
Hitting the Sweet Spot: Glycans as Targets of Fungal Defense Effector Proteins

Abstract
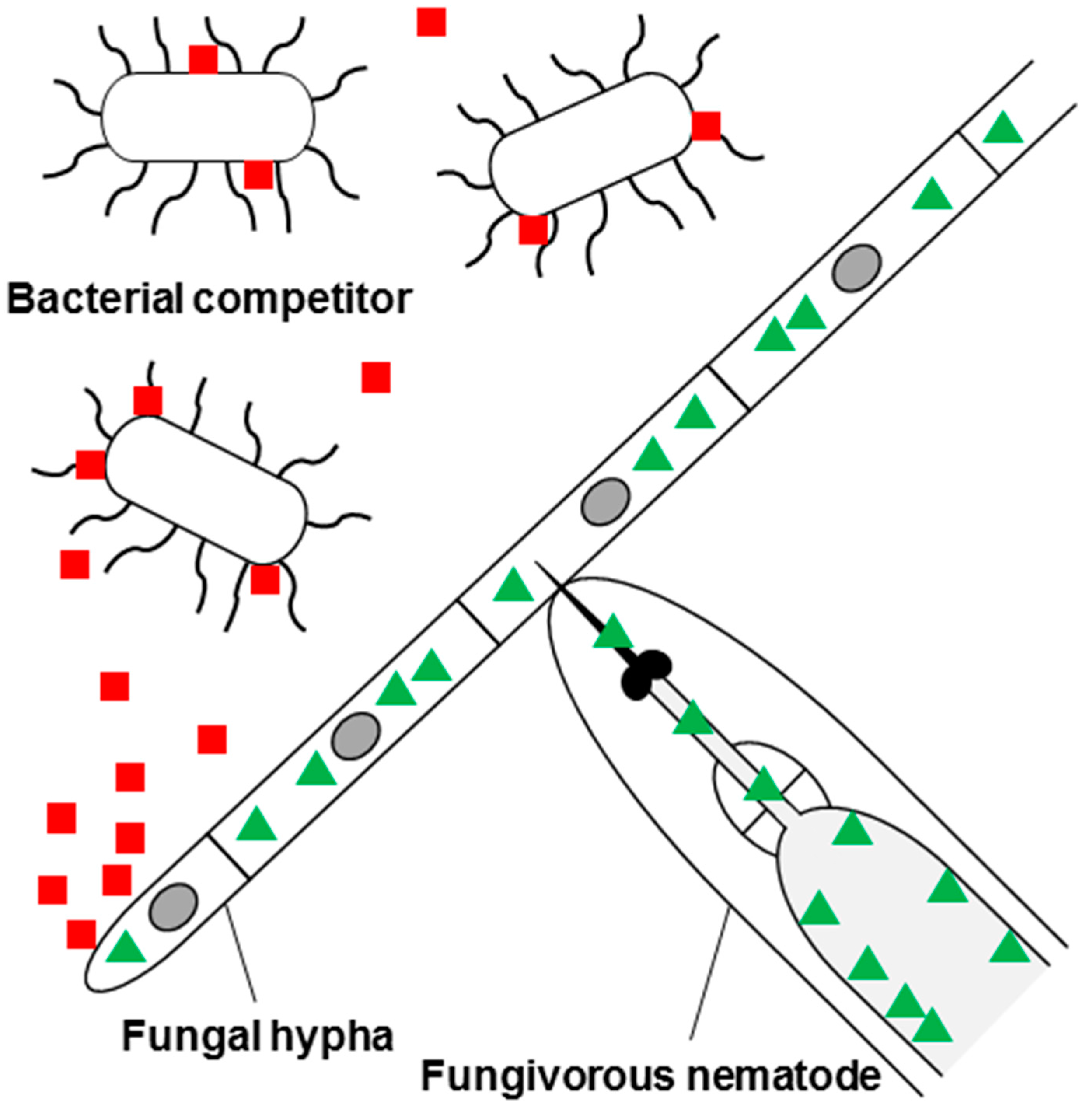
:1. Introduction and Scope of This Review

2. Fungal Defense Effector Proteins Targeting Fungal Glycoepitopes
2.1. Defensins
2.2. LysM Effector Proteins
2.3. Thaumatin-Like Proteins
2.4. Lectins

{kind=link}
{kind=link}
| Protein | Type | Producing Fungus | Target Organism | Target Polysaccharide/Glycoconjugate | Target Glycoepitope (in vitro/in vivo) | References | |
|---|---|---|---|---|---|---|---|
| AFP | Non-classical defensin-like | Aspergillus giganteus | Filamentous fungi | Chitin Glucosylceramide | (GlcNAc-β1,4)n-GlcNAc Glc | [23] | |
| PAF | Non-classical defensin-like | Penicillium chrysogenum | Filamentous fungi | Chitin? Glucosylceramide? | (GlcNAc-β1,4)n-GlcNAc? Glc? | [24] | |
| BP | Non-classical defensin-like | Penicillium brevicompactum | Filamentous fungi | Chitin? Glucosylceramide? | (GlcNAc-β1,4)n-GlcNAc? Glc? | [25] | |
| LysM-effector | Various | Filamentous fungi Gram-positive bacteria? | Chitin Peptidoglycan? | (GlcNAc-β1,4)n-GlcNAc (MurNAc-β1,4-GlcNAc)n? | [33] | ||
| Thaumatin-like | Various | Filamentous fungi | β1,3-glucans | (Glc-β1,3)n-Glc | [39] | ||
| AAL | Hololectin | Aleuria aurantia | Zygomycetes Insects Nematodes Amoeba | Fucose-containing polysaccharides Fucose-containing N- and/or O-glycans? | Fuc-α1,x-X | [44,47,48,49] | |
| Plectasin | Csαβ defensin-like | Pseudoplectania nigrella | Gram-positive bacteria | Lipid II | ? | [6] | |
| Eurocin | Csαβ defensin-like | Eurotium amstelodami | Gram-positive bacteria | Lipid II | ? | [50] | |
| Micasin | Csαβ defensin-like | Microsporum canis | Gram-positive and -negative bacteria | Lipid II | ? | [51] | |
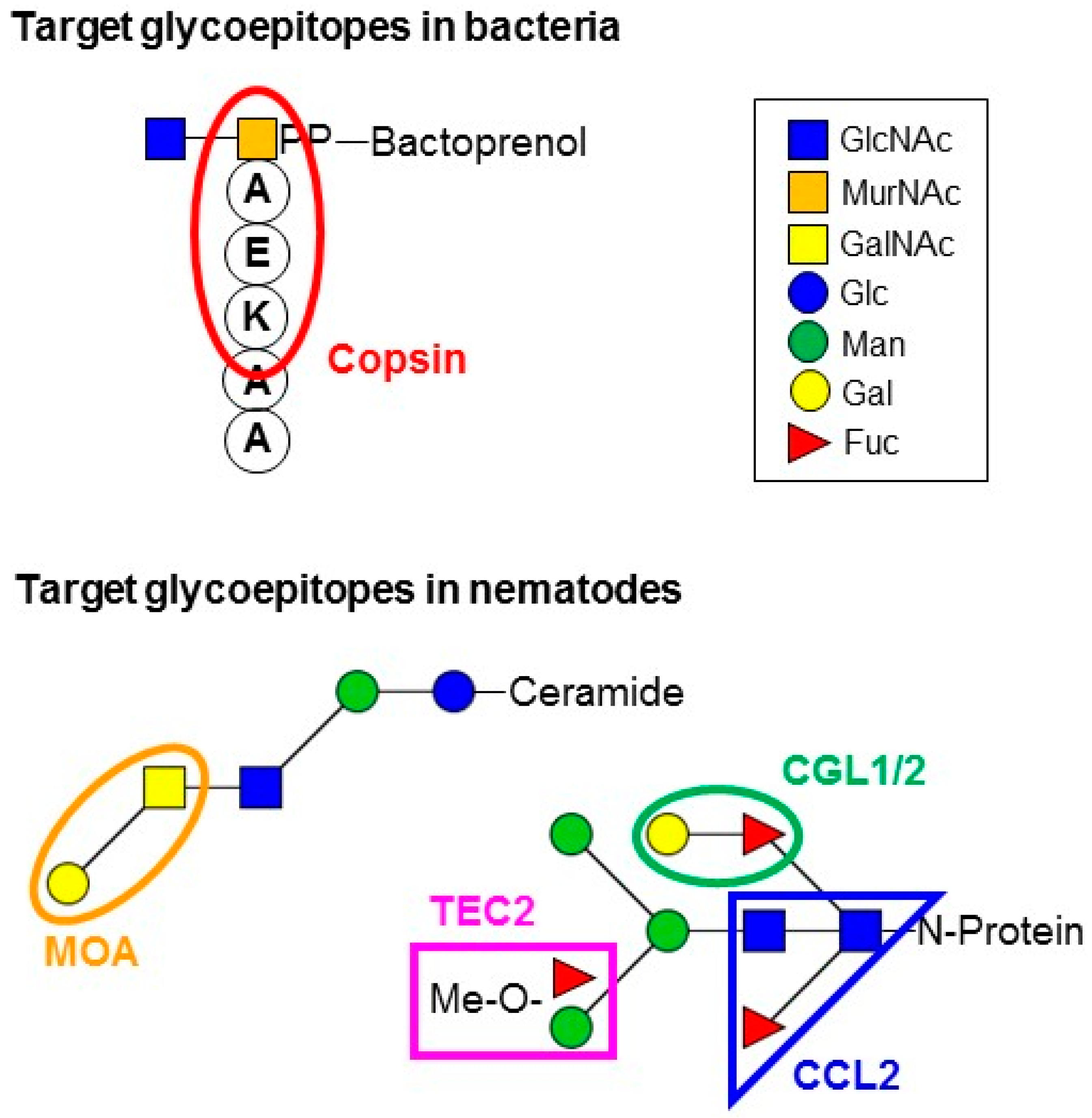
| Copsin | Csαβ defensin-like | Coprinopsis cinerea | Gram-positive bacteria | Lipid II | ? | [52] | |
| GH24-lysozyme | Various | Gram-positive bacteria | Peptidoglycan | (MurNAc-β1,4-GlcNAc)n | |||
| GH25-lysozyme | Chalaropsis sp. | Gram-positive bacteria | Peptidoglycan | (MurNAc-β1,4-GlcNAc)n | [53] | ||
| Ceratoplatanin | Various | Filamentous fungi? Gram-positive bacteria? | Chitin? Peptidoglycan? | (GlcNAc-β1,4)n-GlcNAc? (MurNAc-β1,4-GlcNAc)n? | [54,55] | ||
| XCL | Hololectin | Xerocomus chrysenteron | Insects Nematodes | N- and/or O-glycans | Gal-β1,3-GalNAc and GlcNAc-β1,2-Man | [47,56,57] | |
| TAP1 | Hololectin | Sordaria macrospora | Insects Nematodes Amoeba | O-glycans? | Gal-β1,3-GalNAc | [47,57] | |
| CCL2 | Hololectin | Coprinopsis cinerea | Insects Nematodes | N-glycan core | GlcNAc-β1,4(Fuc-α1,3)-GlcNAc | [19,57] | |
| CNL | Hololectin | Clitocybe nebularis | Mammalian cells Nematodes Amoeba | O-glycans? | GalNAc | [47,58] | |
| MPL | Hololectin | Macrolepiota procera | Nematodes | N- and/or O-glycans? | Gal-β1,4-GlcNAc | [59] | |
| SSA | Hololectin | Sclerotinia sclerotiorum | Insects Amoeba | N- and/or O-glycans? | GalNAc/Gal | [47,60] | |
| RSA | Hololectin | Rhizoctonia solani | Insects | N- and/or O-glycans? | GalNAc/Gal | [61,62,63] | |
| CGL1/2 | Hololectin | Coprinopsis cinerea | Insects Nematodes Amoeba | N- and/or O-glycans? N-glycan core N- and/or O-glycans? | Gal-β1,4-Glc Gal-β1,4-GlcNAc Gal-β1,4-Fuc | [47,57] | |
| Tectonin2 | Hololectin | Laccaria bicolor | Nematodes | N-glycan antenna | 2-O-Me-Fuc/3-O-Me-Man | [20] | |
| MOA | Chimerolectin | Marasmius oreades | Nematodes | Glycosphingolipids | Gal-α1,3-GalNAc | [64,65,66] | |
| LSL | Chimerolectin | Laetiporus sulphureus | Mammalian cells | ? | Gal-β1,4-Glc Gal-β1,4-GlcNAc | [67,68] | |
3. Fungal Defense Effector Proteins Targeting Bacterial Glycoepitopes
3.1. Defensins
3.2. Lysozymes

3.3. Ceratoplatanins
3.4. Lectins
4. Fungal Defense Effector Proteins Targeting Metazoan Glycoepitopes
4.1. Hololectins
4.2. Chimerolectins
5. Mechanism of Toxicities Mediated by Glycan-Protein Interaction
6. Regulation of Fungal Defense Protein Biosynthesis
7. Evolution of Fungal Defense Proteins
8. Conclusions and Outlook
Acknowledgments
Conflicts of Interest
References
- Stajich, J.E.; Berbee, M.L.; Blackwell, M.; Hibbett, D.S.; James, T.Y.; Spatafora, J.W.; Taylor, J.W. The fungi. Curr. Biol. 2009, 19, R840–R845. [Google Scholar] [CrossRef] [PubMed]
- Bills, G.F.; Gloer, J.B.; An, Z. Coprophilous fungi: Antibiotic discovery and functions in an underexplored arena of microbial defensive mutualism. Curr. Opin. Microbiol. 2013, 16, 549–565. [Google Scholar] [CrossRef] [PubMed]
