
Mucin-Type O-Glycosylation in Invertebrates

Abstract
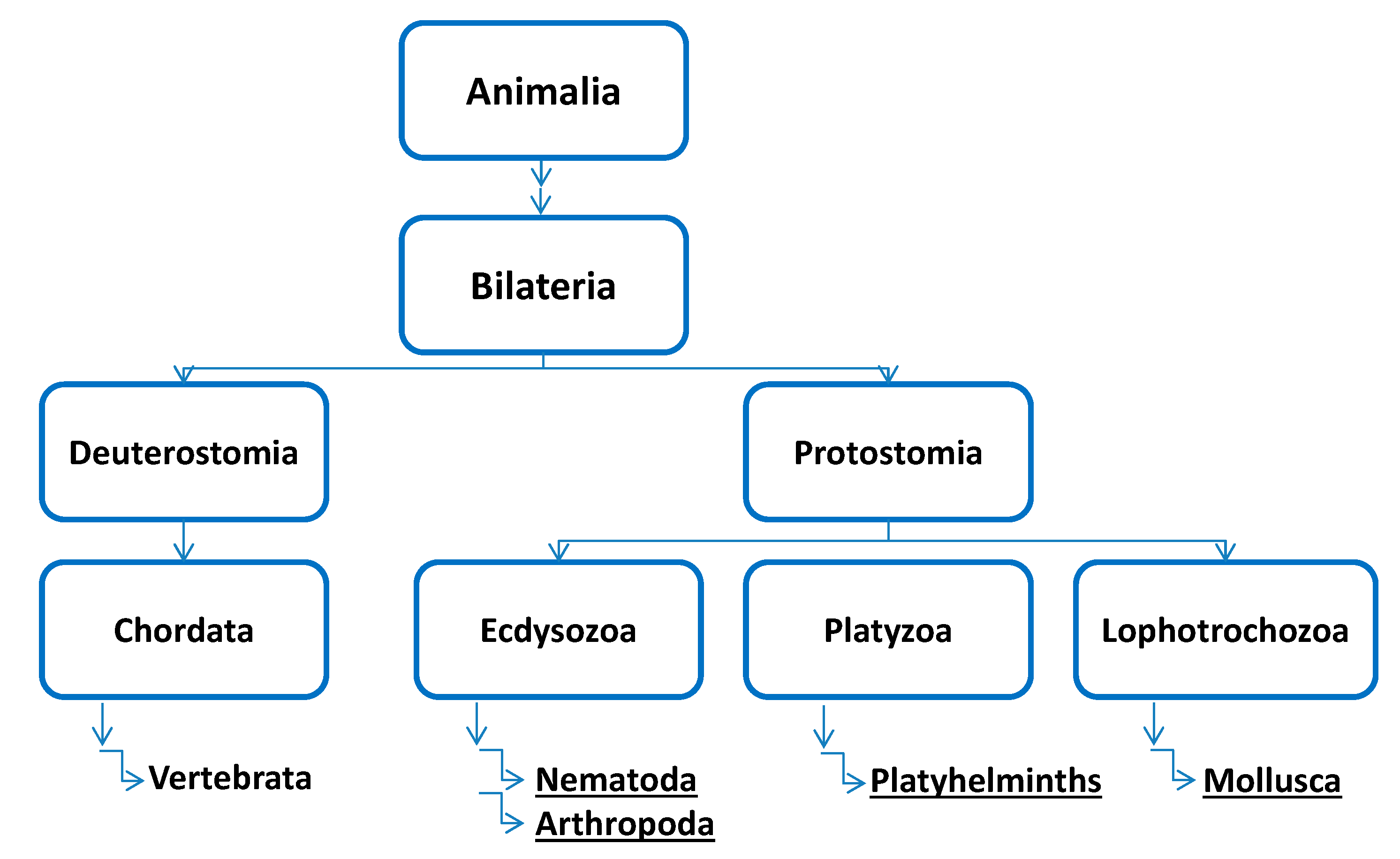
:1. Introduction

2. Nematoda

3. Arthropoda
3.1. Crustacea (Crustaceans)
3.2. Hexapoda (Insects)
3.2.1. Drosophila Melanogaster

3.2.2. Venoms
3.2.3. Insect Expression Systems

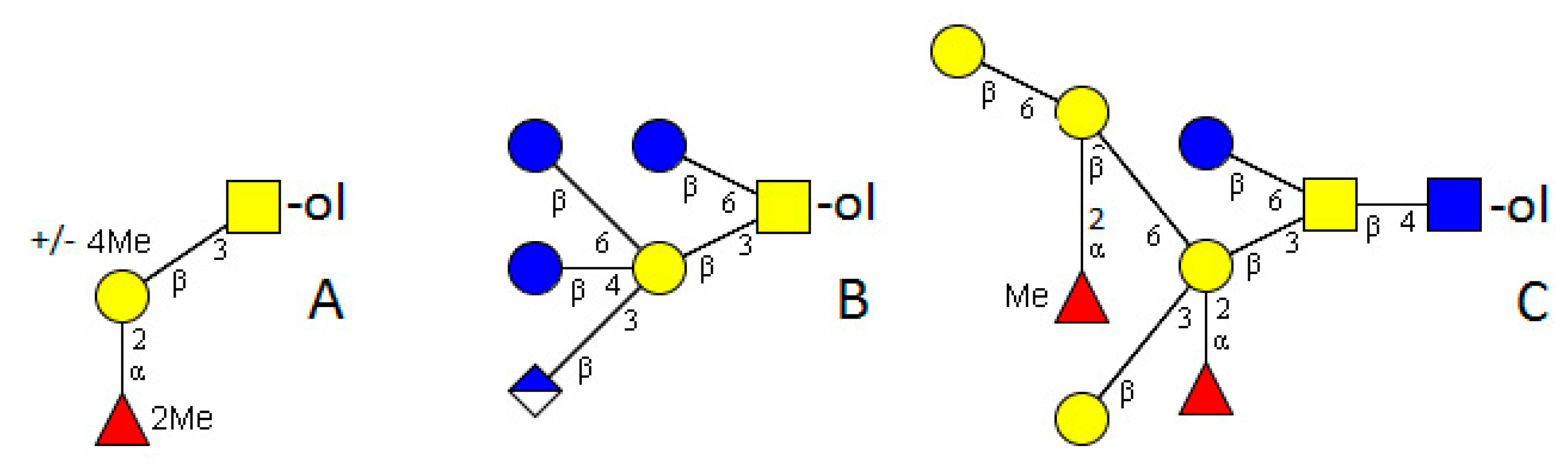
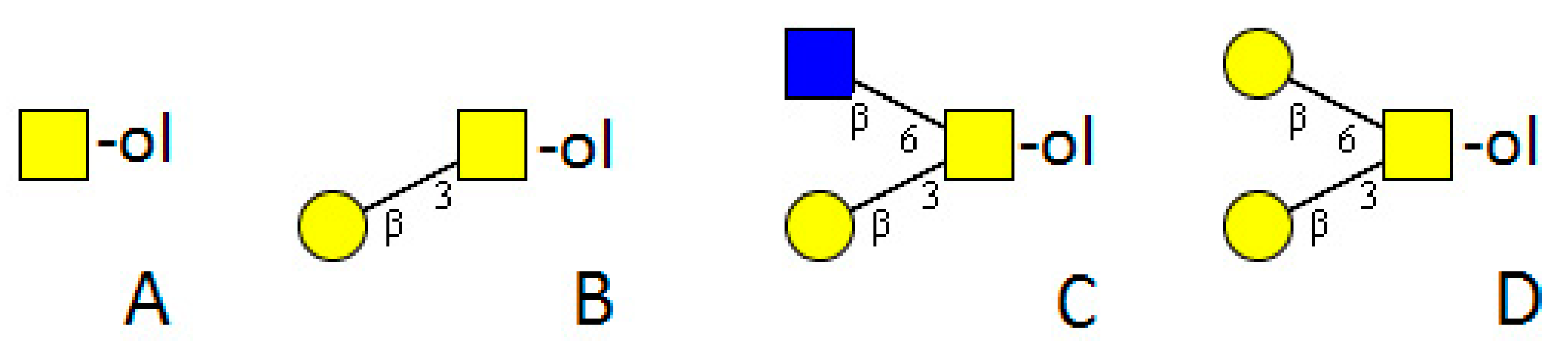
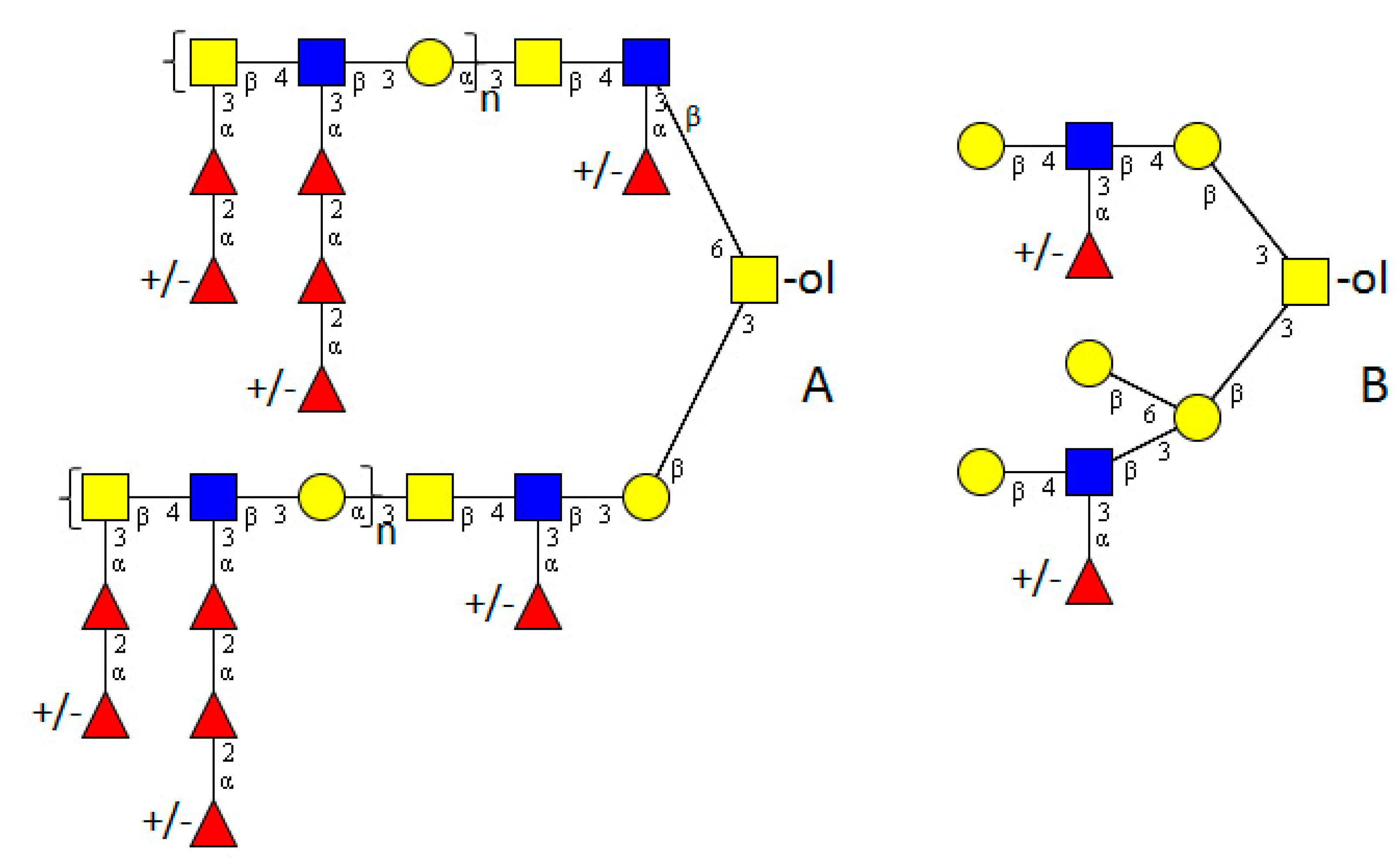
4. Platyhelminthes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
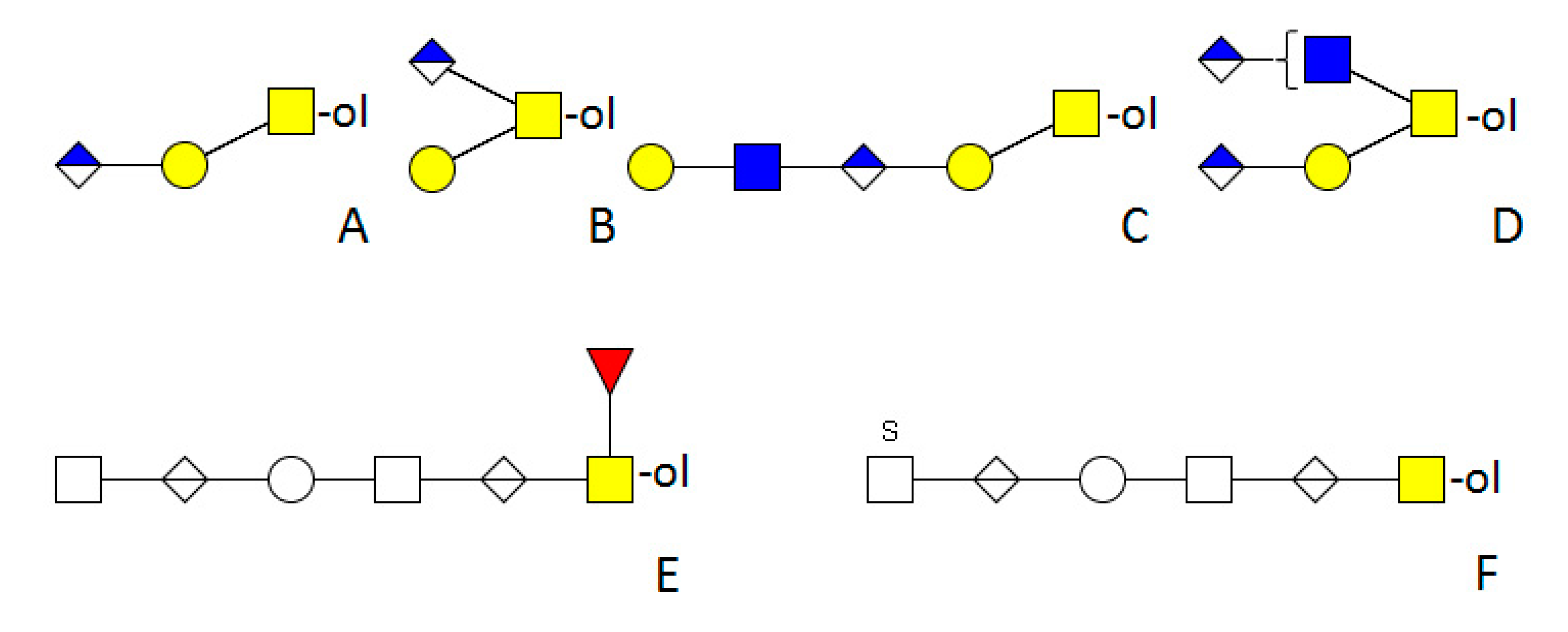
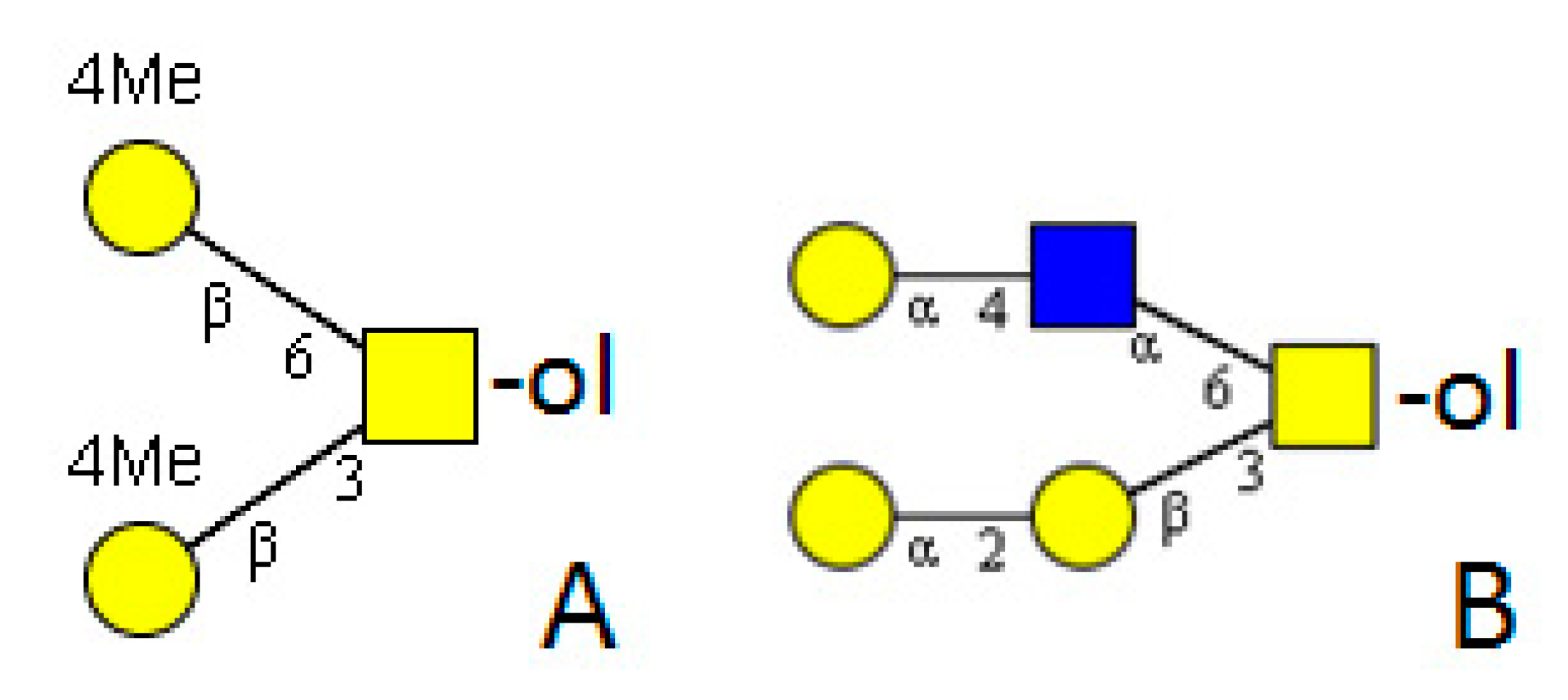
5. Mollusca
5.1. Cephalopoda
5.2. Gastropoda
5.2.1. Bivalves
5.2.2. Snails

6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Van den Steen, P.; Rudd, P.M.; Dwek, R.A.; Opdenakker, G. Concepts and principles of O-linked glycosylation. Crit. Rev. Biochem. Mol. Biol. 1998, 33, 151–208. [Google Scholar] [CrossRef] [PubMed]
- Proszynski, T.J.; Simons, K.; Bagnat, M. O-glycosylation as a sorting determinant for cell surface delivery in yeast. Mol. Biol. Cell 2004, 15, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Robledo, Y.; Marigómez, I.; Angulo, E.; Cajaraville, M.P. Glycosylation and sorting pathways of lysosomal enzymes in mussel digestive cells. Cell Tissue Res. 2006, 324, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Schjoldager, K.T.B.G.; Clausen, H. Site-specific protein O-glycosylation modulates proprotein processing—Decipering specific functions of the large polypeptide GalNAc-transferase gene family. Biochim. Biophys. Acta 2012, 1820, 2079–2094. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.J.; Tham, K.M.; Chia, J.; Wang, S.C.; Steentoft, C.; Clausen, H.; Bard-Chapeau, E.A.; Bard, F.A. Initiation of GalNAc-type O-glycosylation in the endoplasmic reticulum promotes cancer cell invasiveness. Proc. Natl. Acad. Sci. USA 2013, 110, E3152–E3161. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, P.; Dabelsteen, S.; Madsen, F.B.; Francavilla, C.; Kopp, K.L.; Steentoft, C.; Vakhrushev, S.Y.; Olsen, J.V.; Hansen, L.; Bennett, E.P.; et al. Immature truncated O-glycophenotype of cancer directly induces oncogenic features. Proc. Natl. Acad. Sci. USA 2014, 111, E4066–E4075. [Google Scholar] [CrossRef] [PubMed]
- Badirou, I.; Kurdi, M.; Legendre, P.; Rayes, J.; Bryckaert, M.; Casari, C.; Lenting, P.J.; Christophe, O.D.; Denis, C.V. In vivo analysis of the role of O-glycosylations of Von Willebrand factor. PLoS ONE 2012, 7, e37508. [Google Scholar] [CrossRef] [PubMed]
- Herzog, B.H.; Fu, J.; Xia, L. Mucin-type O-glycosylation is critical for vascular integrity. Glycobiology 2014, 24, 1237–1241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Syed, Z.A.; van Dijk Härd, I.; Lim, J.M.; Wells, L.; Ten Hagen, K.G. O-Glycosylation regulates polarized secretion by modulating Tango1 stability. Proc. Natl. Acad. Sci. USA 2014, 111, 7296–7301. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.T.; Ten Hagen, K.G. Mucin-type O-glycosylation during development. J. Biol. Chem. 2013, 288, 6921–6929. [Google Scholar] [CrossRef] [PubMed]
- Spiro, R.G. Protein glycosylation: nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002, 12, 43R–56R. [Google Scholar] [CrossRef] [PubMed]
- Bennett, E.P.; Mandel, U.; Clausen, H.; Gerken, T.A.; Fritz, T.A.; Tabak, L.A. Control of mucin-type O-glycosylation: A classification of the polypeptide GalNAc-transferase gene family. Glycobiology 2012, 22, 736–756. [Google Scholar] [CrossRef] [PubMed]
