
Development and Characterization of Novel EST-SSRs from Larix gmelinii and Their Cross-Species Transferability

Abstract
:1. Introduction
2. Results and Discussion
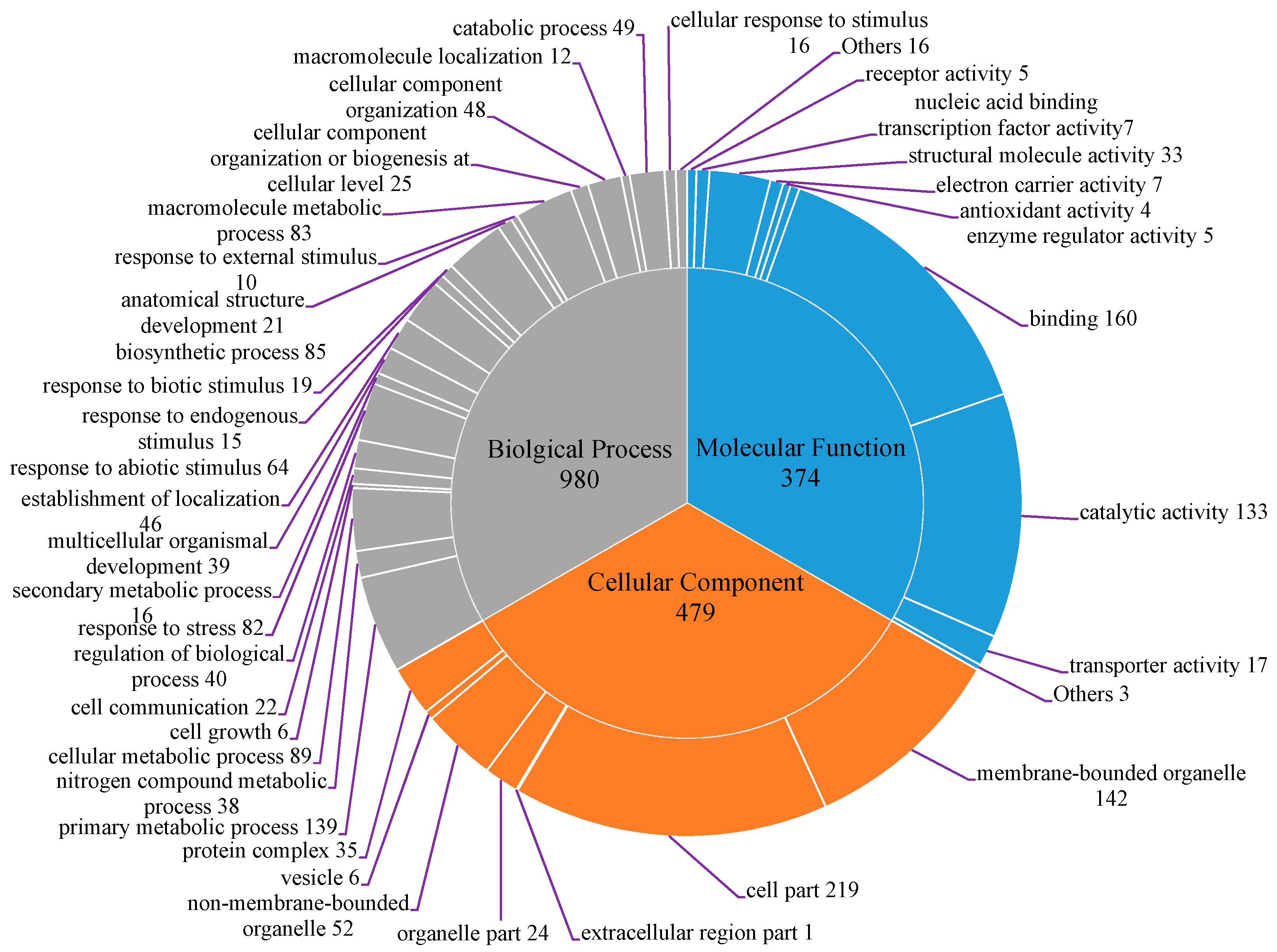
2.1. EST Data Assemble and Annotation

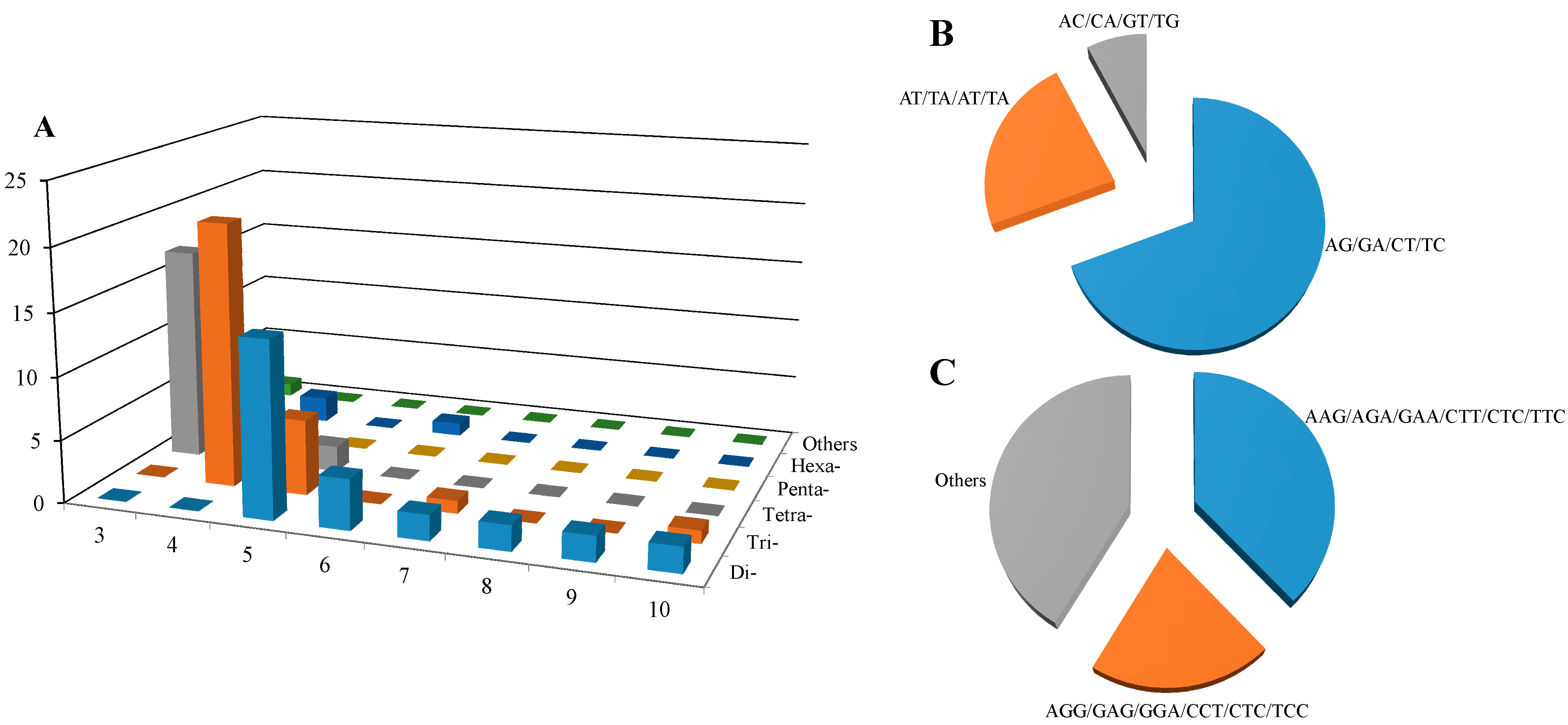
2.2. SSR Identification, Motif Frequency Distribution and ORF Prediction

2.3. EST-SSR Marker Development, Validation and Transferability Analysis in L. gmelinii and Its Related Larch Species

{kind=link}
{kind=link}
| Locus | Primers Sequence (5′-3′) | Repeats Motif | Size Range | Ta (°C) | SSR Locations | BLAST Top Hit Accession No. | Description of Putative Function | E-Value |
|---|---|---|---|---|---|---|---|---|
| Lg01 | F: CAGTGGTGTCCGTGGTGTA R:GACCTCCTCCACACCTAAT | (AGC)4 | 141–160 | 51.3 | ORF | XP_006375910.1 | hypothetical protein POPTR_0013s05890g (Populus trichocarpa) | 2.00 × 10−20 |
| Lg02 | F: CTCTGTGACCAAGAAACCAA R:CATGAAGACGAAGAATGCACT | (AGG)4 | 120–140 | 51.8 | ORF | XP_002306980.2 | hypothetical protein POPTR_0005s27390g (P. trichocarpa) | 3.00 × 10−38 |
| Lg06 | F: CAAGGATGGAGCAGACGAT R:AGCCTCGCACTTTGACAGA | (AGA)5 | 135–150 | 50.4 | ORF | None | None | None |
| Lg14 | F: GGGGATTGCAGAGTAGAAA R:AAACAGCCATCGAAATGAG | (TC)6 | 140–150 | 49.5 | 5′UTR | XP_002319953.1 | Xyloglucan endotransglucosylase/hydrolase protein 9 precursor (P. trichocarpa) | 1.00 × 10−164 |
| Lg25 | F: GTGAGAGGTCAAACCCCAA R: AGAAGAGTCTGGTCCACGCT | (AAG)4 | 105–125 | 53.8 | ORF | XP_003608708.1 | 40S ribosomal protein S30 (Medicago truncatula) | 2.00 × 10−24 |
| Lg32 | F: CTCTGTCGCACCAGCATTG R:TTGTCTTCCGGTATTCACA | (AT)6 | 105–115 | 47.5 | ORF | XP_002307364.1 | 2-dehydro-3-deoxyphosphoheptonate aldolase family protein (P. trichocarpa) | 4.00 × 10−7 |
| Lg36 | F: TGCCCATCCTCTTTGTTTA R: AGCACCTGATTCCACATTCT | (GA)5 | 175–190 | 50.6 | ORF | None | None | None |
| Lg37 | F: ACAATGGCTTCCTTCAACA R: TATGAGGTGGTTAGGGAGA | (CT)6 | 160–170 | 47.1 | ORF | XP_002299125.2 | hypothetical protein POPTR_0001s04570g (P. trichocarpa) | 3.00 × 10−46 |
| Lg41 | F: ACTTCCACTAAGGTTGACA R:ATCCACTGCCTTCTGGTCAT | (AGA)4 | 147–180 | 49.4 | ORF | XP_002313280.1 | 60S ribosomal protein L6 (P. trichocarpa) | 2.00 × 10−49 |