- Ruess, L.; Lussenhop, J. Trophic interactions of fungi and animals. In The Fungal Community: Its Organization and Role in the Ecosystems; Dighton, J., White, J.F., Oudemans, P., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 581–598. [Google Scholar]
- Spiteller, P. Chemical defence strategies of higher fungi. Chemistry 2008, 14, 9100–9110. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism-from biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Mygind, P.H.; Fischer, R.L.; Schnorr, K.M.; Hansen, M.T.; Sonksen, C.P.; Ludvigsen, S.; Raventos, D.; Buskov, S.; Christensen, B.; De Maria, L.; et al. Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 2005, 437, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Bills, G.; Li, Y.; Chen, L.; Yue, Q.; Niu, X.M.; An, Z. New insights into the echinocandins and other fungal non-ribosomal peptides and peptaibiotics. Nat. Prod. Rep. 2014, 31, 1348–1375. [Google Scholar] [CrossRef] [PubMed]
- Umemura, M.; Nagano, N.; Koike, H.; Kawano, J.; Ishii, T.; Miyamura, Y.; Kikuchi, M.; Tamano, K.; Yu, J.; Shin-ya, K.; et al. Characterization of the biosynthetic gene cluster for the ribosomally synthesized cyclic peptide Ustiloxin B in Aspergillus flavus. Fungal. Genet. Biol. 2014, 68, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Trigueros, V.; Paquereau, L.; Chavant, L.; Fournier, D. Proteins as active compounds involved in insecticidal activity of mushroom fruitbodies. J. Econ. Entomol. 2002, 95, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, M.A.; Westerlaken, I.; Leeflang, C.; Kerkman, R.; Bovenberg, R.A. Functional characterization of the penicillin biosynthetic gene cluster of Penicillium chrysogenum Wisconsin 54–1255. Fungal. Genet. Biol. 2007, 44, 830–844. [Google Scholar] [CrossRef] [PubMed]
- Hallen, H.E.; Luo, H.; Scott-Craig, J.S.; Walton, J.D. Gene family encoding the major toxins of lethal Amanita mushrooms. Proc. Natl. Acad. Sci. USA 2007, 104, 19097–19101. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Uehara, T.; Bernhardt, T.G. Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery. Cell 2014, 159, 1300–1311. [Google Scholar] [CrossRef] [PubMed]
- Yocum, R.R.; Waxman, D.J.; Rasmussen, J.R.; Strominger, J.L. Mechanism of penicillin action: Penicillin and substrate bind covalently to the same active site serine in two bacterial D-alanine carboxypeptidases. Proc. Natl. Acad. Sci. USA 1979, 76, 2730–2734. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, D.A.; Cramer, P.; Kornberg, R.D. Structural basis of transcription: Alpha-amanitin-RNA polymerase II cocrystal at 2.8 Å resolution. Proc. Natl. Acad. Sci. USA 2002, 99, 1218–1222. [Google Scholar] [CrossRef] [PubMed]
- Horgen, P.A.; Vaisius, A.C.; Ammirati, J.F. The insensitivity of mushroom nuclear RNA polymerase activity to inhibition by amatoxins. Arch. Microbiol. 1978, 118, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Boehm, T. Evolution of vertebrate immunity. Curr. Biol. 2012, 22, R722–R732. [Google Scholar] [CrossRef] [PubMed]
- Peumans, W.J.; Van Damme, E.J. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Butschi, A.; Titz, A.; Walti, M.A.; Olieric, V.; Paschinger, K.; Nobauer, K.; Guo, X.; Seeberger, P.H.; Wilson, I.B.; Aebi, M.; et al. Caenorhabditis elegans N-glycan core beta-galactoside confers sensitivity towards nematotoxic fungal galectin CGL2. PLoS Pathog. 2010, 6, e1000717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, M.; Bleuler-Martinez, S.; Butschi, A.; Walti, M.A.; Egloff, P.; Stutz, K.; Yan, S.; Wilson, I.B.; Hengartner, M.O.; Aebi, M.; et al. Plasticity of the beta-trefoil protein fold in the recognition and control of invertebrate predators and parasites by a fungal defence system. PLoS Pathog. 2012, 8, e1002706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlschlager, T.; Butschi, A.; Grassi, P.; Sutov, G.; Gauss, R.; Hauck, D.; Schmieder, S.S.; Knobel, M.; Titz, A.; Dell, A.; et al. Methylated glycans as conserved targets of animal and fungal innate defense. Proc. Natl. Acad. Sci. USA 2014, 111, E2787–E2796. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Cummings, R.D.; Doering, T.L. Fungi. In Essentials in Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Hagen, S.; Marx, F.; Ram, A.F.; Meyer, V. The antifungal protein AFP from Aspergillus giganteus inhibits chitin synthesis in sensitive fungi. Appl. Environ. Microbiol. 2007, 73, 2128–2134. [Google Scholar] [CrossRef] [PubMed]
