
Biochanin A Ameliorates Arsenic-Induced Hepato- and Hematotoxicity in Rats

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. General Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight (g) | Food Intake (g/rat/day ) | Water Intake (mL/rat/day) | Liver Weight (g) | Liver-Body Weight Ratio (%) | |
|---|---|---|---|---|---|---|
| Initial | Final | |||||
| Normal control | 203.33 ± 17.42 | 312.33 ± 27.41 | 14.66 ± 1.21 | 20.50 ± 1.84 | 21.16 ± 1.54 | 6.77 ± 0.12 |
| Normal + Vehicle | 202.00 ± 19.26 | 314.25 ± 28.46 | 14.83 ± 1.64 | 20.24 ± 1.92 | 21.06 ± 1.98 | 6.89 ± 0.06 |
| Normal + BCA (40 mg/kg) | 201.66 ± 18.22 | 315.24 ± 28.64 | 13.83 ± 0.98 | 20.74 ± 1.84 | 21.16 ± 1.12 | 6.74 ± 0.06 |
| Arsenic (10 mg/kg) | 205.33 ± 17.24 | 309.65 ± 26.48 | 13.16 ± 0.96 | 19.33 ± 1.74 | 21.86 ± 1.28 | 7.17 ± 0.48 |
| Arsenic +Selenium (3 mg/kg) | 203.45 ± 18.24 | 313.52 ± 28.46 | 13.00 ± 1.09 | 20.64 ± 1.84 | 22.13 ± 1.41 | 7.11 ± 0.14 |
| Normal + BCA (10 mg/kg) | 205.66 ± 18.42 | 309.84 ± 28.42 | 13.16 ± 1.21 | 20.68 ± 1.42 | 22.27 ± 1.58 | 7.22 ± 0.10 |
| Normal + BCA (20 mg/kg) | 208.15 ± 17.42 | 310.34 ± 24.52 | 13.50 ± 1.42 | 19.48 ± 1.75 | 22.12 ± 1.72 | 7.16 ± 0.12 |
| Normal + BCA (40 mg/kg) | 204.64 ± 15.26 | 310.54 ± 26.64 | 13.83 ± 1.16 | 19.95 ± 1.64 | 21.64 ± 1.84 | 6.99 ± 0.18 |
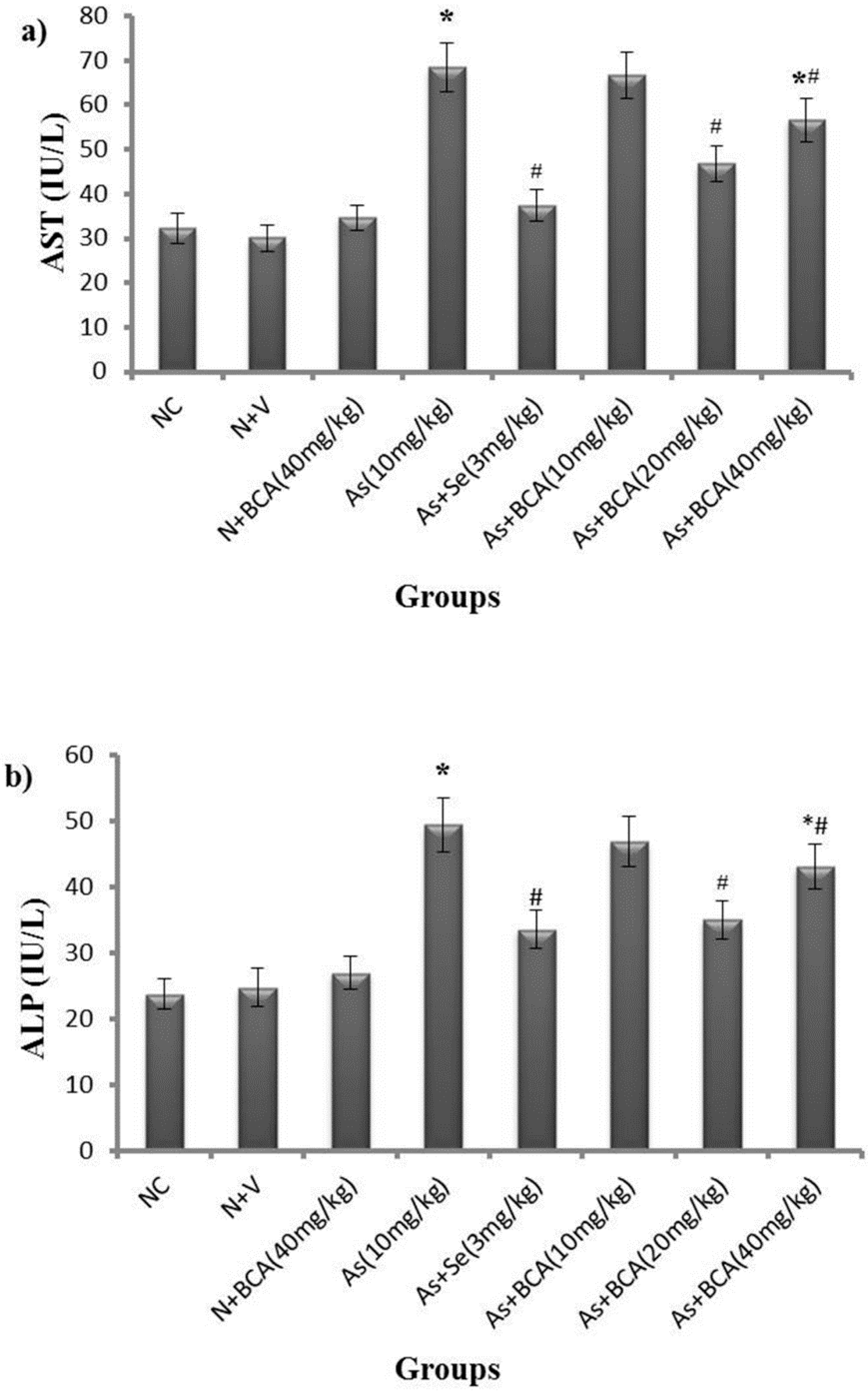
2.2. Hepatic Markers

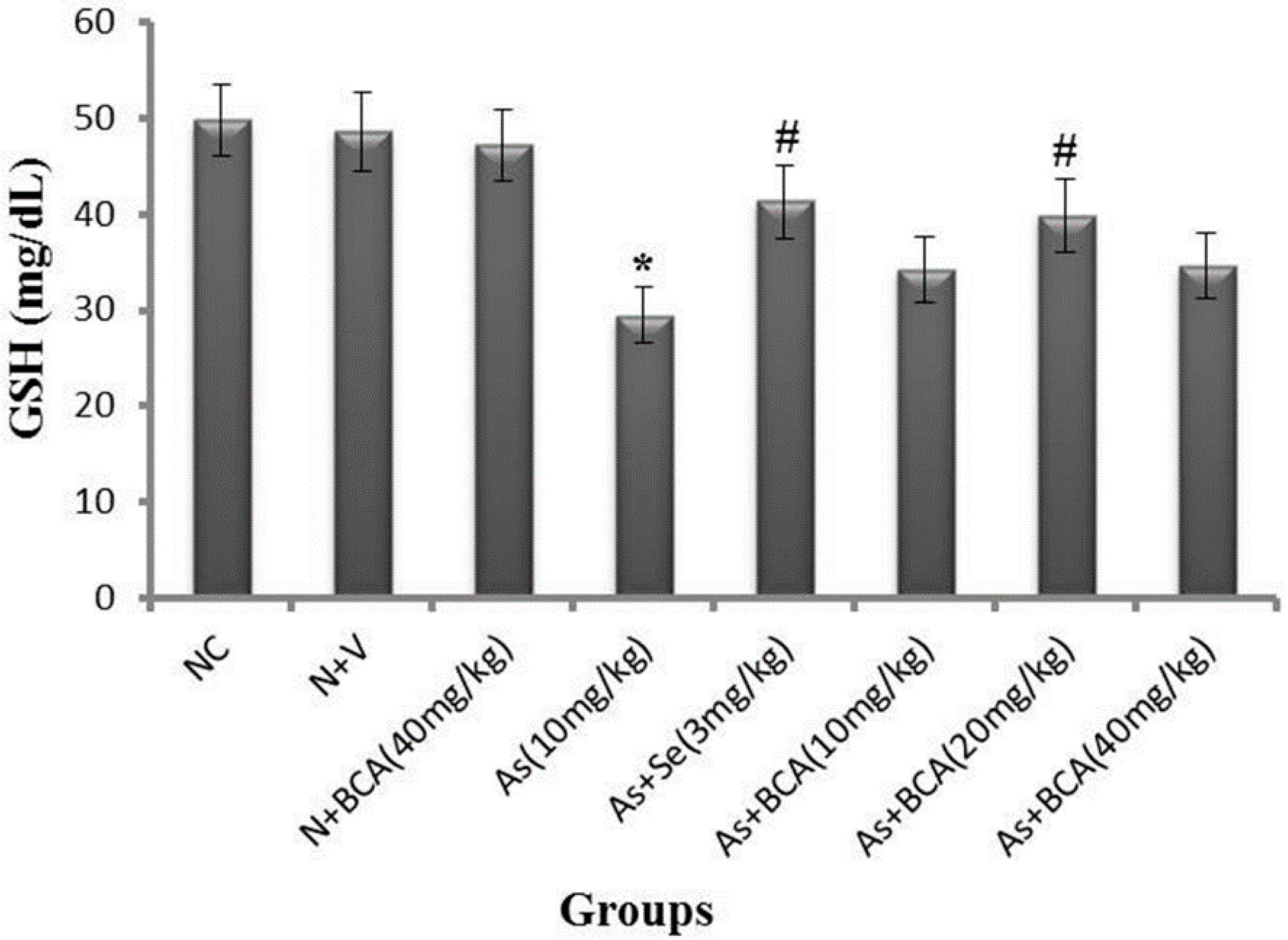
2.3. Plasma Reduced Glutathione Level

2.4. Oxidative Stress Markers
| Groups | GSH μM/mg Tissue | SOD U/mg Tissue | CAT U/mg Tissue | MDA nmole/mg Tissue |
|---|---|---|---|---|
| Normal control | 4.74 ± 0.26 | 3.47 ± 0.17 | 23.36 ± 0.26 | 0.78 ± 0.05 |
| Normal + Vehicle | 4.89 ± 0.86 | 3.69 ± 0.33 | 24.12 ± 0.02 | 0.81 ± 0.06 |
| Normal + BCA (40 mg/kg) | 4.95 ± 0.58 | 3.82 ± 0.10 | 25.04 ± 0.16 | 0.84 ± 0.07 |
| Arsenic (10 mg/kg) | 2.76 ± 0.10 * | 2.06 ± 0.15 * | 14.33 ± 0.06 * | 1.56 ± 0.02 * |
| Arsenic + Selenium (3 mg/kg) | 3.90 ± 0.24 # | 3.12 ± 0.05 # | 21.18 ± 0.15 # | 0.86 ± 0.02 # |
| Arsenic + BCA (10 mg/kg) | 2.98 ± 0.09 | 2.16 ± 0.04 | 16.59 ± 0.05 | 1.27 ± 0.03 |
| Arsenic + BCA (20 mg/kg) | 3.76 ± 0.23 # | 2.94 ± 0.10 # | 19.15 ± 0.04 # | 0.92 ± 0.02 # |
| Arsenic + BCA (40 mg/kg) | 3.37 ± 0.22 | 2.38 ± 0.05 | 18.44 ± 0.13 *# | 0.94 ± 0.04 # |
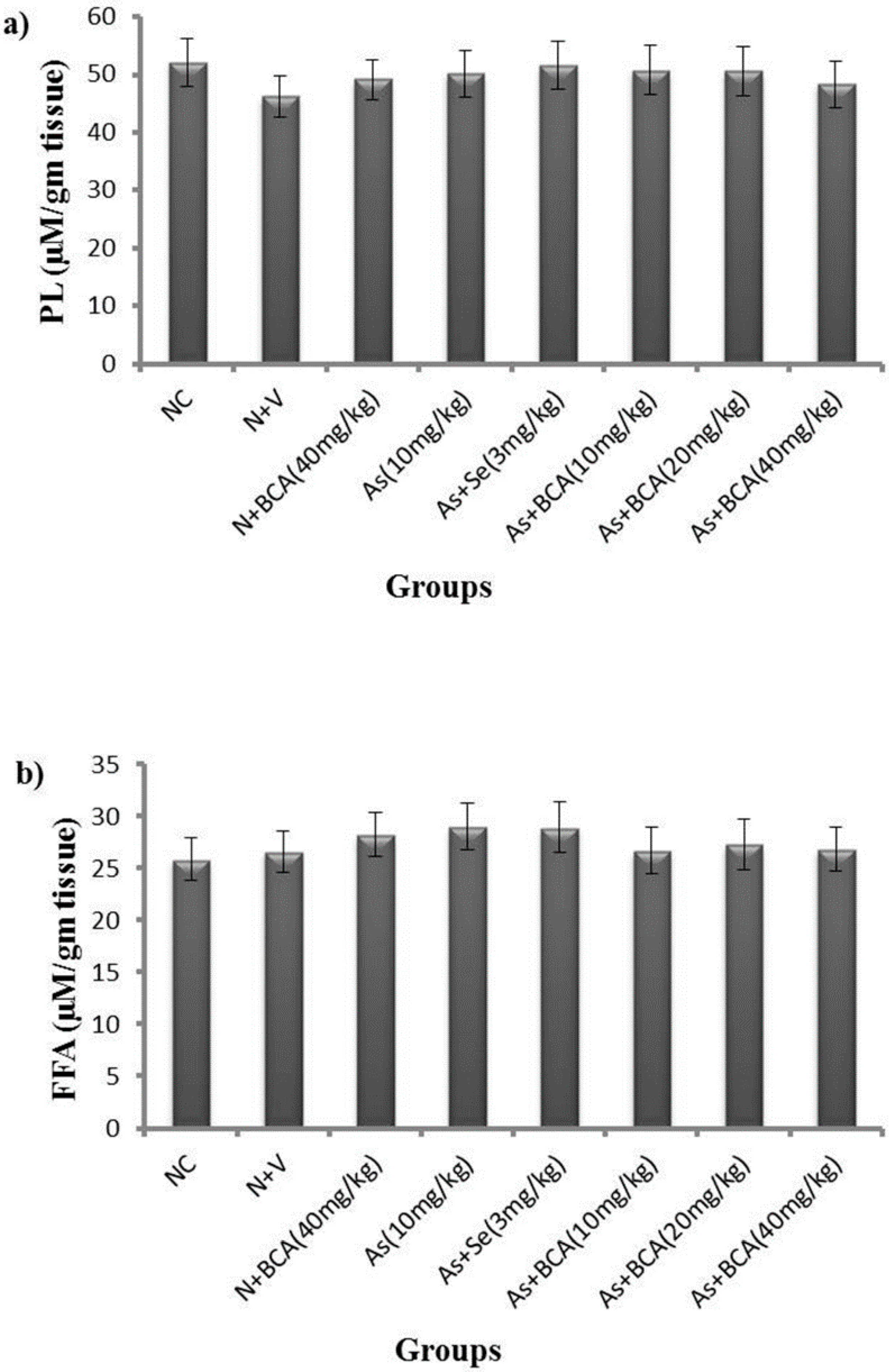
2.5. Phospholipids and Free Fatty Acid Levels

