Ursolic Acid Inhibits Cigarette Smoke Extract-Induced Human Bronchial Epithelial Cell Injury and Prevents Development of Lung Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. UA Reduces CSE-Induced Cytotoxicity in NHBE Cells

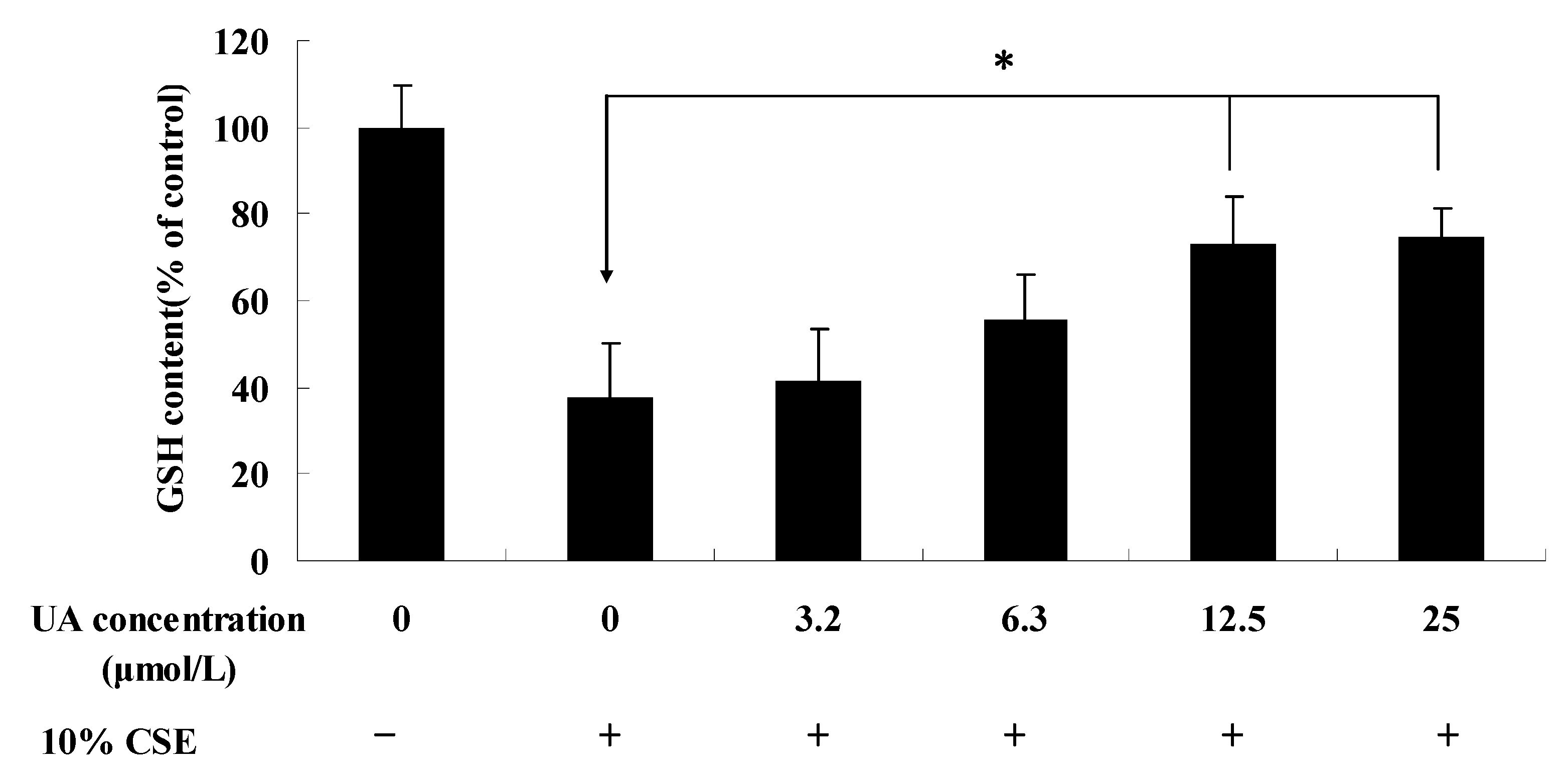
2.2. UA Palliates CSE-Induced GSH Content Reduction

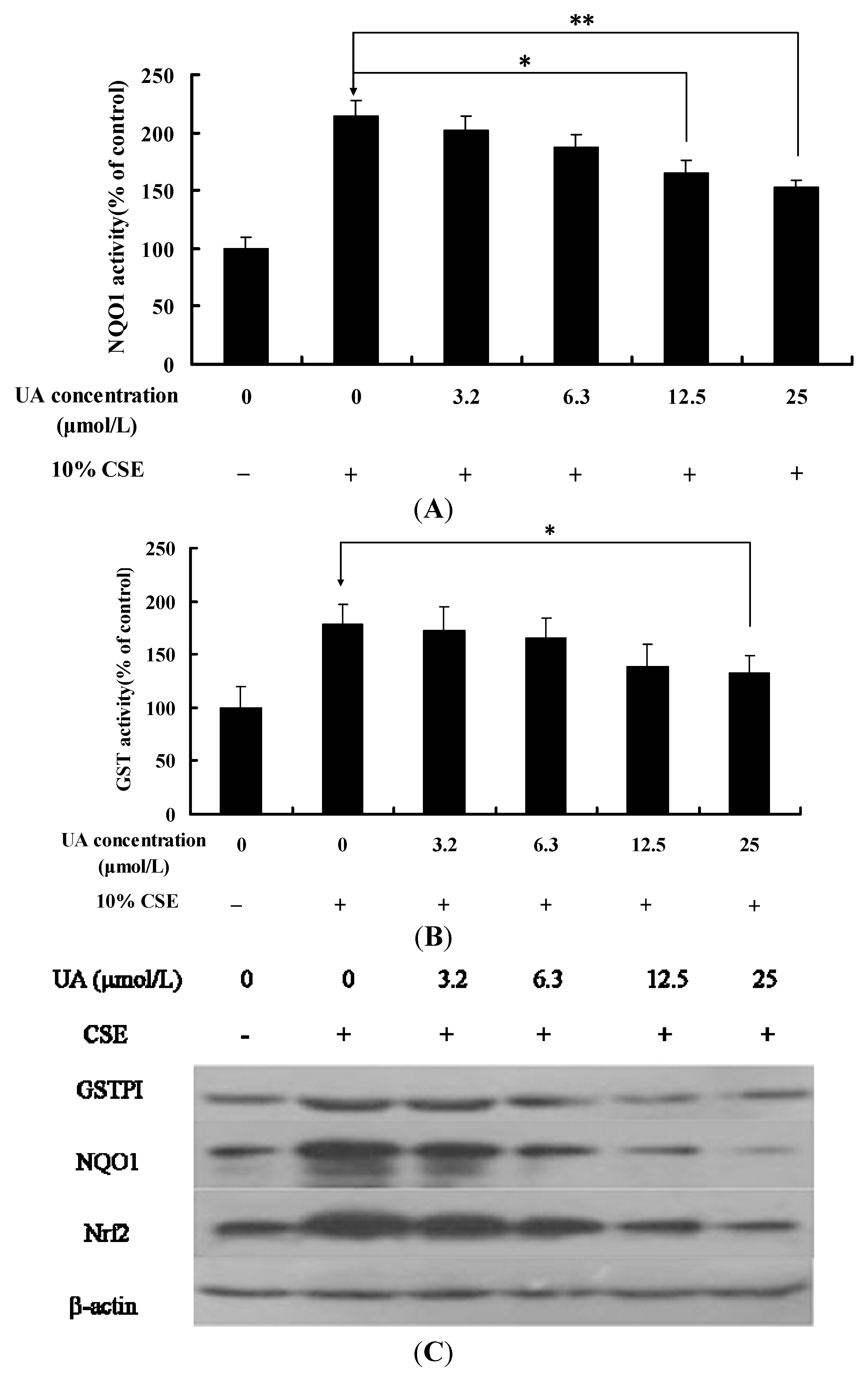
2.3. UA Alleviates CSE-Induced GST and NQO1 Activity and Protein Expression

2.4. UA Palliates CSE-Induced DNA Damage

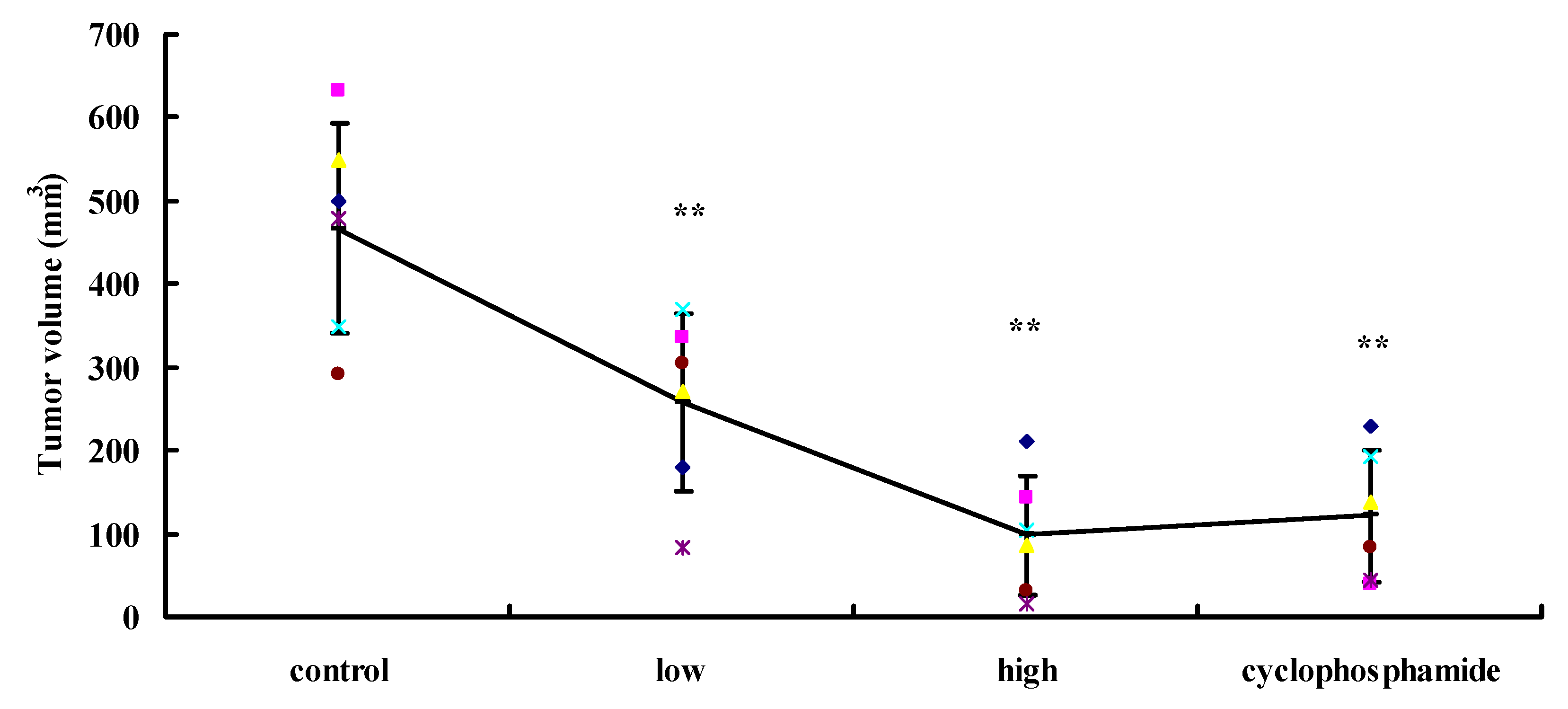
2.5. UA Inhibits Tumorigenesis in Vivo

3. Experimental
3.1. Chemicals
3.2. Cell Culture
3.3. Preparation of CSE
3.4. Cytotoxicity and Viability Assays

3.5. Estimation of DNA Damage by Comet Assay
3.6. Lactate Dehydrogenase Content
3.7. Whole Cell Lysate Preparation
3.8. Measurement of Intracellular GSH Levels
3.9. Analysis of Detoxification Enzyme Activity
3.10. Western Blot Analysis
3.11. Effect of UA on Tumor Growth in Nude Mice
3.12. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Carnevali, S.; Petruzzelli, S.; Longoni, B.; Vanacore, R.; Barale, R.; Cipollini, M.; Scatena, F.; Paggiaro, P.; Celi, A.; Giuntini, C. Cigarette smoke extract induces oxidative stress and apoptosis in human lung fibroblasts. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2003, 284, L955–L963. [Google Scholar]
- Rahman, I.; Biswas, S.K.; Kode, A. Oxidant and antioxidant balance in the airways and airway diseases. Eur. J. Pharm. 2006, 533, 222–239. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Lee, J.S.; Surh, Y.J. Nrf2 as a novel molecular target for chemoprevention. Cancer Lett. 2005, 224, 171–184. [Google Scholar] [CrossRef]