- Batta, G.; Barna, T.; Gaspari, Z.; Sandor, S.; Kover, K.E.; Binder, U.; Sarg, B.; Kaiserer, L.; Chhillar, A.K.; Eigentler, A.; et al. Functional aspects of the solution structure and dynamics of PAF—a highly-stable antifungal protein from Penicillium chrysogenum. FEBS J. 2009, 276, 2875–2890. [Google Scholar] [CrossRef] [PubMed]
- Seibold, M.; Wolschann, P.; Bodevin, S.; Olsen, O. Properties of the bubble protein, a defensin and an abundant component of a fungal exudate. Peptides 2011, 32, 1989–1995. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.M.; Goncalves, S.; Santos, N.C. Defensins: Antifungal lessons from eukaryotes. Front. Microbiol. 2014, 5, 97. [Google Scholar] [PubMed]
- Wilmes, M.; Sahl, H.G. Defensin-based anti-infective strategies. Int. J. Med. Microbiol. 2014, 304, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Unifying themes in host defence effector polypeptides. Nat. Rev. Microbiol. 2007, 5, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V. A small protein that fights fungi: AFP as a new promising antifungal agent of biotechnological value. Appl. Microbiol. Biotechnol. 2008, 78, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Aerts, A.M.; Francois, I.E.J.A.; Cammue, B.P.A.; Thevissen, K. The mode of antifungal action of plant, insect and human defensins. Cell. Mol. Life Sci. 2008, 65, 2069–2079. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, N.; Sigl, C.; Zadra, I.; Pocsi, I.; Marx, F. The PAF gene product modulates asexual development in Penicillium chrysogenum. J. Basic. Microbiol. 2011, 51, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Eigentler, A.; Pocsi, I.; Marx, F. The anisin1 gene encodes a defensin-like protein and supports the fitness of Aspergillus nidulans. Arch. Microbiol. 2012, 194, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Akcapinar, G.B.; Kappel, L.; Sezerman, O.U.; Seidl-Seiboth, V. Molecular diversity of LysM carbohydrate-binding motifs in fungi. Curr. Genet. 2015, 61, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, S.; Dellarole, M.; Baxter, N.J.; Rouget, J.B.; Dimitrov, J.D.; Wang, N.; Fujimoto, Y.; Hounslow, A.M.; Lacroix-Desmazes, S.; Fukase, K.; et al. Molecular basis for bacterial peptidoglycan recognition by LysM domains. Nat. Commun. 2014, 5, 4269. [Google Scholar] [CrossRef] [PubMed]
- Kombrink, A.; Thomma, B.P. LysM effectors: Secreted proteins supporting fungal life. PLoS Pathog. 2013, 9, e1003769. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Sturrock, R.; Ekramoddoullah, A.K. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell. Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Shatters, R.G., Jr.; Boykin, L.M.; Lapointe, S.L.; Hunter, W.B.; Weathersbee, A.A., 3rd. Phylogenetic and structural relationships of the PR5 gene family reveal an ancient multigene family conserved in plants and select animal taxa. J. Mol. Evol. 2006, 63, 12–29. [Google Scholar] [CrossRef] [PubMed]
- Trudel, J.; Grenier, J.; Potvin, C.; Asselin, A. Several thaumatin-like proteins bind to beta-1,3-glucans. Plant Physiol. 1998, 118, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Grenier, J.; Potvin, C.; Asselin, A. Some fungi express beta-1,3-glucanases similar to thaumatin-like proteins. Mycologia 2000, 92, 841–848. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Watanabe, H.; Nagai, M.; Nakade, K.; Takahashi, M.; Sato, T. Lentinula edodes tlg1 encodes a thaumatin-like protein that is involved in lentinan degradation and fruiting body senescence. Plant Physiol. 2006, 141, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, T.; Hatano, K.I.; Miyauchi, Y.; Suwa, Y.I.; Sawano, Y.; Tanokura, M. A secreted protein of the plant-specific DUF26 family functions as a mannose-binding lectin that exhibits antifungal activity. Plant Physiol. 2014, 166, 766–778. [Google Scholar] [CrossRef] [PubMed]
- Regente, M.; Taveira, G.B.; Pinedo, M.; Elizalde, M.M.; Ticchi, A.J.; Diz, M.S.; Carvalho, A.O.; de la Canal, L.; Gomes, V.M. A sunflower lectin with antifungal properties and putative medical mycology applications. Curr. Microbiol. 2014, 69, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Kohatsu, L.; Hsu, D.K.; Jegalian, A.G.; Liu, F.T.; Baum, L.G. Galectin-3 induces death of Candida species expressing specific beta-1,2-linked mannans. J. Immunol. 2006, 177, 4718–4726. [Google Scholar] [CrossRef] [PubMed]
