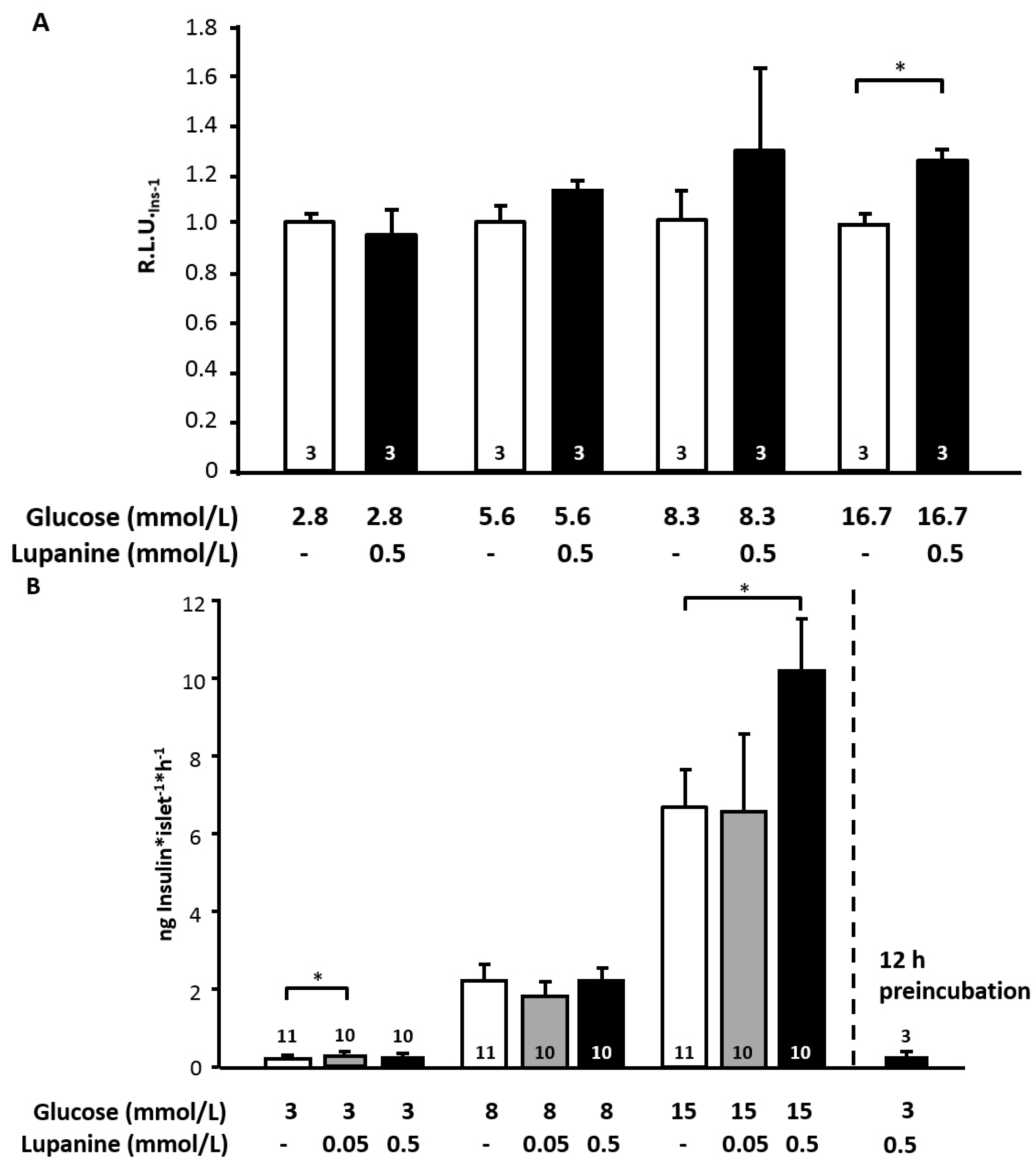
2.1. Lupanine Lowers Plasma Glucose Concentration and Improves Glycemic Control in an in Vivo Model for Diabetes
To characterize the acute effect of lupanine, 20 mg/kg body weight (BW) of the alkaloid were orally administered in non-diabetic and diabetic rats. After 30 min, an oral glucose tolerance test (oGTT, glucose: 2 g/kg BW) was performed. This dosage of lupanine did not lower blood glucose, which was 4.4 ± 0.2 mg/dL before and 4.9 ± 0.2 mg/dL 30 min after administration of lupanine (
n = 3). In comparison to control animals, the rise in plasma glucose concentration during the oGTT expressed as area under the curve (AUC) tended to be reduced in lupanine-treated rats but the effect was not significant (
Figure 1A,B). To test whether lupanine effectively improves glucose tolerance in diabetic animals, the acute, low-dose streptozotocin (STZ) protocol was used. This series of experiments revealed a beneficial effect of lupanine in diabetic animals (
Figure 1C). Glucose tolerance was improved 60 and 90 min after administration of the glucose bolus. In agreement with a better glycemic control, the AUC was significantly smaller in lupanine-treated rats compared to untreated diabetic animals (
Figure 1D). Determination of insulin sensitivity revealed that lupanine did not change the response of STZ-diabetic rats to exogenous insulin (0.5 I.U./kg BW, intraperitoneal injection) in comparison to untreated control animals (
Figure 1E).
Figure 1.
Influence of lupanine on glycemic control. Changes in blood glucose concentration in response to an oral application of 2 g glucose/kg BW (A–D) or in response to intraperitoneal injection of 0.5 I.U. insulin/kg BW (E); (A) Diagram showing the changes in blood glucose concentration (left panel: absolute values, right panel: changes from baseline). The dashed line illustrates the data during the oGTT of control rats, the continuous line represents the results for lupanine-treated rats (20 mg/kg BW, oral administration 