- Tan, X.L.; Spivack, S.D. Dietary chemoprevention strategies for induction of phase II xenobiotic-metabolizing enzymes in lung carcinogenesis: A review. Lung Cancer 2009, 65, 129–137. [Google Scholar] [CrossRef]
- Wilkinson, J.T.; Clapper, M.L. Detoxication enzymes and chemoprevention. Proc. Soc. Exp. Biol. Med. 1997, 216, 192–200. [Google Scholar]
- Kwak, M.K.; Wakabayashi, N.; Kensler, T.W. Chemoprevention through the Keap12Nrf2 signaling pathway by phase 2 enzyme inducers. Mutat. Res. 2004, 555, 133–148. [Google Scholar] [CrossRef]
- Slocum, •S.T.; Kensler, T.W. Nrf2: Control of sensitivity to carcinogens. Arch. Toxicol. 2011, 85, 273–284. [Google Scholar] [CrossRef]
- Miao, W.; Hu, L.; Scrivens, P.J.; Batist, G. Transcriptional regulation of NRF2 expression by the AHR-XRE signaling pathway: Direct cross-talk between phase I and II drug-metabolizing enzymes. J. Biol. Chem. 2005, 280, 20340–20348. [Google Scholar]
- Feng, L.; Jia, X.-B.; Shi, F.; Chen, Y. Identification of two polysaccharides from Prunella vulgaris L. and evaluation on their anti-lung adenocarcinoma activity. Molecules 2010, 15, 5093–5103. [Google Scholar]
- Feng, L.; Jia, X.-B.; Zhu, M.; Chen, Y.; Shi, F. Chemoprevention by Prunella vulgaris L. extract of non-small cell lung cancer via promoting apoptosis and regulating the cell cycle. Asian Pac. J. Cancer Prev. 2010, 11, 1355–1358. [Google Scholar]