- Patsos, G.; Corfield, A. O-Glycosylation: Structural Diversity and Functions. In The Sugar Code; Gabius, H.J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 111–137. [Google Scholar]
- Bullen, J.W.; Balsbaugh, J.L.; Chanda, D.; Shabanowitz, J.; Hunt, D.F.; Neumann, D.; Hart, G.W. Cross-talk between two essential nutrient-sensitive enzymes. O-GlcNAc transferase (OGT) and AMP-activated protein kinase (AMPK). J. Biol. Chem. 2014, 289, 10592–10606. [Google Scholar] [PubMed]
- Live, D.; Wells, L.; Boons, G.J. Dissecting the molecular basis of the role of the O-mannosylation pathway in disease: α-Dystroglycan and forms of muscular dystrophy. ChemBioChem 2013, 14, 2392–2402. [Google Scholar] [CrossRef] [PubMed]
- Bektas, M.; Rubenstein, D.S. The role of intracellular protein O-glycosylation in cell adhesion and disease. J. Biomed. Res. 2011, 25, 227–236. [Google Scholar] [CrossRef]
- Ishio, A.; Sasamura, T.; Ayukawa, T.; Kuroda, J.; Ishikawa, H.O.; Aoyama, N.; Matsumoto, K.; Gushiken, T.; Okyjima, T.; Yamakawa, T.; Matsuno, K. O-Fucose monosaccharde of Drosophila Notch has a temperature-sensitive function and cooperates with O-glucose glycan in Notch transport and Notch signalling activation. J. Biol. Chem. 2015, 290, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Kantharia, J.; Sethi, M.K.; Bakker, H.; Haltiwanger, R.S. Site-specific O-glucosylation of the epidermal growth factor-like (EGF) repeats of Notch: efficiency of glycosylation is affected by proper folding and amino acid sequence of individual EGF repeats. J. Biol. Chem. 2012, 287, 33934–33944. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ng, D.T.W. O-mannosylation: The other glycan player of ER quality control. Semin. Cell Dev. Biol. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Lyalin, D.; Panin, V.M. Protein O-mannosylation in animal development and physiology: From human disorders to Drosophila phenotypes. Sem. Cell Dev. Biol. 2010, 21, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Brunner, A.; Kolarich, D.; Voglmeir, J.; Paschinger, K.; Wilson, I.B.H. Comparative characterisation of recombinant invertebrate and vertebrate peptide O-xylosyltransferases. Glycoconj. J. 2006, 23, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.H.; Maizels, R.M.; Page, A.P.; Taylor, G.W.; Rendell, N.B.; Dell, A. Characterization of nematode glycoproteins: the major O-glycans of Toxocara excretory-secretory antigens are O-methylated trisaccharides. Glycobiology 1991, 2, 163–171. [Google Scholar] [CrossRef]
- Schabussova, I.; Amer, H.; van Die, I.; Kosma, P.; Maizels, R.M. O-Methylated glycans from Toxocara are specific targets for antibody binding in human and animal infections. Int. J. Parasitol. 2007, 37, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Ellis, L.A.; McVay, C.S.; Probert, M.A.; Zhang, J.; Bundle, D.R.; Appleton, J.A. Terminal β-linked tyvelose creates unique epitopes in Trichinella spiralis glycan antigens. Glycobiology 1997, 7, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Romarís, F.; Escalante, M.; Lorenzo, S.; Bonay, P.; Gárate, T.; Leiro, J.; Ubeira, F.M. Monoclonal antibodies raised in Btk xid mice reveal new antigenic relationships and molecular interactions among gp53 and other Trichinella glycoproteins. Mol. Biochem. Parasitol. 2002, 125, 173–183. [Google Scholar] [CrossRef]
- Borloo, J.; de Graef, J.; Peelaers, I.; Nguyen, D.L.; Mitreva, M.; Devereese, B.; Hokke, C.H.; Vercruysse, J.; Claerebout, E.; Geldhof, P. In-depth proteomic and glycomic analysis of the adult-stage Cooperia oncophora excretome/secretome. J. Proteome Res. 2013, 12, 3900–3911. [Google Scholar] [CrossRef] [PubMed]