2.6. Hematological Indices
| Groups | Normal Control | Normal + Vehicle | Normal + BCA (40 mg/kg) | Arsenic (10 mg/kg) | Arsenic + Selenium (3 mg/kg) | Arsenic + BCA (10 mg/kg) | Arsenic + BCA (20 mg/kg) | Arsenic + BCA (40 mg/kg) |
|---|---|---|---|---|---|---|---|---|
| RBC count (×106 cells/L) | 8.44 ± 0.32 | 7.3 ± 0.36 * | 8.14 ± 0.38 | 8.14 ± 0.31 | 8.23 ± 0.31 | 8.67 ± 0.45 | 8.37 ± 0.52 | 8.27 ± 0.28 |
| Hb (g/dL) | 14.97 ± 1.12 | 14.51 ± 0.81 | 14.22 ± 0.73 | 14.5 ± 0.63 | 15.85 ± 1.20 # | 16.4 ± 0.52 # | 15.37 ± 0.84 | 15.17 ± 0.73 |
| Hematocrit (%) | 48.67 ± 3.51 | 47.15 ± 3.40 | 46.35 ± 3.82 | 44.52 ± 3.39 * | 47.77 ± 3.37 | 50.77 ± 4.04 # | 47.4 ± 3.10 | 45.87 ± 3.24 |
| MCV (fL) | 57.57 ± 4.35 | 57.57 ± 4.30 | 56.45 ± 4.12 | 55.35 ± 4.06 | 58.07 ± 4.64 | 57.3 ± 4.13 | 56.65 ± 4.68 | 56.47 ± 4.60 |
| MCH (pg) | 23.70 ± 1.50 | 23.85 ± 1.25 | 23.37 ± 1.38 | 17.40 ± 0.84 * | 23.97 ± 1.66 # | 24.27 ± 1.74 # | 24.32 ± 1.35 # | 23.87 ± 1.60 # |
| MCHC(g/dL) | 41.15 ± 3.98 | 41.45 ± 3.90 | 41.25 ± 3.47 | 43.85 ± 3.70 * | 41.92 ± 3.37 # | 42.14 ± 3.20 # | 42.84 ± 3.21 # | 43.02 ± 3.86 |
| CHCM (g/dL) | 32.25 ± 2.36 | 32.22 ± 2.66 | 31.82 ± 2.35 | 32.35 ± 2.26 | 31.25 ± 2.31 | 31.20 ± 2.14 | 31.92 ± 2.28 | 32.00 ± 2.18 |
| RDW (%) | 13.50 ± 0.29 | 13.20 ± 0.69 | 13.00 ± 0.42 | 13.45 ± 0.31 | 13.25 ± 0.12 | 13.02 ± 0.18 | 13.02 ± 0.45 | 13.00 ± 0.20 |
| HDW(g/dL) | 2.81 ± 0.07 | 2.65 ± 0.03 | 2.69 ± 0.08 | 2.95 ± 0.09 | 2.87 ± 0.08 | 2.89 ± 0.06 | 2.88 ± 0.09 | 2.71 ± 0.02 |
| Groups | Normal Control | Normal + Vehicle | Normal + BCA (40 mg/kg) | Arsenic (10 mg/kg) | Arsenic + Selenium (3 mg/kg) | Arsenic + BCA (10 mg/kg) | Arsenic + BCA (20 mg/kg) | Arsenic + BCA (40 mg/kg) |
|---|---|---|---|---|---|---|---|---|
| WBC count (×106 cells/L) | 2.96 ± 0.12 | 3.10 ± 0.45 | 3.49 ± 0.64 | 6.27 ± 0.75 * | 3.22 ± 0.37 # | 3.58 ± 0.28 # | 4.26 ± 0.31 # | 4.05 ± 0.31 # |
| Monocytes (%) | 2.25 ± 0.36 | 2.62 ± 0.31 | 2.50 ± 0.32 | 1.90 ± 0.59 | 2.32 ± 0.15 | 2.05 ± 0.40 | 2.25 ± 0.23 | 1.85 ± 0.44 |
| Easinophil(%) | 2.22 ± 0.17 | 2.17 ± 0.29 | 2.25 ± 0.62 | 1.30 ± 0.29 * | 1.98 ± 0.43 # | 0.82 ± 0.12 | 1.00 ± 0.08 | 1.22 ± 0.34 |
| Basophil (%) | 0.12 ± 0.02 | 0.10 ± 0.08 | 0.13 ± 0.08 | 0.10 ± 0.01 | 0.14 ± 0.09 | 0.12 ± 0.09 | 0.15 ± 0.05 | 0.10 ± 0.01 |