30 min before oGTT); (B) Calculation of the area under the curve (AUC) for a period of 90 min after application of the glucose bolus for control (white bar) and lupanine-treated (black bar) animals; (C,D) Same maneuver as described for (A,B) but performed with STZ-diabetic rats. STZ-treatment drastically impaired glucose tolerance (compare white bars in (B,D)). Change of blood glucose concentration (C) and AUC (D) were significantly improved by the administration of lupanine; (E) Insulin-induced reduction of blood glucose concentration in STZ-diabetic rats without (dashed line) or with lupanine (continuous line) treatment. The number of animals is given in the bars of (B,D,E). * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001.
Figure 1.
Influence of lupanine on glycemic control. Changes in blood glucose concentration in response to an oral application of 2 g glucose/kg BW (A–D) or in response to intraperitoneal injection of 0.5 I.U. insulin/kg BW (E); (A) Diagram showing the changes in blood glucose concentration (left panel: absolute values, right panel: changes from baseline). The dashed line illustrates the data during the oGTT of control rats, the continuous line represents the results for lupanine-treated rats (20 mg/kg BW, oral administration 30 min before oGTT); (B) Calculation of the area under the curve (AUC) for a period of 90 min after application of the glucose bolus for control (white bar) and lupanine-treated (black bar) animals; (C,D) Same maneuver as described for (A,B) but performed with STZ-diabetic rats. STZ-treatment drastically impaired glucose tolerance (compare white bars in (B,D)). Change of blood glucose concentration (C) and AUC (D) were significantly improved by the administration of lupanine; (E) Insulin-induced reduction of blood glucose concentration in STZ-diabetic rats without (dashed line) or with lupanine (continuous line) treatment. The number of animals is given in the bars of (B,D,E). * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001.