- Guérardel, Y.; Balanzino, L.; Maes, E.; Leroy, Y.; Coddeville, B.; Oriol, R.; Strecker, G. The nematode Caenorhabditis elegans synthesizes unusual O-linked glycans: Identification of glucose-substituted mucin-type O-glycans and short chondroitin-like oligosaccharides. Biochem. J. 2001, 357, 167–182. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.; Copeman, D.B.; Harnett, W. Do excretory-secretory products of Onchocerca gibsoni contain phosphorylcholine attached to O-type glycans? Int. J. Parasitol. 1996, 26, 1075–1080. [Google Scholar] [CrossRef]
- Hagen, F.K.; Nehrke, K. cDNA cloning and expression of a family of UDP-N-acetyl-d-galactosamine:polypeptide N-acetylgalactosaminyltransferase sequence homologs from Caenorhabditis elegans. J. Biol. Chem. 1998, 273, 8268–8277. [Google Scholar] [CrossRef] [PubMed]
- Hagen, F.; Layden, M.; Nehrke, K.; Gentile, K.; Berbach, K.; Tsao, C.C.; Forsythe, M. Mucin-type O-glycosylation in C. elegans is initiated by a family of glycosyltransferases. TIGG 2001, 13, 463–479. [Google Scholar] [CrossRef]
- Schwientek, T.; Bennett, E.P.; Flores, C.; Thacker, J.; Hollmann, M.; Reis, C.A.; Behrens, J.; Mandel, U.; Keck, B.; Schäfer, M.A.; et al. Functional conservation of subfamilies of putative UDP-N-acetylgalactosamine:polypeptide N-acetylgalactosaminyltransferases in Drosophila, Caenorhabditis elegans, and mammals: one subfamily composed of 1(2)35aa is essential in Drosophila. J. Biol. Chem. 2002, 277, 22623–22638. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Zheng, Q.; Cummings, R.D. Identification of core 1 O-glycan T-synthase from Caenorhabditis elegans. Glycobiology 2006, 16, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.; van Die, I.; Grundahl, K.M.; Kawar, Z.S.; Cummings, R.D. Molecular cloning and characterization of the Caenorhabditis elegans α1-3-fucosyltransferase family. Glycobiology 2007, 17, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Kawar, Z.S.; van Die, I.; Cummings, R.D. Molecular cloning and enzymatic characterization of a UDP-GalNAc:GlcNAcβR β1-4-N-acetygalactosaminyltransferase from Caenorhabditis elegans. J. Biol. Chem. 2002, 277, 34924–34932. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.Y.; Olson, S.K.; Esko, J.D.; Horvitz, H.R. Caenorhabditis elegans early embryogenesis and vulval morphogenesis require chondroitin biosynthesis. Nature 2003, 423, 439–443. [Google Scholar]
- Wang, H.; Spang, A.; Sullivan, M.A.; Hryhorenko, J.; Hagen, F.K. The terminal phase of cytokinesis in the Caenorhabditis elegans early embryo requires protein glycosylation. Mol. Biol. Cell 2005, 16, 4202–4213. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, S.; Kiruthika, J.; Ponniah, A.G.; Shekhar, M.S. Identification, cloning and expression of Catechol-O-methyltransferase (COMT) gene from shrimp, Penaeus monodon and its relevance to salinity stress. Fish Shellfish Immunol. 2012, 32, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Dolashka-Angelova, P.; Beltramini, M.; Dolashki, A.; Salvato, B.; Hristova, R.; Voelter, W. Carbohydrate composition of Carcinus aestuarii hemocyanin. Arch. Biochem. Biophys. 2001, 389, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Katoh, T.; Tiemeyer, M. The N’s and O’s of Drosophila glycoprotein glycobiology. Glycoconj. J. 2013, 30, 57–66. [Google Scholar] [CrossRef] [PubMed]