| Locus | L. gmelinii Jiamusi Population | L. gmelinii Hulunbeier Population | L. kaempferi | L. principis-rupprechtii | L. olgensis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (N = 15) | (N = 15) | (N = 8) | (N = 10) | (N = 9) | |||||||||||
| Na | Ho/He | FIS | Na | Ho/He | FIS | Na | Ho/He | FIS | Na | HoHe | FIS | Na | Ho/He | FIS | |
| Lg01 | 2 | 0.467/0.704 | 0.338 | 2 | 0.200/0.278 | 0.280 | 3 | 0.000/0.406 | 1.000 ** | 2 | 0.100/0.375 | 0.733 * | 2 | 0.111/0.500 | 0.778 * |
| Lg02 | 2 | 0.600/0.464 | −0.292 | 4 | 0.733/0.560 | −0.310 | 2 | 0.500/0.469 | −0.067 | 3 | 0.800/0.535 | −0.495 | 2 | 0.667/0.494 | −0.350 |
| Lg06 | 2 | 0.267/0.498 | 0.464 | 2 | 0.267/0.498 | 0.464 | 2 | 0.000/0.219 | 1.000 ** | 2 | 0.000/0.180 | 1.000 ** | 2 | 0.000/0.346 | 1.000 ** |
| Lg14 | 2 | 0.200/0.358 | 0.441 | 2 | 0.267/0.480 | 0.444 | 1 | 0.000/0.000 | -- | 2 | 0.100/0.375 | 0.733 * | 2 | 0.111/0.278 | 0.600 |
| Lg25 | 3 | 0.400/0.371 | −0.078 | 3 | 0.733/0.598 | −0.227 | 2 | 0.250/0.375 | 0.333 | 3 | 0.400/0.465 | 0.140 | 2 | 0.111/0.278 | 0.600 |
| Lg32 | 2 | 0.267/0.444 | 0.400 | 3 | 0.467/0.558 | 0.163 | 1 | 0.000/0.000 | -- | 2 | 0.000/0.180 | 1.000 ** | 2 | 0.000/0.198 | 1.000 ** |
| Lg36 | 2 | 0.267/0.498 | 0.464 | 2 | 0.267/0.320 | 0.167 | 1 | 0.000/0.000 | -- | 2 | 0.100/0.255 | 0.608 | 2 | 0.333/0.401 | 0.169 |
| Lg37 | 2 | 0.200/0.358 | 0.441 | 3 | 0.400/0.611 | 0.345 | 1 | 0.000/0.000 | -- | 2 | 0.100/0.255 | 0.608 | 1 | 0.000/0.000 | -- |
| Lg41 | 3 | 0.467/0.584 | −0.167 * | 3 | 0.600/0.620 | −0.150 * | 3 | 0.625/0.617 | −0.013 | 2 | 0.500/0.495 | −0.010 | 2 | 0.333/0.500 | 0.333 * |
3. Experimental Section
3.1. Plant Materials
3.2. EST Data Assemble and Annotation
3.3. SSR Identification and Primer Designs
3.4. PCR Amplification, Cross-Species Transferability and Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oreshkova, N.V.; Belokon, M.M.; Jamiyansuren, S. Genetic diversity, population structure, and differentiation of Siberian larch, Gmelin larch, and Cajander larch on SSR-marker data. Russ. J. Genet. 2013, 49, 178–186. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, H.G.; Li, X.F. Analysis of genetic diversity in Larix gmelinii (Pinaceae) with RAPD and ISSR markers. Genet. Mol. Res. 2013, 12, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, H.G.; Du, J.; Li, X.F. RAPD and SSR analysis of genetic diversity of natural Larix Gmelinii populations. Biotechnol. Biotechnol. Equip. 2013, 27, 3959–3965. [Google Scholar] [CrossRef]
- Di, X.Y.; Li, X.N.; Wang, Q.X.; Wang, M.B. Genetic diversity of natural populations of Larix principis-rupprechtii in Shanxi Province, China. Biochem. Syst. Ecol. 2014, 54, 71–77. [Google Scholar] [CrossRef]
- Kozyrenko, M.M.; Artyukova, E.V.; Reunova, G.D.; Levina, E.A.; Zhuravlev, Y.N. Genetic diversity and relationships among Siberian and Far Eastern larches inferred from RAPD analysis. Russ. J. Genet. 2004, 40, 401–409. [Google Scholar] [CrossRef]
- Khasa, D.P.; Jaramillo-Correa, J.P.; Jaquish, B.; Bousquet, J. Contrasting microsatellite variation between subalpine and western larch, two closely related species with different distribution patterns. Mol. Ecol. 2006, 15, 3907–3918. [Google Scholar] [CrossRef] [PubMed]
- Gros-Louis, M.C.; Bousquet, J.; Pâques, L.E.; Isabel, N. Species-diagnostic markers in Larix spp. based on RAPDs and nuclear, cpDNA, and mtDNA gene sequences, and their phylogenetic implications. Tree Genet. Genomes 2005, 1, 50–63. [Google Scholar] [CrossRef]
- Nadeem, S.; Jaquish, B.; Newton, C.; Khasa, P.D. Use of diagnostic SSR markers for identification of Larix lyallii and L. occidentalis (Pinaceae). Edinb. J. Bot. 2003, 60, 49–56. [Google Scholar] [CrossRef]
- Peng, M.; Qu, L.N.; Wang, Q.Y. Seed-specific identification of Larix gmelinii, Larix olgensis, and Larix principis-rupprechtii using sequence-characterised amplified region markers. Biochem. Syst. Ecol. 2014, 55, 231–235. [Google Scholar] [CrossRef]
- Funda, T.; Chen, C.C.; Liewlaksaneeyanawin, C.; Kenawy, A.A.; El-Kassaby, Y.A. Pedigree and mating system analyses in a western larch (Larix occidentalis Nutt.) experimental population. Ann. For. Sci. 2008, 65, 705. [Google Scholar] [CrossRef]
