A Study of Proline Metabolism in Canola (Brassica napus L.) Seedlings under Salt Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
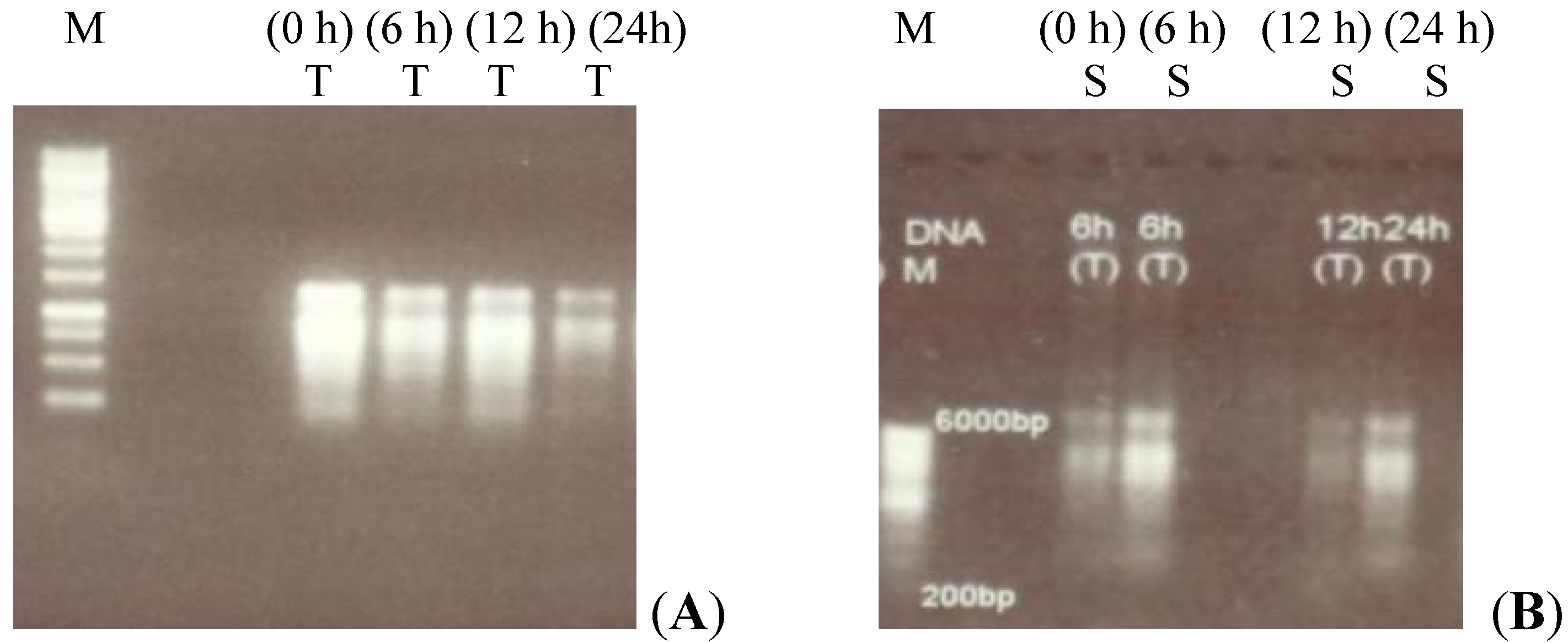
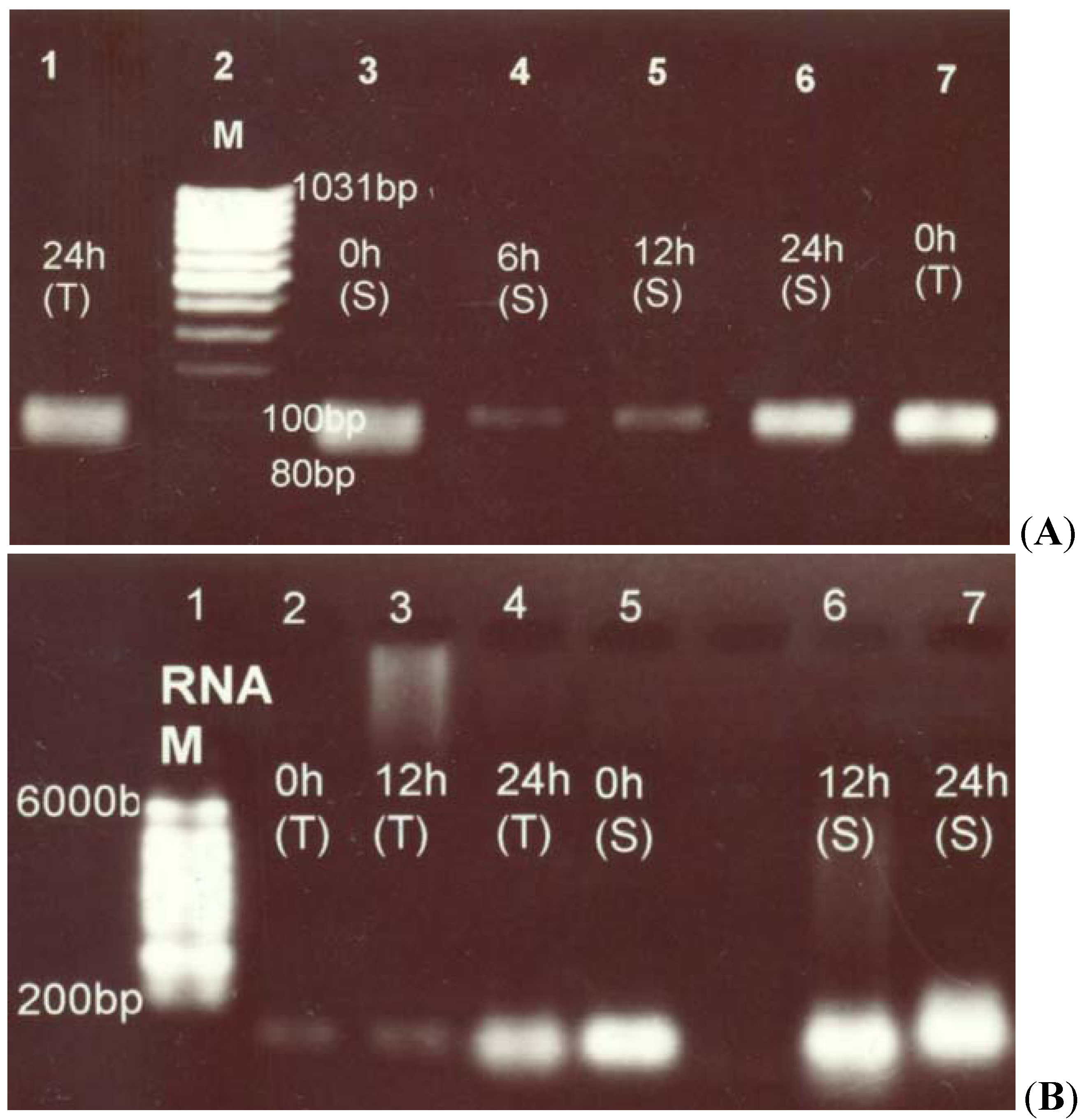
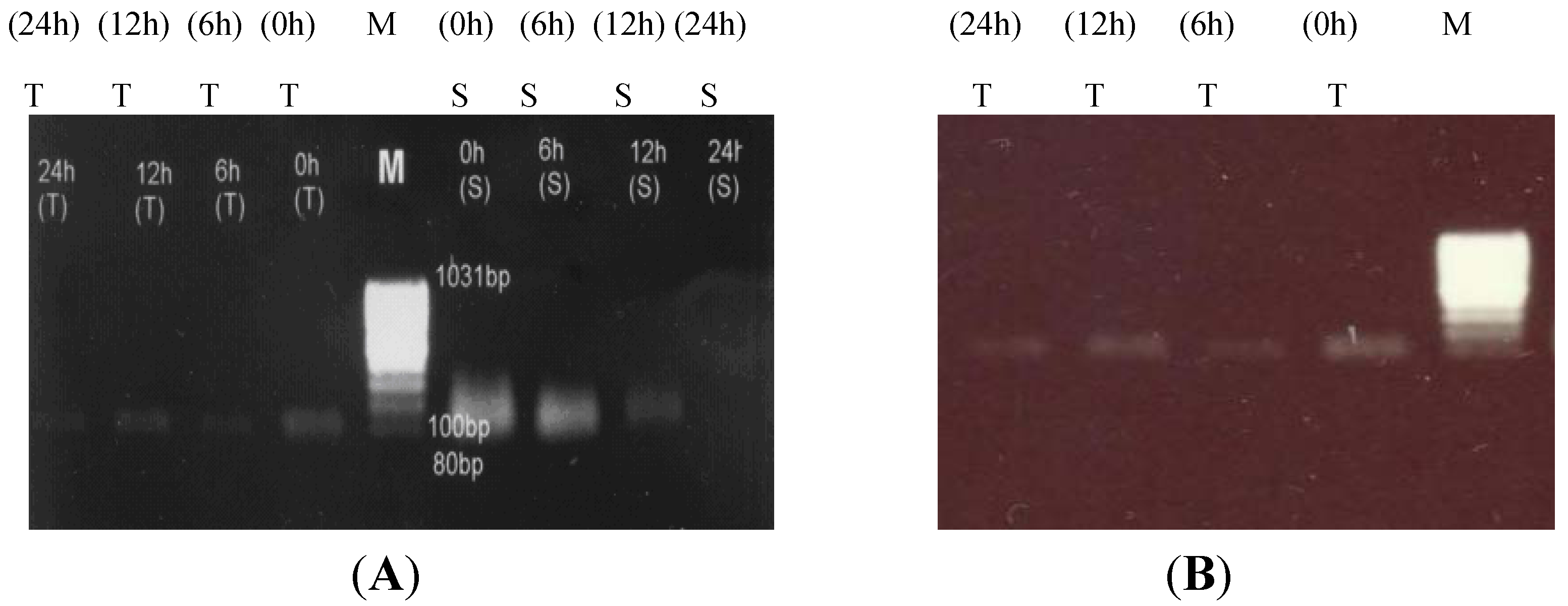
2.1. RT-PCR Analysis of Canola Gene Expression

2.2. Pyrroline-5-Carboxylate Synthetase1 (P5CS1) Up-Regulation in Canola Shoots

2.3. Proline Dehydrogenase (PDH) Down-Regulation under Salt Stress

2.4. Effect of Salt Stress on Proline Accumulation

3. Discussion
3.1. Growth of Canola Cultivars under Saline Conditions
3.2. Comparative P5CS1 Expression
3.3. Comparative PDH Expression
3.4. Comparative Total Proline Expression
4. Experimental
4.1. Plant Material and Salt Treatments
4.2. Total RNA Isolation and Reverse Transcriptase-PCR
- Δ1-Pyrroline-5-carboxylate synthetase1 (P5CS1):F 5/-GAGCTAGATCGTTCACGTGCTTT-3/R 5/-ACAACTGCTGTCCCAACCTTAAC-3/Proline dehydrogenase (PDH):F 5/-TCACAACCACTGAGCTAAAGTGAGA-3/R 5/-CGATGACGCTGTATCTTGTGATG-3/
4.3. PCR Conditions and Analysis
4.4. Total Proline Determination

5. Conclusions
Acknowledgements
References and Notes
- Akhtar, J.; Saqib, Z.A. A Profitable Way to Tackle Salinity. Dawn, the Internet Edition. Available online: http://DAWN.com (accessed on 27 September 2011).