- North, S.J.; Koles, K.; Hembd, C.; Morris, H.R.; Dell, A.; Panin, V.M.; Haslam, S.M. Glycomic studies of Drosophila melanogaster embryos. Glycoconj. J. 2006, 23, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Tiemeyer, M. The glycomics of glycan glucuronylation in Drosophila melanogaster. Meth. Enzymol. 2010, 480, 297–321. [Google Scholar] [PubMed]
- Aoki, K.; Porterfield, M.; Lee, S.S.; Dong, B.; Nguyen, K.; McGlamry, K.H.; Tiemeyer, M. The diversity of O-linked glycans expressed during Drosophila melanogaster development reflects stage-and tissue-specific requirements for cell signalling. J. Biol. Chem. 2008, 283, 30385–30400. [Google Scholar] [CrossRef] [PubMed]
- Ten Hagen, K.G.; Tran, D.T. A UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase is essential for viability in Drosophila melanogaster. J. Biol. Chem. 2002, 277, 22616–22622. [Google Scholar] [CrossRef] [PubMed]
- Ten Hagen, K.G.; Tran, D.T.; Gerken, T.A.; Stein, D.S.; Zhang, Z. Functional characterization and expression analysis of members of the UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase family from Drosophila melanogaster. J. Biol. Chem. 2003, 278, 35039–35048. [Google Scholar] [CrossRef] [PubMed]
- Tian, E.; Ten Hagen, K.G. Expression of the UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase family is spatially and temporally regulated during Drosophila development. Glycobiology 2006, 16, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Tian, E.; Ten Hagen, K.G. O-linked glycan expression during Drosophila development. Glycobiology 2007, 17, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.T.; Zhang, L.; Zhang, Y.; Tian, E.; Earl, L.A.; Ten Hagen, K.G. Multiple members of the UDP-GalNAc: polypeptide N-acetylgalactosaminyltransferase family are essential for viability in Drosophila. J. Biol. Chem. 2011, 287, 5243–5252. [Google Scholar] [CrossRef] [PubMed]
- Gerken, T.A.; Ten Hagen, K.G.; Jamison, O. Conservation of peptide acceptor preferences between Drosophila and mammalian polypeptide-GalNAc transferase ortholog pairs. Glycobiology 2008, 18, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Müller, R.; Hülsmeier, A.J.; Altmann, F.; Ten Hagen, K.; Tiemeyer, M.; Hennet, T. Characterization of mucin-type core-1 beta1-3 galactosyltransferase homologous enzymes in Drosophila melanogaster. FEBS J. 2005, 272, 4295–4305. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.R.; Reddy, B.V.V.G.; Irvine, K.D. Requirement for a core 1 galactosyltransferase in the Drosophila nervous system. Dev. Dyn. 2008, 237, 3703–3714. [Google Scholar] [CrossRef] [PubMed]
- Rendić, D.; Sharrow, M.; Katoh, T.; Overcarsh, B.; Nguyen, K.; Kapurch, J.; Aoki, K.; Wilson, I.B.H.; Tiemeyer, M. Neural specific α3-fucosylation of N-linked glycans in the Drosophila embryo requires fucosyltransferase A and influcences developmental signalling associated with O-glycosylation. Glycobiology 2010, 20, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Sethi, M.K.; Buettner, F.F.R.; Krylov, V.B.; Takeuchi, H.; Nifantiev, N.E.; Haltiwanger, R.S.; Gerardy-Schahn, R.; Bakker, H. Identification of glycosyltransferase 8 family members as xylosyltransferases acting on O-glucosylated Notch epidermal growth factor repeats. J. Biol. Chem. 2010, 285, 1582–1586. [Google Scholar] [CrossRef] [PubMed]
- Yano, H.; Yamamoto-Hino, M.; Abe, M.; Kuwahara, R.; Haraguchi, S.; Kusaka, I.; Awano, W.; Konoshita-Toyoda, A.; Toyoda, H.; Goto, S. Distinct functional units of the Golgi complex in Drosophila cells. Proc. Natl. Acad. Sci. USA 2005, 102, 13467–13472. [Google Scholar] [CrossRef] [PubMed]
