar]
- Hodgson, J.; Croft, K. Tea flavonoids and cardiovascular health. Mol. Aspects Med. 2010, 31, 495–502. [Google Scholar]
- Heim, K.; Tagliaferro, A.; Bobilya, D. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar]
- Orhan, D.; Ozcelik, B.; Ozgen, S.; Ergun, F. Antibacterial, antifungal, and antiviral activities of some flavonoids. Microbiol. Res. 2010, 165, 496–504. [Google Scholar] [CrossRef]
- Quilez, A.; Gilardoni, G.; Souccar, C.; de Mendonca, S.; Oliveira, L.; Martin-Calero, M.; Vidari, G. Antisecretory, anti-inflammatory and anti-Helicobacter pylori activities of several fractions isolated from Piper carpunya Ruiz & Pav. J. Ethnopharmacol. 2010, 128, 583–589. [Google Scholar] [CrossRef]
- Lizcano, L.; Bakkali, F.; Ruiz-Larrea, M.; Ruiz-Sanz, J. Antioxidant activity and polyphenol content of aqueous extracts from Colombian Amazonian plants with medicinal use. Food Chem. 2010, 119, 1566–1570. [Google Scholar]
- Mandel, S. Catechin polyphenols: Neurodegeneration and neuroprotection in neurodegenerative diseases. Free Radic. Biol. Med. 2004, 37, 304–317. [Google Scholar]
- Madhan, B.; Subramanian, V.; Rao, J.; Nair, B.; Ramasami, T. Stabilization of collagen using plant polyphenol: Role of catechin. Int. J. Biol. Macromol. 2005, 37, 47–53. [Google Scholar]
- Araújo, G.; Martel, F. Chemopreventive effect of dietary polyphenols in colorectal cancer cell lines. Nutr. Res. 2011, 31, 77–87. [Google Scholar]
- Steffen, Y.; Gruber, C.; Schewe, T.; Sies, H. Mono-O-methylated flavanols and other flavonoids as inhibitors of endothelial NADPH oxidase. Arch. Biochem. Biophy. 2008, 469, 209–219. [Google Scholar] [CrossRef]
- Arnao, M. Some methodological problems in the determination of antioxidant activity using chromogen radicals: A practical case. Trends Food Sci. Technol. 2000, 11, 419–421. [Google Scholar]
- Gramza-Michałowska, A. Evaluation of the antiradical potential of fruit and vegetable snacks. Acta Sci. Pol. Technol. Aliment. 2001, 10, 63–72. [Google Scholar]
- Péres, V.; Melecchi, M.; Abad, F.; de Assis, J.; Martinez, M.; Oliveira, E.; Caramão, E. Comparison of soxhlet, ultrasound-assisted and pressurized liquid extraction of terpenes, fatty acids and vitamin E from Piper gaudichaudianum Kunt. J. Chromatogr. A 1105, 115–118. [Google Scholar]
- Denery, J.; Dragull, K.; Tang, C.; Li, Q. Pressurized fluid extraction of carotenoids from Haematococcus pluvialis and Dunaliella salina and kavalactones from Piper methysticum. Anal. Chim. Acta 2004, 501, 175–181. [Google Scholar] [CrossRef]
- Prieto, P.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar]
- Agbor, G.; Oben, J.; Ngogang, J.; Cai, X.-X.; Vinson, J. Antioxidant capacity of some herbs/spices from Cameroon: A comparative study of two methods. J. Agric. Food Chem. 2005, 53, 6819–6824. [Google Scholar]
- Patro, B.; Tyagi, M.; Saha, J.; Chattopadhyay, S. Comparative nuclease and anti-cancer properties of the naturally occurring malabaricones. Bioorg. Med. Chem. 2010, 18, 7043–7051. [Google Scholar]
- Zhang, Q.; Zhong, Y.; Yan, L.; Sun, X.; Gong, T.; Zhang, Z. Synthesis and preliminary evaluation of curcumin analogues as cytotoxic agents. Bioorg. Med. Chem. Lett. 2011, 21, 1010. [Google Scholar]