![Molecules 20 19085 g001a]()
![Molecules 20 19085 g001b]()
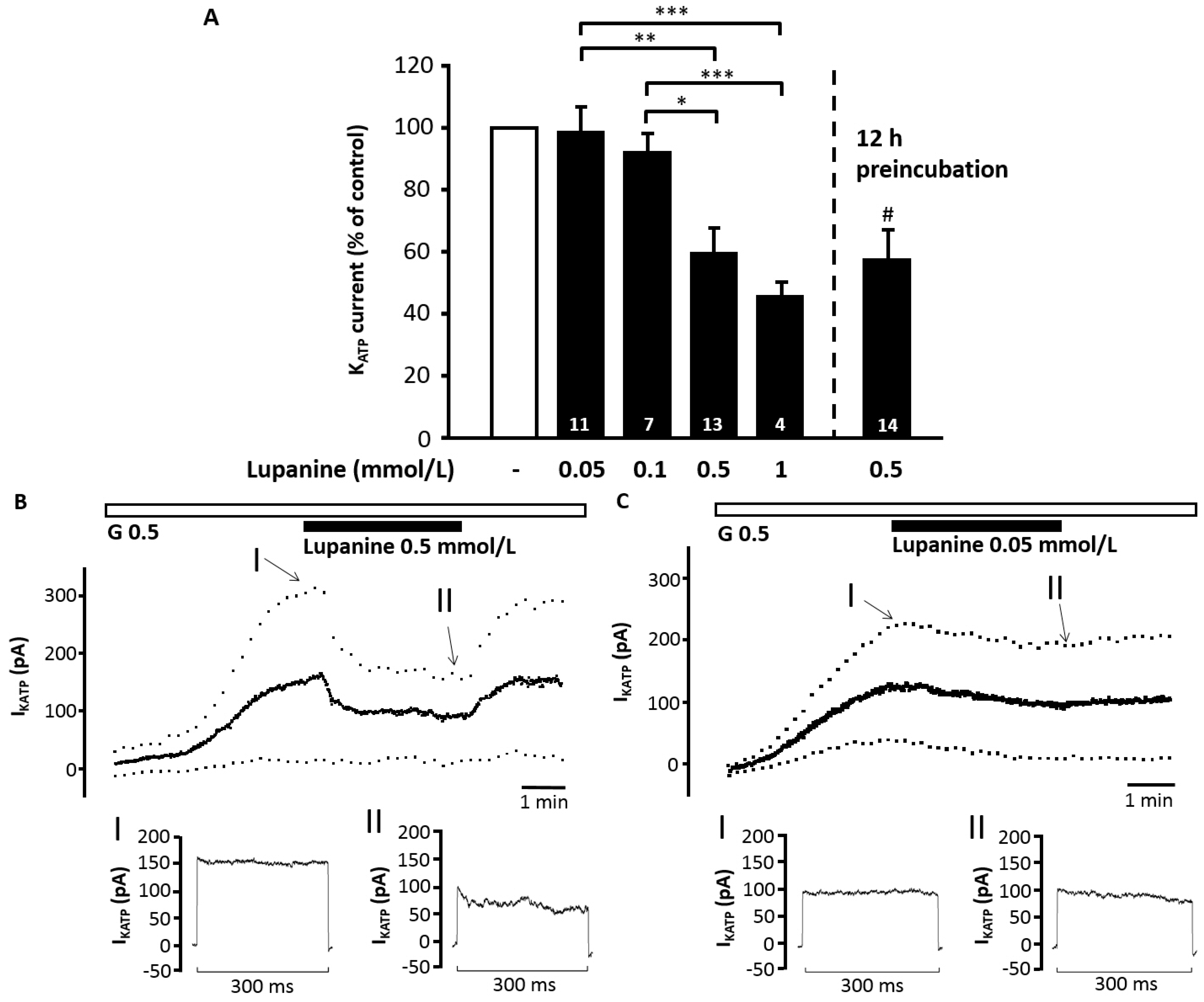
2.4. Lupanine Reduces the Current through KATP Channels and Modifies Electrical Activity of Beta Cells
To check whether the stimulatory action of the alkaloid is mediated by an effect on the stimulus-secretion cascade measurements of ion currents and plasma membrane potential (V
m) were performed. Lupanine had no effect on K
ATP current at a concentration range of 0.05 to 0.1 mmol/L but dose-dependently and reversibly inhibited the current at concentrations of 0.5 mmol/L (from 123 ± 14 pA to 73 ± 12 pA,
n = 13,
p ≤ 0.001) and 1 mmol/L (from 176 ± 42 pA to 84 ± 27 pA,
n = 4,
p ≤ 0.01) (
Figure 3A–C). In cells incubated with 0.5 mmol/L lupanine for 12 h the current was reduced to a similar extent as obtained by acute treatment (control cells: 119 ± 34 pA,
n = 12
vs. lupanine-pretreated cells: 70 ± 14 pA,
n = 14,
p ≤ 0.05). As these experiments were performed in the standard whole-cell configuration (
i.e., without cell metabolism), the data indicate a direct interaction of lupanine with K
ATP channels. The K
ATP channel opener diazoxide (0.1 mmol/L) only transiently antagonized the inhibitory effect of 1 mmol/L lupanine (K
ATP current in the presence of lupanine: 84 ± 27 pA, maximal increase after addition of diazoxide: 110 ± 29 pA,
n = 4,
p ≤ 0.01). Control experiments with tolbutamide confirmed that the current determined by this protocol was exclusively K
ATP current (control: 177 ± 23 pA, after addition of 100 µM tolbutamide: 4 ± 0.4 pA,
n = 6,
p ≤ 0.01).
Figure 3.
Lupanine reduces the KATP current. KATP current of isolated islet cells was determined in the standard whole-cell configuration, i.e., without cell metabolism. Bath solution contained 0.5 mmol/L glucose (G 0.5). (A) Summary of the dose–response relation for the acute application of 0.05–1 mmol/L lupanine (left bars) or the effect of preincubation with 0.5 mmol/L lupanine for 12 h (right bar). Values are normalized to the current recorded under control conditions directly before changing bath solution and in the preincubation experiments to the current of untreated control cells. The two experiments