- Tian, E.; Ten Hagen, K.G. A UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase is required for epithelial tube formation. J. Biol. Chem. 2007, 282, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Y.; Ten Hagen, K.G. A mucin-type O-glycosyltransferase modulates cell adhesion during Drosophila development. J. Biol. Chem. 2008, 283, 34076–34086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ten Hagen, K.G. Dissecting the biological role of mucin-type O-glycosylation using RNA interference in Drosophila cell culture. J. Biol. Chem. 2010, 285, 34477–34484. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, W.; Focke, M.; Kolarich, D.; Wilson, I.B.H.; Altmann, F.; Wöhrl, S.; Götz, M.; Jarisch, R. Antibody binding to venom carbohydrates is a frequent cause for double positivity to honeybee and yellow jacket venom in patients with stinging-insect allergy. J. Allergy Clin. Immunol. 2001, 108, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Maes, E.; Garénaux, E.; Strecker, G.; Leroy, Y.; Wieruszeski, J.M.; Brassart, C.; Guérardel, Y. Major O-glycans from the nest of Vespula germanica contain phospho-ethanolamin. Carbohydr. Res. 2005, 340, 1852–1858. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Tetaert, D.; Julinat, S.; Gazon, M.; Cerutti, M.; Verbert, A.; Delannoy, P. O-Glycosylation potential of lepidopteran insect cell lines. Biochim. Biophys. Acta 1999, 1427, 49–61. [Google Scholar] [CrossRef]
- Lopez, M.; Gazon, M.; Juliant, S.; Plancke, Y.; Leroy, Y.; Strecker, G.; Cartron, M.P.; Bailly, P.; Cerutti, M.; Verbert, A.; Delannoy, P. Characterization of a UDP-Gal:Galβ1,3GalNAc α1,4-galactosyltransferase activity in a Mamestra brassicae cell line. J. Biol. Chem. 1998, 273, 33644–33651. [Google Scholar] [CrossRef] [PubMed]
- Gaunitz, S.; Jin, C.; Nilsson, A.; Liu, J.; Karlsson, N.G.; Holgersson, J. Mucin-type proteins produced in the Trichoplusia ni and Spodoptera frugiperda insect cell lines carry novel O-glycans with phosphocholine and sulfate substitutions. Glycobiology 2013, 23, 778–796. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Nakamura, M.; Scott, H.; Repnikova, E.; Carnahan, M.; Pandey, D.; Caster, C.; Khan, S.; Zimmermann, T.; Zoran, M.J.; Panin, V.M. The role of Drosophila cytidine monophosphate-sialic acid synthetase in the nervous system. J. Neurosci. 2013, 33, 12306–12315. [Google Scholar] [CrossRef] [PubMed]
- Koles, K.; Irvine, K.D.; Panin, V.M. Functional characterization of Drosophila sialyltransferase. J. Biol. Chem. 2004, 279, 4346–4357. [Google Scholar] [CrossRef] [PubMed]
- Freire, T.; Casaravilla, C.; Carmona, C.; Osinaga, E. Mucin-type O-glycosylation in Fasciola hepatica: Characterisation of carcinoma-associated Tn and sialyl-Tn antigens and evaluation of UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase activity. Int. J. Parasitol. 2003, 33, 47–56. [Google Scholar] [CrossRef]
- Errico, D.A.; Medeiros, A.; Míguez, M.; Casaravilla, C.; Malgor, R.; Carmona, C.; Nieto, A.; Osinaga, E. O-Glycosylation in Echinococcus granulosus: Identification and characterization of the carcinoma-associated Tn-antigen. Exp. Parasitol. 2001, 98, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Casaravilla, C.; Freire, T.; Malgor, R.; Medeiros, A.; Osinaga, E.; Carmona, C. Mucin-type O-glycosylation in helminth parasites from major taxonomic groups: Evidence for widespread distribution of the Tn antigen (GalNAc-Ser/Thr) and identification of UDP-GalNAc:poloypeptide N-acetylgalactosaminyltransferase activity. J. Parasitol. 2003, 89, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Freire, T.; Fernández, C.; Chalar, C.; Maizels, R.M.; Alzari, P.; Osinaga, E.; Robello, C. Characterization of a UDP-N-acetyl-d-galactosamine:polypeptide N-acetylgalactosaminyltransferase with an unusual lectin domain from the platyhelminth parasite Echinococcus granulosus. Biochem. J. 2004, 382, 501–510. [Google Scholar] [PubMed]