- Feng, L.; Jia, X.-B.; Jiang, J.; Zhu, M.-M.; Chen, Y.; Tan, X.-B.; Shi, F. Combination of active components enhances the efficacy of prunella in prevention and treatment of lung cancer. Molecules 2010, 15, 7893–7906. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Chang, J.Y.; Kuo, C.C.; Chang, C.Y.; Kuo, Y.H. Cytotoxic triterpenes from the aerial roots of Ficusmicrocarpa. Phytochemistry 2005, 66, 495–501. [Google Scholar]
- Lee, J.H.; Lee, D.U.; Jeong, C.S. Gardenia jasminoides Ellisethanol extract and its constituents reduce the risks of gastritis and reverse gastric lesions in rats. Food Chem. Toxicol. 2009, 47, 1127–1131. [Google Scholar] [CrossRef]
- Yamai, H.; Sawada, N.; Yoshida, T.; Seike, J.; Takizawa, H.; Kenzaki, K.; Miyoshi, T.; Kondo, K.; Bando, Y.; Ohnishi, Y.; et al. Triterpenes augment the inhibitory effects of anticancer drugs on growth of human esophageal carcinoma cells in vitro and suppress experimental metastasis in vivo. Int. J. Cancer 2009, 125, 952–960. [Google Scholar] [CrossRef]
- Yan, S.L.; Huang, C.Y.; Wu, S.T.; Yin, M.C. Oleanolic acid and ursolic acid induce apoptosis in four human liver cancer cell lines. Toxicol. In Vitro 2010, 24, 842–848. [Google Scholar] [CrossRef]
- Yang, L.; Liu, X.; Lu, Z.; Chan, J.Y.-W.; Zhou, L.; Fung, K.P.; Wu, P.; Wu, S. Ursolic acid induces doxorubicin-resistant HepG2 cell death via the release of apoptosis-inducing factor. Cancer Lett. 2010, 298, 128–138. [Google Scholar]