illustrate representative recordings with 0.5 (B) and 0.05 (C) mmol/L lupanine. The time points marked with “I, II” are shown below the traces in higher temporal resolution. The number of cells tested for each condition is given in the bars of the diagram. * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, # p ≤ 0.05 vs. untreated control cells.
Figure 3.
Lupanine reduces the KATP current. KATP current of isolated islet cells was determined in the standard whole-cell configuration, i.e., without cell metabolism. Bath solution contained 0.5 mmol/L glucose (G 0.5). (A) Summary of the dose–response relation for the acute application of 0.05–1 mmol/L lupanine (left bars) or the effect of preincubation with 0.5 mmol/L lupanine for 12 h (right bar). Values are normalized to the current recorded under control conditions directly before changing bath solution and in the preincubation experiments to the current of untreated control cells. The two experiments illustrate representative recordings with 0.5 (B) and 0.05 (C) mmol/L lupanine. The time points marked with “I, II” are shown below the traces in higher temporal resolution. The number of cells tested for each condition is given in the bars of the diagram. * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, # p ≤ 0.05 vs. untreated control cells.
Based on the results obtained for insulin release the inhibitory effect of 0.5 mmol/L lupanine on K
ATP current should not be large enough to induce electrical activity
per se. However, it might already affect V
m. To test for this, V
m was measured in unstimulated beta cells with intact metabolism. Lupanine did not affect V
m in bath solution with low glucose (0.5 mmol/L) or at glucose concentrations close to the threshold for activation (5–6 mmol/L glucose) (
Figure 4A,B). As 0.05 mmol/L lupanine slightly elevated insulin secretion in bath solution supplemented with 3 mmol/L glucose, V
m was also determined under these conditions. These experiments showed that the effect on basal insulin release was not mediated by any changes in V
m which was −69.8 ± 1.7 mV in the presence of 3 mmol/L glucose and −70.0 ± 1.8 mV after addition of 0.05 mmol/L lupanine (
n = 12).