- Koyro, H.W.; Ahmad, P.; Geissler, N. Abiotic Stress Responses in Plants: An Overview. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Nowlin, D. Winter canola. Agric. Consult. 1991, 47, 8. [Google Scholar]
- Francois, L.E. Growth, seed yield and oil content of canola grown under saline conditions. Agron. J. 1994, 86, 233–237. [Google Scholar] [CrossRef]
- Pillai, B.V.S.; Kagale, S.; Chellamma, S. Enhancing Productivity and Performance of Oil Seed Crops under Environmental Stresses. In Crop Stress and its Management: Perspectives and Strategies; Venkateswarlu, B., Ed.; Springer Science + Media: Mumbai, India, 2012; pp. 139–161. [Google Scholar]
- Al-Thabet, S.S.; Leilah, A.A.; Al-Hawass, I. Effect of NaCl and incubation temperature on seed germination of three canola (Brassica napus L.) cultivars. Pak. J. Bot. 2006, 38, 237–247. [Google Scholar]
- Qasim, M.; Ashraf, M.; Ashraf, M.Y.; Rehman, S.U.; Rha, E.S. Salt induced changes in two canola cultivars differing in salt tolerance. Biol. Plant. 2003, 46, 629–632. [Google Scholar] [CrossRef]
- Rodriguez, M.; Canales, E.; Borrás-Hidalgo, O. Molecular aspects of abiotic stress in plants. Biotechnol. Appl. 2005, 22, 1–10. [Google Scholar]
- Gaxiola, R.; Li, J.; Undurraga, S.; Dang, L.M.; Allen, G.J.; Alper, S.L.; Fink, G.R. Drought- and salt-tolerant plants result from overexpression of the AVP1 H+ pump. Proc.Natl. Acad. Sci. USA 2001, 99, 11444–11449. [Google Scholar]
- Xiong, L.; Zhu, J.K. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ. 2002, 25, 131–139. [Google Scholar] [CrossRef]
- Jimenez-Bremont, J.F.; Becerra-Flora, A.; Hernandez-Lucero, E.; Rodrıguez-Kessler, M.; Gallegos, J.A.; Ramırez-Pimentel, J.G. Proline accumulation in two bean cultivars under salt stress and the effect of polyamines and ornithine. Biol. Plant. 2006, 50, 763–766. [Google Scholar] [CrossRef]
- Tripathi, S.B.; Gurumurthi, K.; Panigrahi, A.K.; Shaw, B.P. Salinity induced changes in proline and betaine contents and synthesis in two aquatic macrophytes differing in salt tolerance. Biol. Plant. 2007, 51, 110–115. [Google Scholar] [CrossRef]
- Rajendrakumar, C.S.V.; Reddy, B.V.D.; Reddy, A.R. Proline-protein interactions: Protection of structural and functional integrity of M4 lactate dehydrogenase. Biochem. Biophys.Res. Commun. 1994, 201, 957–963. [Google Scholar] [CrossRef]
- Hong, Z.; Lakkineni, K.; Zhang, Z.; Verma, D.P.S. Removal of feedback inhibition of Δ1-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol. 2000, 122, 1129–1136. [Google Scholar] [CrossRef]
- Delauney, A.J.; Verma, D.P.S. Proline biosynthesis and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar]
- Toorchi, M.; Naderi, R.; Kanbar, A.; Shakiba, M.R. Response of spring canola cultivars to sodium chloride stress. Ann. Biol. Res. 2011, 2, 312–322. [Google Scholar]
- Nazarbeygi, E.; Yazdi, H.L.; Naseri, R.; Soleimani, R. The effects of different levels of salinity on proline and A-, B- chlorophylls in canola. American-Eurasian J. Agric. Environ. Sci. 2011, 10, 70–74. [Google Scholar]
- Xue, X.; Liu, A.; Hua, X. Proline accumulation and transcriptional regulation of proline biosynthesis and degradation in Brassica napus. BMB Rep. 2009, 42, 28–34. [Google Scholar] [CrossRef]