- Amano, K.; Katayama, H.; Saito, A.; Ando, A.; Nagata, Y. Aleuria aurantia lectin exhibits antifungal activity against mucor racemosus. Biosci. Biotechnol. Biochem. 2012, 76, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Cabib, E.; Arroyo, J. How carbohydrates sculpt cells: Chemical control of morphogenesis in the yeast cell wall. Nat. Rev. Microbiol. 2013, 11, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Free, S.J. Fungal cell wall organization and biosynthesis. Adv. Genet. 2013, 81, 33–82. [Google Scholar] [PubMed]
- Bleuler-Martinez, S.; Butschi, A.; Garbani, M.; Walti, M.A.; Wohlschlager, T.; Potthoff, E.; Sabotic, J.; Pohleven, J.; Luthy, P.; Hengartner, M.O.; et al. A lectin-mediated resistance of higher fungi against predators and parasites. Mol. Ecol. 2011, 20, 3056–3070. [Google Scholar] [CrossRef] [PubMed]
- Olausson, J.; Tibell, L.; Jonsson, B.H.; Pahlsson, P. Detection of a high affinity binding site in recombinant Aleuria aurantia lectin. Glycoconj. J. 2008, 25, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Wimmerova, M.; Mitchell, E.; Sanchez, J.F.; Gautier, C.; Imberty, A. Crystal structure of fungal lectin: Six-bladed beta-propeller fold and novel fucose recognition mode for Aleuria aurantia lectin. J. Biol. Chem. 2003, 278, 27059–27067. [Google Scholar] [CrossRef] [PubMed]
- Oeemig, J.S.; Lynggaard, C.; Knudsen, D.H.; Hansen, F.T.; Norgaard, K.D.; Schneider, T.; Vad, B.S.; Sandvang, D.H.; Nielsen, L.A.; Neve, S.; et al. Eurocin, a new fungal defensin: Structure, lipid binding, and its mode of action. J. Biol. Chem. 2012, 287, 42361–42372. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Gao, B.; Harvey, P.J.; Craik, D.J. Dermatophytic defensin with antiinfective potential. Proc. Natl. Acad. Sci. USA 2012, 109, 8495–8500. [Google Scholar] [CrossRef] [PubMed]
- Essig, A.; Hofmann, D.; Munch, D.; Gayathri, S.; Künzler, M.; Kallio, P.T.; Sahl, H.G.; Wider, G.; Schneider, T.; Aebi, M. Copsin, a novel peptide-based fungal antibiotic interfering with the peptidoglycan synthesis. J. Biol. Chem. 2014, 289, 34953–34964. [Google Scholar] [CrossRef] [PubMed]
- Fouche, P.B.; Hash, J.H. The N,O-diacetylmuramidase of Chalaropsis species. Identificaiton of aspartyl and glutamyl residues in the active site. J. Biol. Chem. 1978, 253, 6787–6793. [Google Scholar] [PubMed]
- Chen, H.; Kovalchuk, A.; Kerio, S.; Asiegbu, F.O. Distribution and bioinformatic analysis of the cerato-platanin protein family in dikarya. Mycologia 2013, 105, 1479–1488. [Google Scholar] [CrossRef] [PubMed]
- Frischmann, A.; Neudl, S.; Gaderer, R.; Bonazza, K.; Zach, S.; Gruber, S.; Spadiut, O.; Friedbacher, G.; Grothe, H.; Seidl-Seiboth, V. Self-assembly at air/water interfaces and carbohydrate binding properties of the small secreted protein EPL1 from the fungus Trichoderma atroviride. J. Biol. Chem. 2013, 288, 4278–4287. [Google Scholar] [CrossRef] [PubMed]
- Trigueros, V.; Lougarre, A.; Ali-Ahmed, D.; Rahbe, Y.; Guillot, J.; Chavant, L.; Fournier, D.; Paquereau, L. Xerocomus chrysenteron lectin: Identification of a new pesticidal protein. Biochim. Biophys. Acta 2003, 1621, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Bleuler-Martinez, S.; Plaza Gutierrez, D.F.; Kuenzler, M.; Aebi, M.; Joachim, A.; Razzazi-Fazeli, E.; Jantsch, V.; Geyer, R.; Wilson, I.B.; et al. Galactosylated fucose epitopes in nematodes: Increased expression in a Caenorhabditis mutant associated with altered lectin sensitivity and occurrence in parasitic species. J. Biol. Chem. 2012, 287, 28276–28290. [Google Scholar] [CrossRef] [PubMed]
- Pohleven, J.; Brzin, J.; Vrabec, L.; Leonardi, A.; Cokl, A.; Strukelj, B.; Kos, J.; Sabotic, J. Basidiomycete Clitocybe nebularis is rich in lectins with insecticidal activities. Appl. Microbiol. Biotechnol. 2011, 91, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Zurga, S.; Pohleven, J.; Renko, M.; Bleuler-Martinez, S.; Sosnowski, P.; Turk, D.; Künzler, M.; Kos, J.; Sabotic, J. A novel beta-trefoil lectin from the parasol mushroom (Macrolepiota procera) is nematotoxic. FEBS J. 2014, 281, 3489–3506. [Google Scholar] [CrossRef] [PubMed]