Figure 4.
Lupanine has no effect on Vm of unstimulated beta cells. Cells were stimulated with 15 mmol/L glucose (G 15) at the beginning and the end of each experiments to verify metabolic integrity. Lupanine (0.5 mmol/L) was added in the presence of 0.5 mmol/L glucose (A) or at a glucose concentration of 5–6 mmol/L (B), which is close to the threshold for induction of electrical activity. Under both conditions lupanine did not depolarize the cells. The left part of (A,B) shows representative recordings with 0.5 mmol/L (G 0.5) and 6 mmol/L (G 6) glucose, respectively. The right part summarizes the data. White bars: Vm in the presence of 0.5 or 5–6 mmol/L glucose, black bars: + lupanine, grey bars: depolarization of Vm induced by 15 mmol/L glucose at the end of the experiments. The number of cells tested for each condition is given in the bars of the diagram. *** p ≤ 0.001.
Figure 4.
Lupanine has no effect on Vm of unstimulated beta cells. Cells were stimulated with 15 mmol/L glucose (G 15) at the beginning and the end of each experiments to verify metabolic integrity. Lupanine (0.5 mmol/L) was added in the presence of 0.5 mmol/L glucose (A) or at a glucose concentration of 5–6 mmol/L (B), which is close to the threshold for induction of electrical activity. Under both conditions lupanine did not depolarize the cells. The left part of (A,B) shows representative recordings with 0.5 mmol/L (G 0.5) and 6 mmol/L (G 6) glucose, respectively. The right part summarizes the data. White bars: Vm in the presence of 0.5 or 5–6 mmol/L glucose, black bars: + lupanine, grey bars: depolarization of Vm induced by 15 mmol/L glucose at the end of the experiments. The number of cells tested for each condition is given in the bars of the diagram. *** p ≤ 0.001.