- Liu, X.; Conner, H.; Kobayashi, T.; Kim, H.; Wen, F.; Abe, S.; Fang, Q.; Wang, X.; Hashimoto, M.; Bitterman, P.; et al. Cigarette smoke extract induces DNA damage but not apoptosis in human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 2005, 33, 121–129. [Google Scholar] [CrossRef]
- Togo, S.; Sugiura, H.; Nelson, A.; Kobayashi, T.; Wang, X.; Kamio, K.; Kawasaki, S.; Bitterman, P.; Rennard, S.I.; Liu, X. Hepatic growth factor (HGF) inhibits cigarette smoke extract induced apoptosis in human bronchial epithelial cells. Exp. Cell Res. 2010, 316, 3501–3516. [Google Scholar] [CrossRef]
- Su, Y.; Han, W.; Giraldo, C.; de Li, Y.; Block, E.R. Effect of cigarette smoke extract on nitric oxide synthase in pulmonaryartery endothelial cells. Am. J. Respir. Cell. Mol. Biol. 1998, 19, 819–825. [Google Scholar]
- Chen, Z.-L.; Tao, J.; Yang, J.; Yuan, Z.-L.; Liu, X.H.; Jin, M.; Shen, Z.Q.; Wang, L.; Li, H.F.; Qiu, Z.G.; et al. Vitamin E modulates cigarette smoke extract-induced cell apoptosis in mouse embryonic cells. Int. J. Biol. Sci. 2011, 7, 927–936. [Google Scholar]
- Kim, H.; Liu, X.; Kobayashi, T.; Conner, H.; Kohyama, T.; Wen, F.-Q.; Fang, Q.; Abe, S.; Bitterman, P.; Rennard, S.I. Reversible cigarette smoke extract-induced DNA damage in human lung fibroblasts. Am. J. Respir. Cell. Mol. Biol. 2004, 31, 483–490. [Google Scholar] [CrossRef]
- Aldhous, M.C.; Soo, K.; Stark, L.A.; Ulanicja, A.A.; Easterbrook, J.E.; Dunlop, M.G.; Satsanqi, J. Cigarette smoke extract (CSE) Delays NOD2 expression and affects NOD2/RIPK2 interactions in intestinal epithelial cells. PLoS One 2011, 6, e24715. [Google Scholar]