- Sulzenbacher, G.; Roig-Zamboni, V.; Peumans, W.J.; Rouge, P.; Van Damme, E.J.; Bourne, Y. Crystal structure of the GalNAc/Gal-specific agglutinin from the phytopathogenic ascomycete Sclerotinia sclerotiorum reveals novel adaptation of a beta-trefoil domain. J. Mol. Biol. 2010, 400, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamshou, M.; Van Damme, E.J.; Caccia, S.; Cappelle, K.; Vandenborre, G.; Ghesquiere, B.; Gevaert, K.; Smagghe, G. High entomotoxicity and mechanism of the fungal GalNAc/Gal-specific Rhizoctonia solani lectin in pest insects. J. Insect. Physiol. 2013, 59, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Hamshou, M.; Van Damme, E.J.; Smagghe, G. Entomotoxic effects of fungal lectin from Rhizoctonia solani towards Spodoptera littoralis. Fungal. Biol. 2010, 114, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Hamshou, M.; van Damme, E.J.; Vandenborre, G.; Ghesquiere, B.; Trooskens, G.; Gevaert, K.; Smagghe, G. GalNAc/Gal-binding Rhizoctonia solani agglutinin has antiproliferative activity in Drosophila melanogaster S2 cells via MAPK and JAK/STAT signaling. PLoS ONE 2012, 7, e33680. [Google Scholar] [CrossRef] [PubMed]
- Wohlschlager, T.; Butschi, A.; Zurfluh, K.; Vonesch, S.C.; Auf dem Keller, U.; Gehrig, P.; Bleuler-Martinez, S.; Hengartner, M.O.; Aebi, M.; Künzler, M. Nematotoxicity of Marasmius oreades agglutinin (MOA) depends on glycolipid-binding and cysteine protease activity. J. Biol. Chem. 2011, 286, 30337–30343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordara, G.; Egge-Jacobsen, W.; Johansen, H.T.; Winter, H.C.; Goldstein, I.J.; Sandvig, K.; Krengel, U. Marasmius oreades agglutinin (MOA) is a chimerolectin with proteolytic activity. Biochem. Biophys. Res. Commun. 2011, 408, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Grahn, E.M.; Winter, H.C.; Tateno, H.; Goldstein, I.J.; Krengel, U. Structural characterization of a lectin from the mushroom Marasmius oreades in complex with the blood group B trisaccharide and calcium. J. Mol. Biol. 2009, 390, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Mancheno, J.M.; Tateno, H.; Goldstein, I.J.; Martinez-Ripoll, M.; Hermoso, J.A. Structural analysis of the Laetiporus sulphureus hemolytic pore-forming lectin in complex with sugars. J. Biol. Chem. 2005, 280, 17251–17259. [Google Scholar] [CrossRef]
- Angulo, I.; Acebron, I.; de las Rivas, B.; Munoz, R.; Rodriguez-Crespo, I.; Menendez, M.; Garcia, P.; Tateno, H.; Goldstein, I.J.; Perez-Agote, B.; et al. High-resolution structural insights on the sugar-recognition and fusion tag properties of a versatile beta-trefoil lectin domain from the mushroom Laetiporus sulphureus. Glycobiology 2011, 21, 1349–1361. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Cornet, B.; Bonmatin, J.M.; Hetru, C.; Hoffmann, J.A.; Ptak, M.; Vovelle, F. Refined three-dimensional solution structure of insect defensin A. Structure 1995, 3, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Raventos, D.S.; et al. Plectasin, a fungal defensin, targets the bacterial cell wall precursor lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Sahl, H.G. An oldie but a goodie-cell wall biosynthesis as antibiotic target pathway. Int. J. Med. Microbiol. 2010, 300, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Gao, B.; Zhu, S. The fungal defensin family enlarged. Pharmaceuticals 2014, 7, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhu, S. Alteration of the mode of antibacterial action of a defensin by the amino-terminal loop substitution. Biochem. Biophys. Res. Commun. 2012, 426, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Dimarcq, J.L.; Bulet, P.; Hetru, C.; Hoffmann, J. Cysteine-rich antimicrobial peptides in invertebrates. Biopolymers 1998, 47, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Callewaert, L.; Michiels, C.W. Lysozymes in the animal kingdom. J. Biosci. 2010, 35, 127–160. [Google Scholar] [CrossRef] [PubMed]
- Beintema, J.J.; Terwisscha van Scheltinga, A.C. Plant lysozymes. EXS 1996, 75, 75–86. [Google Scholar] [PubMed]
- During, K.; Porsch, P.; Mahn, A.; Brinkmann, O.; Gieffers, W. The non-enzymatic microbicidal activity of lysozymes. FEBS Lett. 1999, 449, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Matsuzaki, T.; Aoki, T. Genetic evidence that antibacterial activity of lysozyme is independent of its catalytic function. FEBS Lett. 2001, 506, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Van Herreweghe, J.M.; Michiels, C.W. Invertebrate lysozymes: Diversity and distribution, molecular mechanism and in vivo function. J. Biosci. 2012, 37, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Vanscheltinga, A.C.T.; Kalk, K.H.; Beintema, J.J.; Dijkstra, B.W. Crystal-structures of hevamine, a plant defense protein with chitinase and lysozyme activity, and its complex with an inhibitor. Structure 1994, 2, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.D.; Prosser, B.A.; Asher, R.A. A bacteriolytic muramidase from the basidiomycete Schizophyllum commune. J. Gen. Microbiol. 1990, 136, 2267–2273. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, S.P.; Fermor, T.R.; Wood, D.A. Production and detection of muramidase and acetylglucosaminidase from Agaricus bisporus. Lett. Appl. Microbiol. 1997, 25, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Baccelli, I. Cerato-platanin family proteins: One function for multiple biological roles? Front. Plant Sci. 2014, 5, 769. [Google Scholar] [PubMed]