| Leucocytes (%) | 0.12 ± 0.05 | 0.20 ± 0.03 | 0.17 ± 0.05 | 0.22 ± 0.05 | 0.15 ± 0.05 | 0.12 ± 0.05 | 0.15 ± 0.05 | 0.12 ± 0.09 |
| Neutrophil (%) | 16.60 ± 0.98 | 13.52 ± 0.85 | 15.75 ± 1.24 | 26.1 ± 1.21 * | 18.77 ± 1.63 # | 23.27 ± 1.37 | 19.22 ± 1.28 # | 21.05 ± 1.99 |
| Lymphocytes (%) | 69.42 ± 4.58 | 81.55 ± 6.51 * | 77.75 ± 5.97 | 80.32 ± 6.83 * | 74.90 ± 5.50 | 80.87 ± 6.78 | 77.52 ± 5.42 | 75..62 ± 6.24 |
| Platelets (×106 cells/L) | 943 ± 83.44 | 917.25 ± 76.50 | 941.25 ± 75.56 | 616.50 ± 59.37 * | 1034.8 ± 98.77 # | 721.50 ± 66.92 | 936.50 ± 82.73 # | 864.50 ± 76.41 # |
| MPV(fL) | 10.12 ± 0.94 | 11.63 ± 0.94 | 10.5 ± 0.84 | 6.78 ± 0.83 * | 11.55 ± 0.80 # | 12.37 ± 0.79 # | 10.95 ± 0.67 # | 11.50 ± 0.45 # |
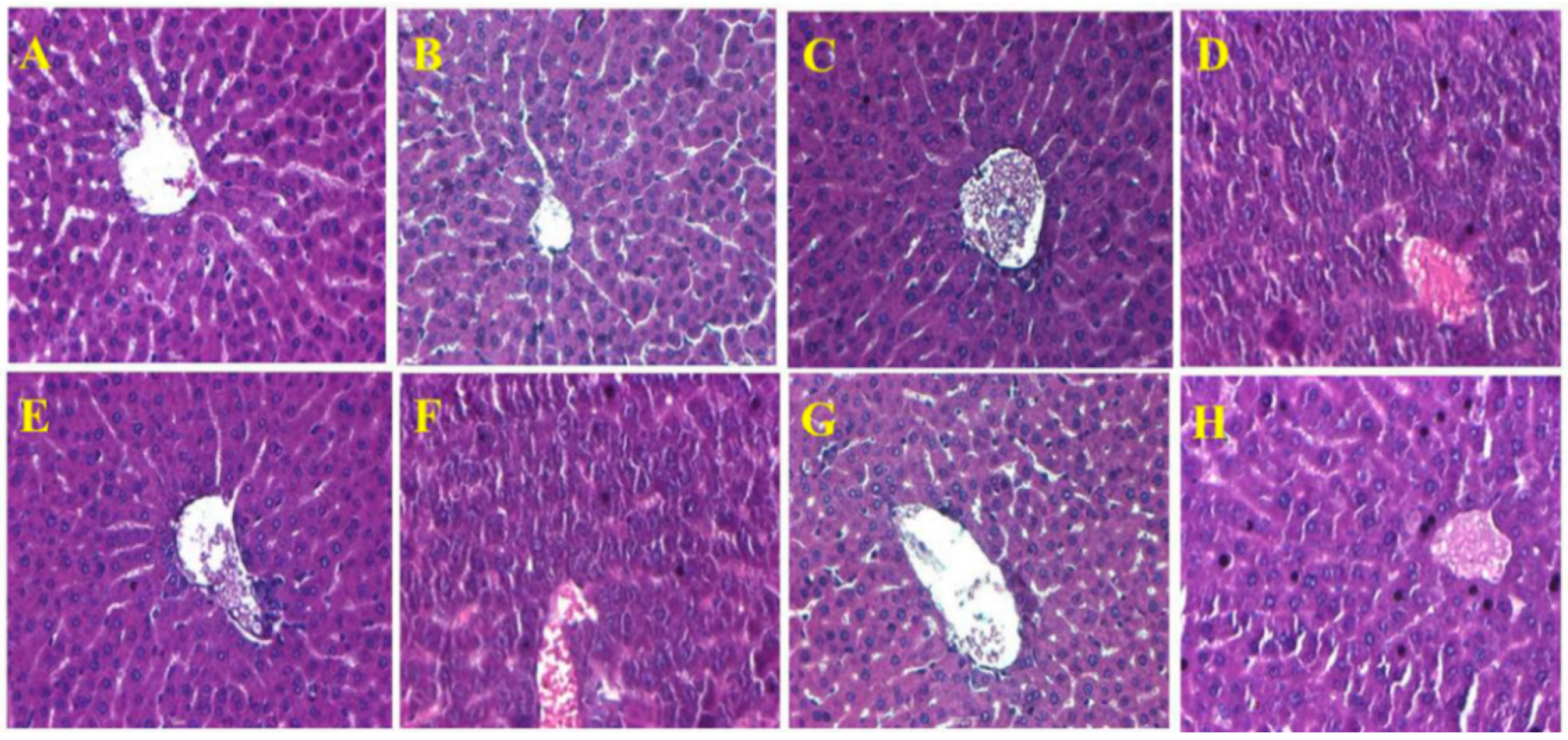
2.7. Histopathological Observations in the Liver

3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals and Reagents
4.3. Preparation and Administration of BCA, Selenium and Arsenic
4.4. Experimental Time Line
4.5. Collection of Samples
4.6. Measurement of Body Weight and Organ Body Weight Ratio
4.7. Assessment of Hepatic Damage
4.8. Estimation of Reduced Glutathione
4.9. Estimation of Lipid Peroxidation
4.10. Assay of Antioxidant Enzymes
4.11. Evaluation of Lipid Profiles
4.12. Clinical Hematological Variables
4.13. Morphological Studies of the Liver
4.14. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mandal, B.K.; Suzuki, K.T. Arsenic round the world: A review. Talanta 2002, 58, 201–235. [Google Scholar] [CrossRef]
- Shi, H.; Shi, X.; Liu, K.J. Oxidative mechanism of arsenic toxicity and carcinogenesis. Mol. Cell. Biochem. 2004, 255, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Hasegawa, H.; Rahman, M.M.; Miah, M.M.; Tasmin, A. Arsenic accumulation in rice (Oryza sativa L.): Human exposure through food chain. Ecotoxicol. Environ. Saf. 2008, 69, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Messarah, M.; Klibet, F.; Boumendjel, A.; Abdennour, C.; Bouzerna, N.; Boulakoud, M.S.; el Feki, A. Hepatoprotective role and antioxidant capacity of selenium on arsenic-induced liver injury in rats. Exp. Toxicol. Pathol. 2012, 64, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.; Mehta, A.; Gupta, R. Prevention of arsenic-induced hepatic apoptosis by concomitant administration of garlic extracts in mice. Chem. Biol. Interact. 