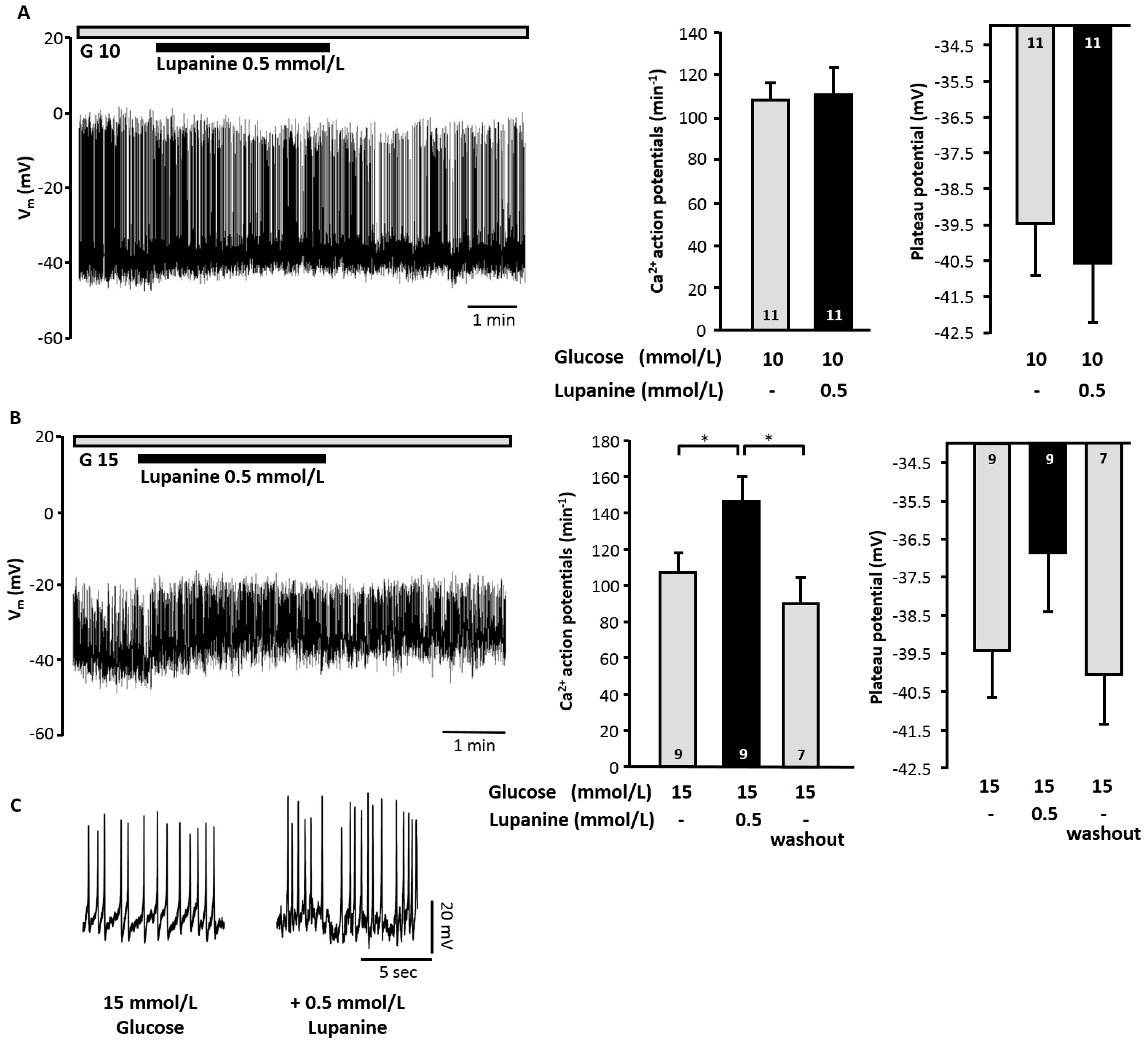
To investigate whether the potentiating effect of lupanine on insulin secretion in glucose-stimulated islets was caused by alterations in V
m we tested the influence of lupanine on beta cells that were already electrically active. The frequency of Ca
2+ action potentials was unaffected by 0.5 mmol/L lupanine in the presence of 10 mmol/L glucose (
Figure 5A) but—in agreement with the results obtained for insulin release—increased by ~40% in the presence of 15 mmol/L glucose (
Figure 5B,C). Similar to the inhibitory influence on K
ATP currents the effects of lupanine on action potential frequency was reversible (
Figure 5B). Correspondingly, the plateau potential (potential at which Ca
2+ action potentials start) was slightly more depolarized after addition of lupanine. This increase occurred in all experiments but was not statistically different.
Figure 5.
Lupanine increases electrical activity of glucose-stimulated beta cells. Lupanine has no effect on Vm and on the frequency of Ca2+ action potentials in the presence of 10 mmol/L glucose (G 10) (A) but increases the frequency of action potentials in the presence of 15 mmol/L glucose (G 15) (B). The left part of (A,B) shows representative recordings, in the middle of (A,B) analysis of action potential frequency is summarized in the diagrams. The diagrams on the right illustrate the influence of lupanine on the plateau potential, i.e., the potential at which Ca2+ action potentials start; and (C) illustrates changes in action potentials of the series of experiments presented in (B) in a higher temporal resolution. The number of cells tested for each condition is given in the bars of the diagrams. * p ≤ 0.05.
Figure 5.