- Guan, C.Y. Construction of Genetic Linkage Map of RAPD, SSR and QTL Analysis in Larch (Larix Kaempferi × Larix gmelini). Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2010. [Google Scholar]
- Krutovsky, K.V.; Tretyakova, I.N.; Oreshkova, N.V.; Pak, M.E.; Kvitko, O.V.; Vaganov, E.A. Somaclonal variation of haploid in vitro tissue culture obtained from Siberian larch (Larix sibirica Ledeb.) megagametophytes for whole genome de novo sequencing. In Vitro Cell. Dev. Biol. Plant 2014, 50, 655–664. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Isoda, K.; Eatanabe, A. Isolation and characterization of microsatellite loci from Larix kaempferi. Mol. Ecol. Notes 2006, 6, 664–666. [Google Scholar] [CrossRef]
- Chen, C.; Liewlaksaneeyanawin, C.; Funda, T.; Kenawy, A.; Newton, C.H.; El-kassaby, Y.A. Development and characterization of microsatellite loci in western larch (Larix occidentalis Nutt.). Mol. Ecol. Resour. 2009, 9, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Sun, X.M.; Zhang, S.G. Short note: Development of six EST-SSR markers in Larch. Silvae Genet. 2011, 60, 161–163. [Google Scholar]
- Wagner, S.; Gerber, S.; Petit, R.J. Two highly informative dinucleotide SSR multiplexes for the conifer Larix decidua (European larch). Mol. Ecol. Resour. 2012, 12, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Jing, H. Construction and Analysis of an Inducible Insect Resistant cDNA Library and Its Expressed Sequence Tags from Jasmonic Acid-Treated Larix gmelinii Needles. Master Thesis, Northeast Forestry University, Harbin, China, 2010. [Google Scholar]
- Men, L.N.; Yan, S.C.; Liu, G.J. De novo characterization of Larix gmelinii (Rupr.) Rupr. transcriptome and analysis of its gene expression induced by jasmonates. BMC Genomics 2013, 14, 548. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Sun, X.M.; Zhang, S.G.; Xie, Y.H.; Han, H. Development of EST-SSR markers and genetic diversity analysis of second cycle elite population in Larix Kaempferi. Sci. Silvae Sin. 2011, 47, 52–58. [Google Scholar]