2009, 177, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Brinkel, J.; Khan, M.H.; Kraemer, A. A systematic review of arsenic exposure and its social and mental health effects with special reference to bangladesh. Int. J. Environ. Res. Public Health 2009, 6, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Aposhian, H.V.; Aposhian, M.M. Newer developments in arsenic toxicity. Int. J. Toxicol. 1989, 8, 1297–1305. [Google Scholar] [CrossRef]
- Majhi, C.R.; Khan, S.; Leo, M.D.M.; Manimaran, A.; Sankar, P.; Sarkar, S.N. Effects of acetaminophen on reactive oxygen species and nitric oxide redox signaling in kidney of arsenic-exposed rats. Food Chem. Toxicol. 2011, 49, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Patra, P.H.; Bandyopadhyay, S.; Kumar, R.; Datta, B.K.; Maji, C.; Biswas, S.; Dash, J.R.; Sar, T.K.; Sarkar, S.; Manna, S.K. Quantitative imaging of arsenic and its species in goat following long term oral exposure. Food Chem. Toxicol. 2012, 50, 1946–1950. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, D.G. Effect of chronic intake of arsenic-contaminated water on liver. Toxicol. Appl. Pharmacol. 2005, 206, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, J.; Waalkes, M.P.; Cheng, M.L.; Li, L.; Li, C.X.; Yang, Q. High dietary fat exacerbates arsenic-induced liver fibrosis in mice. Exp. Biol. Med. 2008, 233, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Dutta, M.; Ghosh, D.; Ghosh, A.K.; Bose, G.; Chattopadhyay, A.; Rudra, S.; Dey, M.; Bandyopadhyay, A.; Pattari, S.K.; Mallick, S. High fat diet aggravates arsenic induced oxidative stress in rat heart and liver. Food Chem. Toxicol. 2014, 66, 262–277. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.K.; Dwivedi, S.; Yadav, S.S.; Sharma, P.; Khattri, S. Arsenic-induced hepatic toxicity and its attenuation by fruit extract of Emblica officinalis (amla) in mice. Indian J. Clin. Biochem. 2014, 29, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Dowling, S.; Regan, F.; Hughes, H. The characterisation of structural and antioxidant properties of isoflavone metal chelates. J. Inorg. Biochem. 2010, 104, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lin, S.; Li, Z.; Yang, D.; Wang, Z. Protective effects of puerarin on experimental chronic lead nephrotoxicity in immature female rats. Hum. Exp. Toxicol. 2013, 32, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Cassady, J.M.; Zennie, T.M.; Chae, Y.H.; Ferin, M.A.; Portuondo, N.E.; Baird, W.M. Use of a mammalian cell culture benzo (a) pyrene metabolism assay for the detection of potential anticarcinogens from natural products: Inhibition of metabolism by biochanin a, an isoflavone from trifolium pratense l. Cancer Res. 1988, 48, 6257–6261. [Google Scholar] [PubMed]