Lupanine increases electrical activity of glucose-stimulated beta cells. Lupanine has no effect on Vm and on the frequency of Ca2+ action potentials in the presence of 10 mmol/L glucose (G 10) (A) but increases the frequency of action potentials in the presence of 15 mmol/L glucose (G 15) (B). The left part of (A,B) shows representative recordings, in the middle of (A,B) analysis of action potential frequency is summarized in the diagrams. The diagrams on the right illustrate the influence of lupanine on the plateau potential, i.e., the potential at which Ca2+ action potentials start; and (C) illustrates changes in action potentials of the series of experiments presented in (B) in a higher temporal resolution. The number of cells tested for each condition is given in the bars of the diagrams. * p ≤ 0.05.
2.5. Lupanine Potentiates the Influence of l-Arginine on Insulin Release
As our data indicate that lupanine acts by augmentation of electrical activity we tested whether potentiation of insulin release is shifted to lower glucose concentrations when cells are already partly depolarized by
l-arginine. Lupanine (0.5 mmol/L) in combination with
l-arginine (10 mmol/L) had no effect in the presence of 3 mmol/L glucose (0.47 ± 0.12
vs. 0.39 ± 0.17 ng/(islet·h) insulin,
n = 5) but elevated insulin release in the presence of 8 and 15 mmol/L glucose (
Figure 6).
Figure 6.
Effect of lupanine in combination with l-arginine on insulin secretion. Murine islets were incubated with 0.5 mmol/L lupanine for 60 min in the presence of 10 mmol/L l-arginine and 8 mmol/L glucose (A) or 15 mmol/L glucose (B). The number of independent experiments is given in the bars. * p ≤ 0.05, ** p ≤ 0.01.
Figure 6.
Effect of lupanine in combination with l-arginine on insulin secretion. Murine islets were incubated with 0.5 mmol/L lupanine for 60 min in the presence of 10 mmol/L l-arginine and 8 mmol/L glucose (A) or 15 mmol/L glucose (B). The number of independent experiments is given in the bars. * p ≤ 0.05, ** p ≤ 0.01.
Our
in vivo investigation shows that lupanine improves glucose homeostasis in STZ-diabetic animals but not in normoglycemic controls. These data are in agreement with human studies which also demonstrate that the effect of
Lupinus raw material or extracts depends on the glycemic status of the patients [
3]. The lack of effect on insulin sensitivity suggests that lupanine interacts with the endocrine pancreas. Analysis of beta cell stimulus-secretion coupling revealed that lupanine directly inhibits K
ATP channels. The transient, antagonizing effect of diazoxide most likely indicates competition for the same binding sites. The effect of lupanine is dose-dependent but importantly even with the high concentration of 1 mmol/L only ~50% of the current are blocked. This observation explains why lupanine does not depolarize V
m to the threshold for opening of Ca
v channels in the presence of low glucose concentrations. Obviously, the remaining K
+ conductance is large enough to maintain V
m hyperpolarized. However, when beta cells are already electrically active,
i.e., when membrane resistance is high due to glucose- or arginine-mediated stimulation, further reduction of K
ATP current can potentiate membrane depolarization resulting in a rise in action potential frequency. With respect to our
in vivo data and the studies with humans these characteristics may explain why the effect of lupanine increases with rising hyperglycemia. Another possibility is that lupanine might exert additional effects on the amplifying pathway. The fact that lupanine does not act as a primary stimulus is very important regarding its potential as an antidiabetic drug. It is well known that hypoglycemia induced by K
ATP channel inhibitors like sulfonylureas or glinides and the associated complications are limiting the clinical value of these drugs [
13,
14]. K
ATP channel inhibition has also been shown for the lupin alkaloid sparteine in the insulin-secreting cell line HIT-T15 and in murine beta cells [
10,
15]. In contrast to lupanine the inhibitory effect of sparteine was much more pronounced and large enough to induce insulin secretion at micromolar concentrations in unstimulated islets [
10]. For skeletal muscle cells, it has been shown that lupanine inhibits voltage-dependent Na
+ channels (Na
v channels) at concentrations similar to those of our study [
7]. However, as in rodent beta cells, glucose-induced action potentials are solely carried by Ca
2+ [
16] we can exclude any contribution of interactions with Na
v channels. Our preincubation experiments show that after prolonged exposure to lupanine inhibition of K
ATP channels persists even in the absence of the alkaloid. This most likely indicates membrane enrichment of the compound. Importantly, the extent of inhibition is similar to the acute effect of lupanine and basal insulin secretion is not affected. These observations suggest that this degree of reduction in K
ATP current represents a saturated condition that is not cumulative during increasing exposure time.