- Wei, X.; Wang, L.H.; Zhang, Y.X.; Qi, X.Q.; Wang, X.L.; Ding, X.; Zhang, J.; Zhang, X.R. Development of simple sequence repeat (SSR) markers of sesame (Sesamum indicum) from a genome survey. Molecules 2014, 19, 5150–5162. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.P.; Liu, P.; Luo, D.; Liu, W.X.; Wang, Y.R. Exploiting Illumina sequencing for the development of 95 novel polymorphic EST-SSR markers in common vetch (Vicia sativa subsp. sativa). Molecules 2014, 19, 5777–5578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, W.G.; Song, X.M.; Lin, Q.H.; Gui, J.F.; Mei, J. Characterization and development of EST-SSR markers derived from transcriptome of Yellow Catfish. Molecules 2014, 19, 16402–16415. [Google Scholar] [CrossRef] [PubMed]
- Acuña, C.V.; Fernandez, P.; Villalba, P.V.; García, M.N.; Hopp, H.E.; Poltri, S.N.M. Discovery, validation, and in silico functional characterization of EST-SSR markers in Eucalyptus globulus. Tree Genet. Genomes 2012, 8, 289–301. [Google Scholar] [CrossRef]
- Li, W.F.; Han, S.Y.; Qi, L.W.; Zhang, S.G. Transcriptome resources and genome-wide marker development for Japanese larch Larix kaempferi. Front. Agric. Sci. Eng. 2014, 1, 77–84. [Google Scholar]
- Sample Availability: All samples are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Sun, Z.; Zhou, D.; Xiong, M.; Wang, X.; Yang, J.; Wei, Z. Development and Characterization of Novel EST-SSRs from Larix gmelinii and Their Cross-Species Transferability. Molecules 2015, 20, 12469-12480. https://doi.org/10.3390/molecules200712469
Zhang G, Sun Z, Zhou D, Xiong M, Wang X, Yang J, Wei Z. Development and Characterization of Novel EST-SSRs from Larix gmelinii and Their Cross-Species Transferability. Molecules. 2015; 20(7):12469-12480. https://doi.org/10.3390/molecules200712469
Chicago/Turabian StyleZhang, Guojun, Zhenzhen Sun, Di Zhou, Min Xiong, Xian Wang, Junming Yang, and Zunzheng Wei. 2015. "Development and Characterization of Novel EST-SSRs from Larix gmelinii and Their Cross-Species Transferability" Molecules 20, no. 7: 12469-12480. https://doi.org/10.3390/molecules200712469
APA StyleZhang, G., Sun, Z., Zhou, D., Xiong, M., Wang, X., Yang, J., & Wei, Z. (2015). Development and Characterization of Novel EST-SSRs from Larix gmelinii and Their Cross-Species Transferability. Molecules, 20(7), 12469-12480. https://doi.org/10.3390/molecules200712469