- Chung, M.J.; Sohng, J.K.; Choi, D.J.; Park, Y.I. Inhibitory effect of phloretin and biochanin a on ige-mediated allergic responses in rat basophilic leukemia rbl-2h3 cells. Life Sci. 2013, 93, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Sithisarn, P.; Michaelis, M.; Schubert-Zsilavecz, M.; Cinatl, J. Differential antiviral and anti-inflammatory mechanisms of the flavonoids biochanin a and baicalein in h5n1 influenza a virus-infected cells. Antivir. Res. 2013, 97, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.W.; Tham, C.L.; Israf, D.A.; Lee, S.H.; Kim, M.K. Neuroprotective effects of biochanin a against glutamate-induced cytotoxicity in pc12 cells via apoptosis inhibition. Neurochem. Res. 2013, 38, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Jalaludeen, A.M.; Lee, W.Y.; Kim, J.H.; Jeong, H.Y.; Ki, K.S.; Kwon, E.G.; Song, H. Therapeutic efficacy of biochanin a against arsenic-induced renal and cardiac damage in rats. Environ. Toxicol. Pharmacol. 2015, 39, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Breikaa, R.M.; Algandaby, M.M.; El-Demerdash, E.; Abdel-Naim, A.B. Biochanin a protects against acute carbon tetrachloride-induced hepatotoxicity in rats. Biosci. Biotechnol. Biochem. 2013, 77, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tang, L.J.; Sun, Y.N.; Qiu, P.H.; Liang, G. Syntheses, characterization and antitumor activities of transition metal complexes with isoflavone. J. Inorg. Biochem. 2010, 104, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Gailer, J.; George, G.N.; Pickering, I.J.; Prince, R.C.; Ringwald, S.C.; Pemberton, J.E.; Glass, R.S.; Younis, H.S.; DeYoung, D.W.; Aposhian, H.V. A metabolic link between arsenite and selenite: The seleno-bis (s-glutathionyl) arsinium ion. J. Am. Chem. Soc. 2000, 122, 4637–4639. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, Z.; Li, J.J.; Chen, C.; Zhang, P.C.; Dong, L.; Chen, J.H.; Chen, Q.; Zhang, X.T.; Wang, Z.L. Protective effects of selenium on oxidative damage and oxidative stress related gene expression in rat liver under chronic poisoning of arsenic. Food Chem. Toxicol. 2013, 58, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Ferreira, D. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Dalal, B.; Nayak, K.; Gangopadhyay, P.; Mukherjee, A. Identification of indicators of arsenic induced hepatic damage in human. Internet J. Toxicol. 2009, 7. [Google Scholar]