- Medeiros, A.; Chiribao, M.L.; Ubillos, L.; Festari, M.F.; Saldaña, J.; Tobello, C.; Domínguez, L.; Calvete, J.J.; Osinaga, E. Mucin-type O-glycosylation in Mesocestoides vogae (syn. Corti). Int. J. Parasitol. 2008, 38, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Ubillos, L.; Medeiros, A.; Cancela, M.; Casaravilla, C.; Saldaña, J.; Domínguez, L.; Carmona, C.; le Pendu, J.; Osinaga, E. Characterization of the carcinoma-associated Tk antigen in helminth parasites. Exp. Parasitol. 2007, 116, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.H.; Sarda, S.; Xu, X.; Caufield, J.P.; Mcneil, M.R.; Homans, S.W.; Morris, H.R.; Dell, A. A unique multifucosylated -3GalNAcβ1-4GlcNAcβ1-3Galα1-motif constitutes the repeating unit of the complex O-glycans derived from the cercarial glycocalyx of Schistosoma mansoni. J. Biol. Chem. 1995, 270, 17114–17123. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.H.; Chatterjee, D.; Caulfield, J.P.; Morris, H.R.; Dell, A. Structural mapping of the glycans from the egg glycoproteins of Schistosoma mansoni and Schistosoma japonicum: identification of novel core structures and terminal sequences. Glycobiology 1997, 7, 663–677. [Google Scholar] [CrossRef] [PubMed]
- Robijn, M.L.M.; Koeleman, C.A.M.; Wuhrer, M.; Royle, L.; Geyer, R.; Dwek, R.A.; Rudd, P.M.; Deelder, A.M.; Hokke, C.H. Targeted identification of a unique glycan epitope of Schistosoma mansoni egg antigens using a diagnostic antibody. Mol. Biochem. Parasitol. 2007, 151, 148–161. [Google Scholar] [CrossRef] [PubMed]
- De Walick, S.; Bexkens, M.L.; van Balkom, B.W.M.; Wu, Y.P.; Smit, C.H.; Hokke, C.H.; de Groot, P.G.; Heck, A.J.R.; Tielens, A.G.M.; van Hellemond, J.J. The proteome of the insoluble Schistosoma mansoni eggshell skeleton. J. Parasitol. 2013, 41, 523–532. [Google Scholar]
- Huang, H.H.; Tsai, P.L.; Khoo, K.H. Selective expression of different fucosylated epitopes on two distinct sets of Schistosoma mansoni cercarial O-glycans: Identification of a novel core type and Lewis X structure. Glycobiology 2001, 11, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Peterson, N.A.; Hokke, C.H.; Deelder, A.M.; Yoshino, T.P. Glycotope analysis in miracidia and primary sporocysts of Schistosoma mansoni: Differential expression during the miracidium-to-sporocyst transformation. Int. J. Parasitol. 2009, 39, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Van Roon, A.-M.M.; Aguilera, B.; Cuenca, F.; van Remoortere, A.; van der Marel, G.A.; Deelder, A.M.; Overkleeft, H.S.; Hokke, C.H. Synthesis and antibody-binding studies of a series of parasite fuco-oligosaccharides. Bioorg. Med. Chem. 2005, 13, 3553–3564. [Google Scholar] [CrossRef] [PubMed]
- Marques, E.T.A., Jr.; Ichikawa, Y.; Strand, M.; August, J.T.; Hart, G.W.; Schnaar, R.L. Fucosyltranferases in Schistosoma mansoni development. Glycobiology 2001, 11, 249–259. [Google Scholar] [CrossRef]