- Abraham, E.; Rigo, G.; Szekely, G.; Nagy, R.; Koncz, C.; Szabados, L. Light-dependent induction of proline biosynthesis by abscisic acid and salt stress is inhibited by brassinosteroid in Arabidopsis. Plant Mol. Biol. 2003, 51, 363–372. [Google Scholar] [CrossRef]
- Hellmann, H.; Funck, D.; Rentsch, D.; Frommer, W.B. Hypersensitivity of an Arabidopsis sugar signaling mutant toward exogenous proline application. Plant Physiol. 2000, 122, 357–367. [Google Scholar] [CrossRef]
- Nanjo, T.; Fujita, M.; Seki, M.; Kato, T.; Tabata, S.; Shinozaki, K. Toxicity of free proline revealed in an Arabidopsis T-DNA-tagged mutant deficient in proline dehydrogenase. Plant Cell Physiol. 2003, 44, 541–548. [Google Scholar] [CrossRef]
- Deuschle, K.; Funck, D.; Forlani, G.; Stransky, H.; Biehl, A.; Leister, D.; vander Graaff, E.; Kunze, R.; Frommer, W.B. The role of Δ1-pyrroline-5-carboxylate dehydrogenase in proline degradation. Plant Cell 2004, 16, 3413–3425. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Lv, W.T.; Lin, B.; Zhang, M.; Hua, X.J. Proline accumulation is inhibitory to Arabidopsis seedlings during heat stress. Plant Physiol. 2011, 156, 1921–1933. [Google Scholar] [CrossRef]
- Kant, S.; Kant, P.; Raveh, E.; Barak, S. Evidence that differential gene expression between the halophyte, Thellungiella halophila and Arabidopsis thaliana is responsible for higher levels of the compatible osmolyte proline and tight control of Na+ uptake in T. halophila. Plant Cell Environ. 2006, 29, 1220–1234. [Google Scholar] [CrossRef]
- Strizhov, N.; Abraham, E.; Okresch, L.; Blickling, S.; Zilberstein, A.; Schell, J.; Koncz, C.; Szabados, L. Differential expression of two P5CS genes controlling proline accumulation during salt stress requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis. Plant J. 1997, 12, 557–569. [Google Scholar]
- Yoshiba, Y.; Kiyosue, T.; Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of the level of proline as an osmolyte in plants under water stress. Plant Cell Physiol. 1997, 38, 1095–1102. [Google Scholar] [CrossRef]
- Taji, T.; Seki, M.; Satou, M.; Sakurai, T.; Kobayashi, M.; Ishiyama, K.; Narusaka, Y.; Narusaka, M.; Zhu, J.K.; Shinozaki, K. Comparative genomics in salt tolerance between Arabidopsis and Arabidopsis-related halophyte salt cress using Arabidopsis microarray. Plant Physiol. 2004, 135, 1697–1709. [Google Scholar] [CrossRef]
- Gong, Q.; Li, P.; Ma, S.; Rupassara, S.I.; Bohnert, H.J. Salinity stress adaptation competence in the extremophile Thellungiella halophila in comparison with its relative Arabidopsis thaliana. Plant J. 2005, 44, 826–839. [Google Scholar] [CrossRef]
- Hua, X.J.; van de Cotte, B.; van Montagu, M.; Verbruggen, N. Developmental regulation of pyrroline-5-carboxylate reductase gene expression in Arabidopsis. Plant Physiol. 1997, 114, 1215–1224. [Google Scholar]
- Verbruggen, N.; Villarroel, R.; Van Montagu, M. Osmoregulation of a pyrroline-5-carboxylate reductase gene in Arabidopsis thaliana. Plant Physiol. 1993, 103, 771–781. [Google Scholar]
- Kiyosue, K.; Yoshiba, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A nuclear gene encoding mitochondrial proline dehydrogenase, and enzyme involved in proline metabolism is upregulated by proline but downregulated by dehydration in Arabidopsis. Plant Cell 1996, 8, 1323–1335. [Google Scholar]