- Miltonprabu, S.; Sumedha, N. Arsenic-induced hepatic mitochondrial toxicity in rats and its amelioration by diallyl trisulfide. Toxicol. Mech. Methods 2014, 24, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Kotyzová, D.; Bludovská, M.; Eybl, V. Differential influences of various arsenic compounds on antioxidant defense system in liver and kidney of rats. Environ. Toxicol. Pharmacol. 2013, 36, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Muthumani, M. Tetrahydrocurcumin potentially attenuates arsenic induced oxidative hepatic dysfunction in rats. J. Clin. Toxicol. 2013, 3. [Google Scholar] [CrossRef]
- Bhattacharjee, A.; Basu, A.; Ghosh, P.; Biswas, J.; Bhattacharya, S. Protective effect of selenium nanoparticle against cyclophosphamide induced hepatotoxicity and genotoxicity in swiss albino mice. J. Biomater. Appl. 2014, 29, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Kale, R.; Kar, A. Chemoprevention of mammary tumorigenesis and chemomodulation of the antioxidative enzymes and peroxidative damage in prepubertal sprague dawley rats by biochanin a. Mol. Cell. Biochem. 2008, 312, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.N.; Niedzwiecki, M.M.; Liu, X.; Harper, K.N.; Alam, S.; Slavkovich, V.N.; Ilievski, V.; Levy, D.K.; Siddique, A.B.; Parvez, M.F. Chronic arsenic exposure and blood glutathione and glutathione disulfide concentrations in bangladeshi adults. Environ. Health Perspect. 2013, 121, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Song, L.L.; Tu, Y.Y.; Xia, L.; Wang, W.W.; Wei, W.; Ma, C.M.; Wen, D.H.; Lei, H.; Xu, H.Z.; Wu, Y.L. Targeting catalase but not peroxiredoxins enhances arsenic trioxide-induced apoptosis in k562 cells. PLoS ONE 2014, 9, e104985. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Hasegawa, A.; Sawamura, R.; Okada, S. Cellular response to oxidative damage in lung induced by the administration of dimethylarsinic acid, a major metabolite of inorganic arsenics, in mice. Toxicol. Appl. Pharmacol. 1991, 108, 205–213. [Google Scholar] [CrossRef]