- Hokke, C.H.; Neeleman, A.P.; Koelman, C.A.M.; van den Eijneden, D.H. Identification of an α3-fucosyltransferase and a novel α2-fucosyltransferase in cercariae of the schistosome Trichobilharzia ocellata: Biosynthesis of the Fucα1-2Fucα1-3[Gal(NAc)β1-4]GlcNAc sequence. Glycobiology 1998, 8, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Hokke, C.H.; Yazdanbakhsh, M. Schistosome glycans and innate immunity. Parasite Immunol. 2005, 27, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Hokke, C.H.; Fitzpatrick, J.M.; Hoffmann, K.F. Integrating transcriptome, proteome and glycome analyses of Schistosoma biology. Trends Parasitol. 2007, 23, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.G.; Harn, D.A., Jr. Immune biasing by helminth glycans. Cell. Microbiol. 2004, 6, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Osinaga, E. Expression of cancer-associated simple mucin-type O-glycosylated antigens in parasites. IUBMB Life 2007, 59, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Bui, C.T.; Shollenberger, L.M.; Paterson, Y.; Harn, D.A. Schistosoma mansoni soluble egg antigens enhance T cell responses to a newly identified HIV-1 Gag H-2b epitope. Clin. Vaccine Immunol. 2015, 22, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Bui, C.T.; Shollenberger, L.M.; Paterson, Y.; Harn, D.A. Schistosoma mansoni soluble egg antigens enhance Listeria vector HIV-1 vaccine induction of cytotoxic T cells. Clin. Vaccine Immunol. 2014, 21, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Van Remoortere, A.; Hokke, C.H.; van Dam, G.J.; van Die, I.; Deelder, A.M.; van den Eijnden, D.H. Various stages of Schistosoma express Lewisx, LacdiNAc, GalNAcβ1-4(Fucα1-3)GlcNAc and GalNAcβ1-4(Fucα1-2Fucα1-3)GlcNAc carbohydrate epitopes: detection with monoclonal antibodies that are charcterized by enzymatically synthesized neoglycoproteins. Glycobiology 2000, 10, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Wammes, L.J.; Mpairwe, H.; Elliott, A.M.; Yazdanbakhsh, M. Helminth therapy or elimination: epidemiological, immunological, and clinical considerations. Lancet Infect. Dis. 2014, 14, 1150–1162. [Google Scholar] [CrossRef]
- Nakagawa, M.; Miyamoto, T.; Kusakabe, R.; Takasaki, S.; Takao, T.; Shichida, Y.; Tsuda, M. O-Glycosylation of G-protein-coupled receptor, octopus rhodopsin. Direct analysis by FAB mass spectrometry. FEBS Lett. 2001, 496, 19–24. [Google Scholar] [CrossRef]
- Kurz, S.; Jin, C.; Hykollari, A.; Gregorich, D.; Giomarelli, B.; Vasta, G.R.; Wilson, I.B.H.; Paschinger, K. Hemocytes and plasma of the Eastern oyster (Crassostrea virginica) display a diverse repertoire of sulfated and blood group A-modified N-glycans. J. Biol. Chem. 2013, 288, 24410–24428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Gerwig, G.J.; Hocking, H.G.; Stöcklin, R.; Kamerling, J.P.; Boelens, R. Glycosylation of conotoxins. Mar. Drugs 2013, 11, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Hocking, H.G.; Gerwig, G.J.; Dutertre, S.; Violette, A.; Favreau, P.; Stöcklin, R.; Kamerling, J.P.; Boelens, R. Structure of the O-glycosylated conopeptide CcTx from Conus consors venom. Chem. Eur. J. 2013, 19, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Geyer, H.; Wuhrer, M.; Kurokawa, T.; Geyer, R. Charcterization of keyhole limpet hemocyanin (KLH) glycans sharing a carbohydrate epitope with Schistosoma mansoni glycoconjugates. Micron 2004, 35, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Lehr, T.; Frank, S.; Natsuka, S.; Geyer, H.; Beuerlein, K.; Doenhoff, M.J.; Hase, S.; Geyer, R. N-Glycosylation patterns of hemolymph glycoproteins from Biomphalaria glabrata strains expressing different susceptibility to Schistosma mansoni infection. Exp. Parasitol. 2010, 126, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.P.; Wu, X.J.; Gonzalez, L.A.; Hokke, C.H. Circulating Biomphalaria glabrata hemocyte subpopulations possess shared schistosome glycans and receptors capable of binding larval glycoconjugates. Exp. Parasitol. 2013, 133, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Stoeva, S.; Schütz, J.; Gebauer, W.; Hundsdörfer, T.; Manz, C.; Markl, J.; Voelter, W. Primary structure and unusual arbohydrate moiety of functional unit 2-c of keyhole limpet hemocyanin (KLH). Biochim. Biophys. Acta 1999, 1435, 94–109. [Google Scholar] [CrossRef]