- Cabello, C.M.; Bair, W.B., 3rd; Bause, A.S.; Wondrak, G.T. Antimelanoma activity of the redox dye DCPIP (2,6-dichlorophenolindophenol) is antagonized by NQO1. Biochem. Pharmacol. 2009, 78, 344–354. [Google Scholar]
- Sample Availability: Samples of the compound ursolic acid are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, W.; Tan, X.; Shu, L.; Sun, H.; Song, J.; Jin, P.; Yu, S.; Sun, M.; Jia, X. Ursolic Acid Inhibits Cigarette Smoke Extract-Induced Human Bronchial Epithelial Cell Injury and Prevents Development of Lung Cancer. Molecules 2012, 17, 9104-9115. https://doi.org/10.3390/molecules17089104
Liu W, Tan X, Shu L, Sun H, Song J, Jin P, Yu S, Sun M, Jia X. Ursolic Acid Inhibits Cigarette Smoke Extract-Induced Human Bronchial Epithelial Cell Injury and Prevents Development of Lung Cancer. Molecules. 2012; 17(8):9104-9115. https://doi.org/10.3390/molecules17089104
Chicago/Turabian StyleLiu, Wenbo, Xiaobin Tan, Luan Shu, Hanyan Sun, Jie Song, Ping Jin, Siming Yu, Min Sun, and Xiaobin Jia. 2012. "Ursolic Acid Inhibits Cigarette Smoke Extract-Induced Human Bronchial Epithelial Cell Injury and Prevents Development of Lung Cancer" Molecules 17, no. 8: 9104-9115. https://doi.org/10.3390/molecules17089104
APA StyleLiu, W., Tan, X., Shu, L., Sun, H., Song, J., Jin, P., Yu, S., Sun, M., & Jia, X. (2012). Ursolic Acid Inhibits Cigarette Smoke Extract-Induced Human Bronchial Epithelial Cell Injury and Prevents Development of Lung Cancer. Molecules, 17(8), 9104-9115. https://doi.org/10.3390/molecules17089104