- Pazzagli, L.; Seidl-Seiboth, V.; Barsottini, M.; Vargas, W.A.; Scala, A.; Mukherjee, P.K. Cerato-platanins: Elicitors and effectors. Plant Sci. 2014, 228C, 79–87. [Google Scholar] [CrossRef]
- De Oliveira, A.L.; Gallo, M.; Pazzagli, L.; Benedetti, C.E.; Cappugi, G.; Scala, A.; Pantera, B.; Spisni, A.; Pertinhez, T.A.; Cicero, D.O. The structure of the elicitor cerato-platanin (cp), the first member of the cp fungal protein family, reveals a double psibeta-barrel fold and carbohydrate binding. J. Biol. Chem. 2011, 286, 17560–17568. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Holst, O.; Hardt, W.D. The bactericidal activity of the c-type lectin RegIIIbeta against Gram-negative bacteria involves binding to lipid A. J. Biol. Chem. 2012, 287, 34844–34855. [Google Scholar] [CrossRef] [PubMed]
- Lehotzky, R.E.; Partch, C.L.; Mukherjee, S.; Cash, H.L.; Goldman, W.E.; Gardner, K.H.; Hooper, L.V. Molecular basis for peptidoglycan recognition by a bactericidal lectin. Proc. Natl. Acad. Sci. USA 2010, 107, 7722–7727. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Arthur, C.M.; McBride, R.; Berger, O.; Razi, N.; Heimburg-Molinaro, J.; Rodrigues, L.C.; Gourdine, J.P.; Noll, A.J.; von Gunten, S.; et al. Microbial glycan microarrays define key features of host-microbial interactions. Nat. Chem. Biol. 2014, 10, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.K.; Huang, Y.T.; Lee, Y.C.; Low, E.L.; Chiu, C.H.; Chen, S.L.; Mao, L.C.; Chang, M.D. A recombinant horseshoe crab plasma lectin recognizes specific pathogen-associated molecular patterns of bacteria through rhamnose. PLoS ONE 2014, 9, e115296. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Arthur, C.M.; Dias-Baruffi, M.; Rodrigues, L.C.; Gourdine, J.P.; Heimburg-Molinaro, J.; Ju, T.; Molinaro, R.J.; Rivera-Marrero, C.; Xia, B.; et al. Innate immune lectins kill bacteria expressing blood group antigen. Nat. Med. 2010, 16, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Hardt, W.D. Outer membrane permeabilization is an essential step in the killing of Gram-negative bacteria by the lectin RegIIIbeta. PLoS ONE 2013, 8, e69901. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Zheng, H.; Derebe, M.G.; Callenberg, K.M.; Partch, C.L.; Rollins, D.; Propheter, D.C.; Rizo, J.; Grabe, M.; Jiang, Q.X.; et al. Antibacterial membrane attack by a pore-forming intestinal c-type lectin. Nature 2014, 505, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Künzler, M.; Bleuler-Martinez, S.; Butschi, A.; Garbani, M.; Luthy, P.; Hengartner, M.O.; Aebi, M. Biotoxicity assays for fruiting body lectins and other cytoplasmic proteins. Methods Enzymol. 2010, 480, 141–150. [Google Scholar] [PubMed]
- Guillot, J.; Konska, G. Lectins in higher fungi. Biochem. Syst. Ecol. 1997, 25, 203–230. [Google Scholar] [CrossRef]
- Goldstein, I.J.; Winter, H.C. Mushroom Lectins. In Comprehensive Glycoscience: From Chemistry to Systems Biology; Kamerling, J.P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2007; Volume 3. [Google Scholar]
- Wang, H.; Ng, T.B.; Ooi, V.E. Lectins from mushrooms. Mycol. Res. 1998, 102, 897–906. [Google Scholar] [CrossRef]
- Kawagishi, H. Mushroom lectins. Food Rev. Int. 1995, 11, 63–68. [Google Scholar] [CrossRef]
- Varrot, A.; Basheer, S.M.; Imberty, A. Fungal lectins: Structure, function and potential applications. Curr. Opin. Struct. Biol. 2013, 23, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Birck, C.; Damian, L.; Marty-Detraves, C.; Lougarre, A.; Schulze-Briese, C.; Koehl, P.; Fournier, D.; Paquereau, L.; Samama, J.P. A new lectin family with structure similarity to actinoporins revealed by the crystal structure of Xerocomus chrysenteron lectin XCL. J. Mol. Biol. 2004, 344, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Paschinger, K.; Razzazi-Fazeli, E.; Furukawa, K.; Wilson, I.B. Presence of galactosylated core fucose on N-glycans in the planaria Dugesia japonica. J. Mass Spectrom. 