- Cechin, I.; Rossi, S.C.; Oliveira, V.C.; Fumis, T.F. Photosynthetic responses and proline content of mature and young leaves of sunflower plants under water deficit. Photosynthetica 2006, 44, 143–146. [Google Scholar] [CrossRef]
- Maggio, A.; Miyazaki, S.; Veronese, P.; Fujita, T.; Ibeas, J.I.; Damsz, B.; Narasimhan, M.L.; Hasegawa, P.M.; Joly, R.J.; Bressan, R.A. Does proline accumulation play an active role in stress-induced growth reduction? Plant J. 2002, 31, 699–712. [Google Scholar] [CrossRef]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Mohammad, F.; Khan, M.N. Morphological and physio-biochemical characterization of Brassica juncea L. Czern. & Coss. genotypes under salt stress. J. Plant Interact. 2009, 4, 67–80. [Google Scholar]
- Heidari, M. Nucleic acid metabolism, proline concentration and antioxidants enzyme activity in canola (Brassica nupus L.) under salinity stress. Agric. Sci. China 2010, 9, 504–511. [Google Scholar]
- Omidi, H. Changes of proline content and activity of antioxidative enzymes in two canola genotype under drought stress. Am. J. Plant Physiol. 2010, 5, 338–349. [Google Scholar] [CrossRef]
- He, X.J.; Mu, R.L.; Cao, W.H.; Zhang, Z.G.; Zhang, J.S.; Chen, S.Y. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005, 44, 903–916. [Google Scholar] [CrossRef]
- Ma, S.S.; Gong, Q.Q.; Bohnert, H.J. Dissecting salt stress pathways. J. Exp. Bot. 2006, 57, 1097–1107. [Google Scholar] [CrossRef]
- Weltmeier, F.; Ehlert, A.; Mayer, C.S.; Dietrich, K.; Wang, X.; Schutze, K.; Alonso, R.; Harter, K.; Vicente-Carbajosa, J.; Droge-Laser, W. Combinatorial control of Arabidopsis proline dehydrogenase transcription by specific heterodimerisation of bZIP transcription factors. EMBO J. 2006, 25, 3133–3143. [Google Scholar] [CrossRef]
- Athar, H.-R.; Ashraf, M.; Wahid, A.; Jamil, A. Inducing salt tolerance in canola (Brassica napus L.) by exogenous application of glycinebetaine and proline: Response at the initial growth stages. Pak. J. Bot. 2009, 41, 1311–1319. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Univ. California Agric. Exp. Stn. Circ. 1950, 347, 1–32. [Google Scholar]
- Sambrook, J.; Russel, D.W. Molecular Cloning. A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Sample Availability: Contact the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Saadia, M.; Jamil, A.; Akram, N.A.; Ashraf, M. A Study of Proline Metabolism in Canola (Brassica napus L.) Seedlings under Salt Stress. Molecules 2012, 17, 5803-5815. https://doi.org/10.3390/molecules17055803
Saadia M, Jamil A, Akram NA, Ashraf M. A Study of Proline Metabolism in Canola (Brassica napus L.) Seedlings under Salt Stress. Molecules. 2012; 17(5):5803-5815. https://doi.org/10.3390/molecules17055803
Chicago/Turabian StyleSaadia, Mubshara, Amer Jamil, Nudrat Aisha Akram, and Muhammad Ashraf. 2012. "A Study of Proline Metabolism in Canola (Brassica napus L.) Seedlings under Salt Stress" Molecules 17, no. 5: 5803-5815. https://doi.org/10.3390/molecules17055803
APA StyleSaadia, M., Jamil, A., Akram, N. A., & Ashraf, M. (2012). A Study of Proline Metabolism in Canola (Brassica napus L.) Seedlings under Salt Stress. Molecules, 17(5), 5803-5815. https://doi.org/10.3390/molecules17055803