- Petraccioli, A.; Maio, N.; Guarino, F.M.; Scillitani, G. Seasonal variation in glycoconjugates of the pedal glandular system of the rayed Mediterranean limpet, Patella caerulea (Gastropoda: Patellidae). Zoology 2013, 116, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Bürgmayr, S.; Grabher-Meier, H.; Staudacher, E. Sialic acids in gastropods. FEBS Lett. 2001, 508, 95–98. [Google Scholar] [CrossRef]
- Stepan, H.; Bleckmann, C.; Geyer, H.; Geyer, R.; Staudacher, E. Determination of 3-O- and 4-O-methylated monosaccharide constituents in snail glycans. Carbohydr. Res. 2010, 345, 1504–1507. [Google Scholar] [CrossRef] [PubMed]
- Stepan, H.; Staudacher, E. Optimization of monosaccharide determination using anthranilic acid and 1-phenyl-3-methyl-5-pyrazolone for gastropod analysis. Anal. Biochem. 2011, 418, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Stepan, H.; Pabst, M.; Altmann, F.; Geyer, H.; Geyer, R.; Staudacher, E. O-Glycosylation of snails. Glycoconj. J. 2012, 29, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Taus, C.; Lucini, C.; Sato, T.; Furukawa, K.; Grabherr, R.; Staudacher, E. Expression and characterization of the first snail-derived UDP-N-acetyl-α-d-galactosamine:polypeptide N-acetylgalactosaminyltransferase. Glycoconj. J. 2013, 30, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Taus, C.; Windwarder, M.; Altmann, F.; Grabherr, R.; Staudacher, E. UDP-N-acetyl-α-d-galactosamine:polypeptide N-acetylgalactosaminyl-transferase from the snail Biomphalaria glabrata—Substrate specificity and preference of glycosylation sites. Glycoconj. J. 2014, 31, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Van Die, I.; Cummings, R.D.; van Tetering, A.; Hokke, C.H.; Koeleman, C.A.M.; Van den Eijnden, D.H. Identification of a novel UDP-Glc:GlcNAc β1-4-glucosyltransferase in Lymnaea stagnalis that may be involved in the synthesis of complex-type oligosaccharide chains. Glycobiology 2000, 10, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, E. Methylation—An uncommon modification of glycans. Biol. Chem. 2012, 393, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Wohlschlager, T.; Butschi, A.; Grassi, P.; Sutov, G.; Gauss, R.; Hauck, D.; Schmieder, S.S.; Knobel, M.; Titz, A.; Dell, A.; et al. Methylated glycans as conserved targets of animal and fungal innate defense. Proc. Natl. Acad. Sci. USA 2014, 111, E2787–E2796. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staudacher, E. Mucin-Type O-Glycosylation in Invertebrates. Molecules 2015, 20, 10622-10640. https://doi.org/10.3390/molecules200610622
Staudacher E. Mucin-Type O-Glycosylation in Invertebrates. Molecules. 2015; 20(6):10622-10640. https://doi.org/10.3390/molecules200610622
Chicago/Turabian StyleStaudacher, Erika. 2015. "Mucin-Type O-Glycosylation in Invertebrates" Molecules 20, no. 6: 10622-10640. https://doi.org/10.3390/molecules200610622
APA StyleStaudacher, E. (2015). Mucin-Type O-Glycosylation in Invertebrates. Molecules, 20(6), 10622-10640. https://doi.org/10.3390/molecules200610622