2011, 46, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, E. Methylation—An uncommon modification of glycans. Biol. Chem. 2012, 393, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Paschinger, K.; Rendic, D.; Wilson, I.B. Revealing the anti-HRP epitope in Drosophila and Caenorhabditis. Glycoconj. J. 2009, 26, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Hayama, K.; Hirabayashi, J.; Kasai, K. Caenorhabditis elegans N-glycans containing a Gal-Fuc disaccharide unit linked to the innermost GlcNAc residue are recognized by C. elegans galectin lec-6. Glycobiology 2008, 18, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Grahn, E.; Askarieh, G.; Holmner, A.; Tateno, H.; Winter, H.C.; Goldstein, I.J.; Krengel, U. Crystal structure of the Marasmius oreades mushroom lectin in complex with a xenotransplantation epitope. J. Mol. Biol. 2007, 369, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Kadirvelraj, R.; Grant, O.C.; Goldstein, I.J.; Winter, H.C.; Tateno, H.; Fadda, E.; Woods, R.J. Structure and binding analysis of Polyporus squamosus lectin in complex with the Neu5Acα2-6Galβ1-4GlcNAc human-type influenza receptor. Glycobiology 2011, 21, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Mancheno, J.M.; Tateno, H.; Sher, D.; Goldstein, I.J. Laetiporus sulphureus lectin and aerolysin protein family. Adv. Exp. Med. Biol. 2010, 677, 67–80. [Google Scholar] [PubMed]
- Szczesny, P.; Iacovache, I.; Muszewska, A.; Ginalski, K.; van der Goot, F.G.; Grynberg, M. Extending the aerolysin family: From bacteria to vertebrates. PLoS ONE 2011, 6, e20349. [Google Scholar] [CrossRef] [PubMed]
- Douglas, L.M.; Konopka, J.B. Fungal membrane organization: The eisosome concept. Annu. Rev. Microbiol. 2014, 68, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Pohleven, J.; Renko, M.; Magister, S.; Smith, D.F.; Künzler, M.; Strukelj, B.; Turk, D.; Kos, J.; Sabotic, J. Bivalent carbohydrate binding is required for biological activity of Clitocybe nebularis lectin (CNL), the N,N'-diacetyllactosediamine (GalNAcβ1–4GlcNAc, LacdiNAc)-specific lectin from basidiomycete C. nebularis. J. Biol. Chem. 2012, 287, 10602–10612. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, G.A.; Toscano, M.A.; Jackson, S.S.; Vasta, G.R. Functions of cell surface galectin-glycoprotein lattices. Curr. Opin. Struct. Biol. 2007, 17, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Boscher, C.; Dennis, J.W.; Nabi, I.R. Glycosylation, galectins and cellular signaling. Curr. Opin. Cell Biol. 2011, 23, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.W.; Nabi, I.R.; Demetriou, M. Metabolism, cell surface organization, and disease. Cell 2009, 139, 1229–1241. [Google Scholar] [CrossRef] [PubMed]
- Sacchettini, J.C.; Baum, L.G.; Brewer, C.F. Multivalent protein-carbohydrate interactions. A new paradigm for supermolecular assembly and signal transduction. Biochemistry 2001, 40, 3009–3015. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.K.; Ortega, B.; Kurosu, H.; Rosenblatt, K.P.; Kuro, O.M.; Huang, C.L. Removal of sialic acid involving Klotho causes cell-surface retention of TRPV5 channel via binding to galectin-1. Proc. Natl. Acad. Sci. USA 2008, 105, 9805–9810. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayan, R.; Wunder, C.; Becken, U.; Howes, M.T.; Benzing, C.; Arumugam, S.; Sales, S.; Ariotti, N.; Chambon, V.; Lamaze, C.; et al. Galectin-3 drives glycosphingolipid-dependent biogenesis of clathrin-independent carriers. Nat. Cell. Biol. 2014, 16, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.C.; Lord, J.M.; Roberts, L.M.; Johannes, L. Glycosphingolipids as toxin receptors. Semin. Cell Dev. Biol. 2004, 15, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Meldau, S.; Erb, M.; Baldwin, I.T. Defence on demand: Mechanisms behind optimal defence patterns. Ann. Bot. 2012, 110, 1503–1514. [Google Scholar] [CrossRef] [PubMed]
- Plaza, D.F.; Lin, C.W.; van der Velden, N.S.; Aebi, M.; Künzler, M. Comparative transcriptomics of the model mushroom Coprinopsis cinerea reveals tissue-specific armories and a conserved circuitry for sexual development. BMC Genomics 2014, 15, 492. [Google Scholar] [CrossRef] [PubMed]
- Schroeckh, V.; Scherlach, K.; Nutzmann, H.W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [PubMed]
- Rohlfs, M.; Albert, M.; Keller, N.P.; Kempken, F. Secondary chemicals protect mould from fungivory. Biol. Lett. 2007, 3, 523–525. [Google Scholar] [CrossRef] [PubMed]