Of note, lupanine is not only acting on the stimulus-secretion cascade but also affects the expression of insulin mRNA. The elevated gene transcription level may contribute to a better secretory response especially during long-term treatment with the drug. At present, we cannot explain the small but significant alterations in basal insulin release observed with 0.05 and 1 mmol/L lupanine, respectively. As lupanine does not influence V
m in the presence of 0.5–6 mmol/L glucose in a concentration range from 0.05 to 1 mmol/L these changes are not linked to a V
m-dependent pathway and may result from direct interference with the exocytotic machinery. Additional effects not-related to interactions with ion channels have also been suggested to be involved in the action of sparteine on insulin release [
10].
Lupins gain increasing significance as functional foods. When studying lupin components one must be aware of its limitations with respect to therapeutic use. The neurotoxic quinolizidine alkaloids are known for its hazardous potential inducing trembling, seizures and disturbance of blood pressure regulation [
17,
18,
19]. Case reports also point to anticholinergic symptoms after ingestion of bitter lupin flour [
20]. In the majority of studies adverse effects are most prominent for sparteine [
18,
21]. In a feeding study with
Lupinus angustifolius [
22], no toxic effects were observed in rats over a period of 90 days. The main alkaloids of this variety were identified as lupanine and 13-hydroxylupanine and the daily intake was calculated to range from 400 to 500 mg alkaloid/kg BW. For isolated lupanine toxicity studies revealed an LD
50 of 174–177 mg/kg BW (intraperitoneal application) and 1664 mg/kg BW (oral intake) in mice and rats [
19],
i.e., at a concentration ~80-fold higher compared to the 20 mg/kg BW used in our
in vivo experiment. Human studies emphasizing the potential of lupin raw material for improvement of glycemic control do not point to drug-related adverse effects [
2,
3] but mostly sweet lupins with low alkaloid contents were used. However, as different
Lupinus species vary substantially with respect to their alkaloid content and alkaloid composition [
21,
23], standardization is very important especially considering the use of lupin-based products to support control of glucose homeostasis.
To avoid unforeseeable effects pure lupanine or lupanine-enriched supplements might be more appropriate. Importantly, analysis of protein isolates obtained by different methods from different lupin varieties (
L. albus and
L. angustifolius) showed that these isolates all contained low amounts (<0.002%) of alkaloids [
24]. As it is known that the protein γ-conglutin improves glucose transport [
25] and elevates pancreatic insulin content [
26] it is tempting to speculate whether combined application of lupanine and γ-conglutin act in synergy. Our
in vivo data clearly show that lupanine improves glucose tolerance in response to glucose ingestion. Further studies are needed to investigate whether lupanine interacts with the incretine system. In agreement with the results obtained for stimulation of insulin release the effects of lupanine get important when blood glucose levels are pathologically high. Bobkiewicz-Kozlowska [
4] reported that approximately the same concentration of lupanine as used in our present study did not lower blood glucose in non-diabetic or STZ-diabetic rats without any glucose challenge over a period of 2 h. This is in agreement with our data where blood glucose at the beginning of the oGTT (
i.e., 30 min after lupanine-injection) was not reduced and only a tendency to improved glucose tolerance was observed in the oGTT with non-diabetic animals.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