- Kizhakkayil, J.; Thayyullathil, F.; Chathoth, S.; Hago, A.; Patel, M.; Galadari, S. Modulation of curcumin-induced Akt phosphorylation and apoptosis by PI3K inhibitor in MCF-7 cells. Biochem. Biophys. Res. Commun. 2010, 394, 476–481. [Google Scholar]
- Agbor, G.; Vinson, J.; Oben, J.; Ngogang, J. Comparative analysis of the in vitro antioxidant activity of white and black pepper. Nutr. Res. 2006, 26, 659–663. [Google Scholar] [CrossRef]
- Agbor, G.; Vinson, J.; Patel, S.; Patel, K.; Scarpati, J.; Shiner, D.; Wardrop, F.; Tompkins, T.A. Effect of selenium- and glutathione-enriched yeast supplementation on a combined atherosclerosis and diabetes hamster model. J. Agric. Food Chem. 2007, 55, 8731–8736. [Google Scholar]
- Agbor, G.; Vinson, J.; Oben, J.; Ngogang, J. Antioxidant effect of herbs and spices on copper mediated oxidation of lower and very low density lipoprotein. Chin. J. Natl. Med. 2010, 8, 114–120. [Google Scholar]
- Salleh, W.; Ahmad, F.; Yen, K.; Sirat, H. Chemical compositions, antioxidant and antimicrobial activities of essential oils of Piper caninum Blume. Int. J. Mol. Sci. 2011, 12, 7720–7731. [Google Scholar] [CrossRef]
- Rao, V.; Suresh, G.; Babu, K.; Raju, S.; Vishnu vardhan, M.; Ramakrishna, S.; Rao, J. Novel dimeric amide alkaloids from Piper chaba Hunter: Isolation, cytotoxic activity, and their biomimetic synthesis. Tetrahedron 2011, 67, 1885–1892. [Google Scholar] [CrossRef]
- Mishra, P.; Sinha, S.; Guru, S.; Bhushan, S.; Vishwakarma, R.; Ghosal, S. Two new amides with cytotoxic activity from the fruits of Piper longum. J. Asian Nat. Prod. Res. 2011, 13, 143–148. [Google Scholar]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Hanato, T.; Kagawa, H.; Yasuhara, T.; Okuda, T. Two new flavonoids and other constituents in licorice root: Their relative astringency and radical scavenging effect. Chem. Pharm. Bull. 1988, 36, 1090–1097. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar]
- Evangelopoulos, D.; Bhakta, S. Rapid methods for testing inhibitors of mycobacterial growth. Methods Mol. Biol. 2010, 642, 193–201. [Google Scholar]
- Hongo, T.; Mizuno, Y.; Haraguchi, S.; Yoshida, T. A new anticancer drug sensitivity test using the microplate culture and surviving tumor cell staining method. Gan to Kagaku Ryoho 1986, 13, 247–254. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Diaz, L.E.; Munoz, D.R.; Prieto, R.E.; Cuervo, S.A.; Gonzalez, D.L.; Guzman, J.D.; Bhakta, S. Antioxidant, Antitubercular and Cytotoxic Activities of Piper imperiale. Molecules 2012, 17, 4142-4157. https://doi.org/10.3390/molecules17044142
Diaz LE, Munoz DR, Prieto RE, Cuervo SA, Gonzalez DL, Guzman JD, Bhakta S. Antioxidant, Antitubercular and Cytotoxic Activities of Piper imperiale. Molecules. 2012; 17(4):4142-4157. https://doi.org/10.3390/molecules17044142
Chicago/Turabian StyleDiaz, Luis E., Diego R. Munoz, Rosa E. Prieto, Sergio A. Cuervo, Diego L. Gonzalez, Juan D. Guzman, and Sanjib Bhakta. 2012. "Antioxidant, Antitubercular and Cytotoxic Activities of Piper imperiale" Molecules 17, no. 4: 4142-4157. https://doi.org/10.3390/molecules17044142
APA StyleDiaz, L. E., Munoz, D. R., Prieto, R. E., Cuervo, S. A., Gonzalez, D. L., Guzman, J. D., & Bhakta, S. (2012). Antioxidant, Antitubercular and Cytotoxic Activities of Piper imperiale. Molecules, 17(4), 4142-4157. https://doi.org/10.3390/molecules17044142