- Rana, T.; Bera, A.K.; Das, S.; Bhattacharya, D.; Pan, D.; Bandyopadhyay, S.; Mondal, D.K.; Samanta, S.; Bandyopadhyay, S.; Das, S.K. Pleurotus florida lectin normalizes duration dependent hepatic oxidative stress responses caused by arsenic in rat. Exp. Toxicol. Pathol. 2012, 64, 665–671. [Google Scholar] [CrossRef]
- Zhang, W.; Xue, J.; Ge, M.; Yu, M.; Liu, L.; Zhang, Z. Resveratrol attenuates hepatotoxicity of rats exposed to arsenic trioxide. Food Chem. Toxicol. 2013, 51, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Flora, G.; Bhatnagar, P.; Flora, S. Comparative oxidative stress, metallothionein induction and organ toxicity following chronic exposure to arsenic, lead and mercury in rats. Cell. Mol. Biol. (Noisy-le-Grand) 2014, 60, 13–21. [Google Scholar]
- Sárközi, K.; Papp, A.; Horváth, E.; Máté, Z.; Ferencz, Á.; Hermesz, E.; Krisch, J.; Paulik, E.; Szabó, A. Green tea and vitamin C ameliorate some neuro-functional and biochemical signs of arsenic toxicity in rats. Nutr. Neurosci. 2014. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Duan, X.; Wang, X.; Dong, D.; Liu, D.; Li, X.; Sun, G.; Li, B. Curcumin attenuates arsenic-induced hepatic injuries and oxidative stress in experimental mice through activation of nrf2 pathway, promotion of arsenic methylation and urinary excretion. Food Chem. Toxicol. 2013, 59, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Tayman, C.; Cekmez, F.; Kafa, I.M.; Canpolat, F.E.; Cetinkaya, M.; Tonbul, A.; Uysal, S.; Tunc, T.; Sarici, S.U. Protective effects of nigella sativa oil in hyperoxia-induced lung injury. Arch. Bronconeumol. 2013, 49, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Muthumani, M.; Prabu, S.M. Silibinin potentially attenuates arsenic-induced oxidative stress mediated cardiotoxicity and dyslipidemia in rats. Cardiovasc. Toxicol. 2014, 14, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.J.; Chuu, J.J.; Chang, C.Y.; Tsai, W.C.; Chen, K.J.; Guo, H.R. Atherosclerosis induced by arsenic in drinking water in rats through altering lipid metabolism. Toxicol. Appl. Pharmacol. 2011, 256, 146–153. [Google Scholar] [PubMed]