- Doll, K.; Chatterjee, S.; Scheu, S.; Karlovsky, P.; Rohlfs, M. Fungal metabolic plasticity and sexual development mediate induced resistance to arthropod fungivory. Proc. Biol. Sci. 2013, 280, 20131219. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.L.; Lee, R.T.; Fang, H.M.; Wang, Y.M.; Li, R.; Zou, H.; Zhu, Y.; Wang, Y. Bacterial peptidoglycan triggers Candida albicans hyphal growth by directly activating the adenylyl cyclase Cyr1p. Cell Host Microbe 2008, 4, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Svahn, K.S.; Goransson, U.; Chryssanthou, E.; Olsen, B.; Sjolin, J.; Stromstedt, A.A. Induction of gliotoxin secretion in Aspergillus fumigatus by bacteria-associated molecules. PLoS ONE 2014, 9, e93685. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.P.; Mahanti, P.; Schroeder, F.C.; Sternberg, P.W. Nematode-trapping fungi eavesdrop on nematode pheromones. Curr. Biol. 2013, 23, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Nutzmann, H.W.; Reyes-Dominguez, Y.; Scherlach, K.; Schroeckh, V.; Horn, F.; Gacek, A.; Schumann, J.; Hertweck, C.; Strauss, J.; Brakhage, A.A. Bacteria-induced natural product formation in the fungus Aspergillus nidulans requires Saga/Ada-mediated histone acetylation. Proc. Natl. Acad. Sci. USA 2011, 108, 14282–14287. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.N.; Boulianne, R.P.; Charlton, S.; Farrell, E.M.; Sucher, A.; Lu, B.C. Fungal galectins, sequence and specificity of two isolectins from Coprinus cinereus. J. Biol. Chem. 1997, 272, 1514–1521. [Google Scholar] [CrossRef] [PubMed]
- Walti, M.A.; Walser, P.J.; Thore, S.; Grunler, A.; Bednar, M.; Künzler, M.; Aebi, M. Structural basis for chitotetraose coordination by CGL3, a novel galectin-related protein from Coprinopsis cinerea. J. Mol. Biol. 2008, 379, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, D.A. Horizontal gene transfer in fungi. FEMS Microbiol. Lett. 2012, 329, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Slot, J.C.; Rokas, A. Horizontal transfer of a large and highly toxic secondary metabolic gene cluster between fungi. Curr. Biol. 2011, 21, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J. Functional and ecological impacts of horizontal gene transfer in eukaryotes. Curr. Opin. Genet. Dev. 2009, 19, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Marcet-Houben, M.; Gabaldon, T. Acquisition of prokaryotic genes by fungal genomes. Trends Genet. 2009, 26, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Fredman, D.; Szczesny, P.; Grynberg, M.; Technau, U. Recurrent horizontal transfer of bacterial toxin genes to eukaryotes. Mol. Biol. Evol. 2012, 29, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Evolutionary forces shaping the golgi glycosylation machinery: Why cell surface glycans are universal to living cells. Cold Spring Harb. Perspect. Biol. 2011, 3, a005462. [Google Scholar] [CrossRef] [PubMed]
- Renko, M.; Sabotic, J.; Mihelic, M.; Brzin, J.; Kos, J.; Turk, D. Versatile loops in mycocypins inhibit three protease families. J. Biol. Chem. 2010, 285, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Sabotic, J.; Bleuler-Martinez, S.; Renko, M.; Avanzo Caglic, P.; Kallert, S.; Strukelj, B.; Turk, D.; Aebi, M.; Kos, J.; Künzler, M. Structural basis of trypsin inhibition and entomotoxicity of Cospin, serine protease inhibitor involved in defense of Coprinopsis cinerea fruiting bodies. J. Biol. Chem. 2012, 287, 3898–3907. [Google Scholar] [CrossRef] [PubMed]
- Zurga, S.; Pohleven, J.; Kos, J.; Sabotic, J. Beta-trefoil structure enables interactions between lectins and protease inhibitors that regulate their biological functions. J. Biochem. 2015. [Google Scholar] [CrossRef]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Künzler, M. Hitting the Sweet Spot: Glycans as Targets of Fungal Defense Effector Proteins. Molecules 2015, 20, 8144-8167. https://doi.org/10.3390/molecules20058144
Künzler M. Hitting the Sweet Spot: Glycans as Targets of Fungal Defense Effector Proteins. Molecules. 2015; 20(5):8144-8167. https://doi.org/10.3390/molecules20058144
Chicago/Turabian StyleKünzler, Markus. 2015. "Hitting the Sweet Spot: Glycans as Targets of Fungal Defense Effector Proteins" Molecules 20, no. 5: 8144-8167. https://doi.org/10.3390/molecules20058144
APA StyleKünzler, M. (2015). Hitting the Sweet Spot: Glycans as Targets of Fungal Defense Effector Proteins. Molecules, 20(5), 8144-8167. https://doi.org/10.3390/molecules20058144