- Orazizadeh, M.; Fakhredini, F.; Mansouri, E.; Khorsandi, L. Effect of glycyrrhizic acid on titanium dioxide nanoparticles-induced hepatotoxicity in rats. Chem. Biol. Interact. 2014, 220, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, N.; Flora, G.; Kushwaha, P.; Flora, S.J. Alpha-lipoic acid protects oxidative stress, changes in cholinergic system and tissue histopathology during co-exposure to arsenic-dichlorvos in rats. Environ. Toxicol. Pharmacol. 2014, 37, 7–23. [Google Scholar] [PubMed]
- Dwivedi, N.; Flora, S.J. Concomitant exposure to arsenic and organophosphates on tissue oxidative stress in rats. Food Chem. Toxicol. 2011, 49, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Kannan, G.M.; Sharma, M.; Flora, S.J. Therapeutic effects of moringa oleifera on arsenic-induced toxicity in rats. Environ. Toxicol. Pharmacol. 2005, 20, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.J.; Sagawa, K.; Frederick, K.; Zhang, S.; Morris, M.E. Pharmacokinetics and bioavailability of the isoflavone biochanin a in rats. AAPS J. 2006, 8, E433–E442. [Google Scholar] [CrossRef] [PubMed]
- Barnes, S. Phyto-oestrogens and osteoporosis: What is a safe dose? Br. J. Nutr. 2003, 89, S101–S108. [Google Scholar] [CrossRef] [PubMed]
- Sárközi, K.; Máté, Z.; Vezér, T. General and behavioral toxicological effects of subchronic inorganic arsenic and fluoride treatment in adult wistar rats. Eur. Chem. Bull. 2012, 1, 531–534. [Google Scholar]
- Sample Availability: Not Available
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalaludeen, A.M.; Ha, W.T.; Lee, R.; Kim, J.H.; Do, J.T.; Park, C.; Heo, Y.T.; Lee, W.Y.; Song, H. Biochanin A Ameliorates Arsenic-Induced Hepato- and Hematotoxicity in Rats. Molecules 2016, 21, 69. https://doi.org/10.3390/molecules21010069
Jalaludeen AM, Ha WT, Lee R, Kim JH, Do JT, Park C, Heo YT, Lee WY, Song H. Biochanin A Ameliorates Arsenic-Induced Hepato- and Hematotoxicity in Rats. Molecules. 2016; 21(1):69. https://doi.org/10.3390/molecules21010069
Chicago/Turabian StyleJalaludeen, Abdulkadhar Mohamed, Woo Tae Ha, Ran Lee, Jin Hoi Kim, Jeong Tae Do, Chankyu Park, Young Tae Heo, Won Young Lee, and Hyuk Song. 2016. "Biochanin A Ameliorates Arsenic-Induced Hepato- and Hematotoxicity in Rats" Molecules 21, no. 1: 69. https://doi.org/10.3390/molecules21010069
APA StyleJalaludeen, A. M., Ha, W. T., Lee, R., Kim, J. H., Do, J. T., Park, C., Heo, Y. T., Lee, W. Y., & Song, H. (2016). Biochanin A Ameliorates Arsenic-Induced Hepato- and Hematotoxicity in Rats. Molecules, 21(1), 69. https://doi.org/10.3390/molecules21010069