
The Protective Effects of HJB-1, a Derivative of 17-Hydroxy-Jolkinolide B, on LPS-Induced Acute Distress Respiratory Syndrome Mice

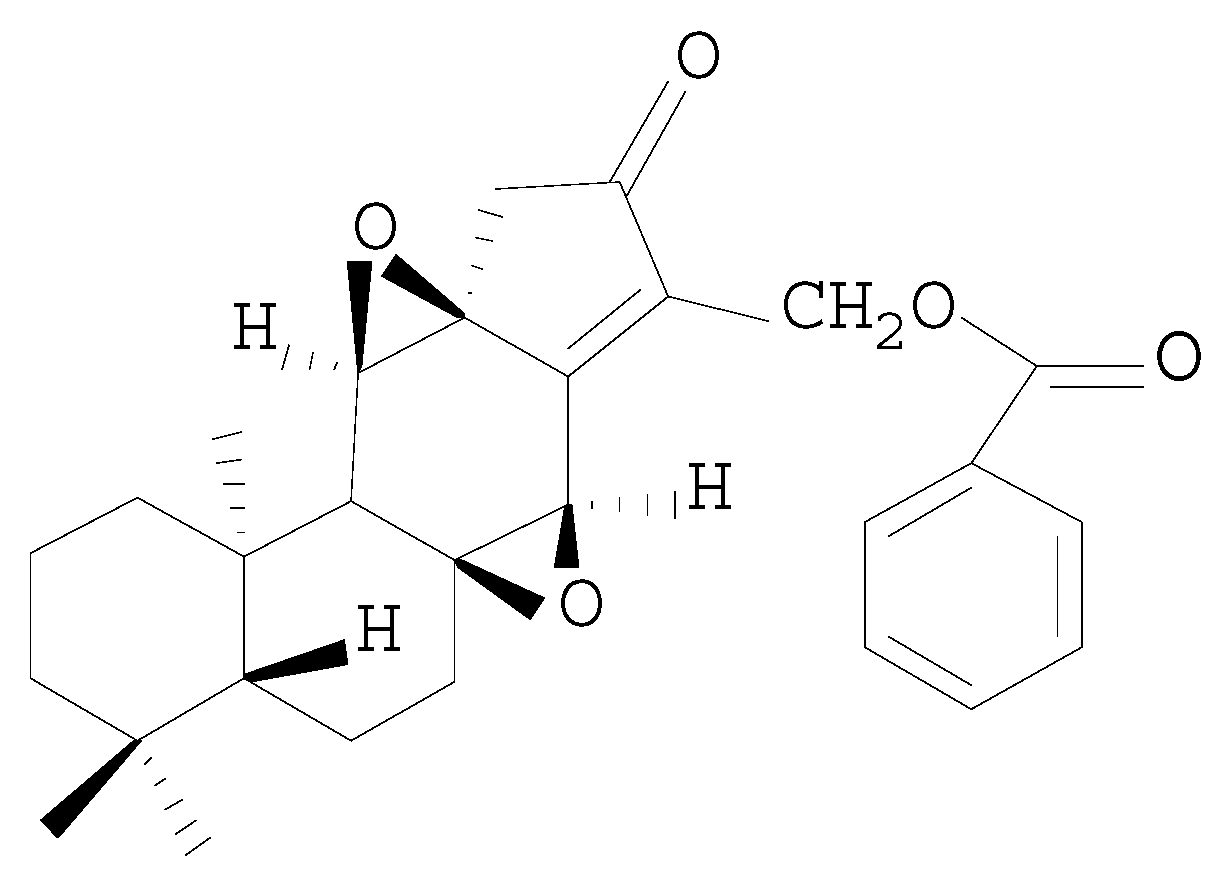{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
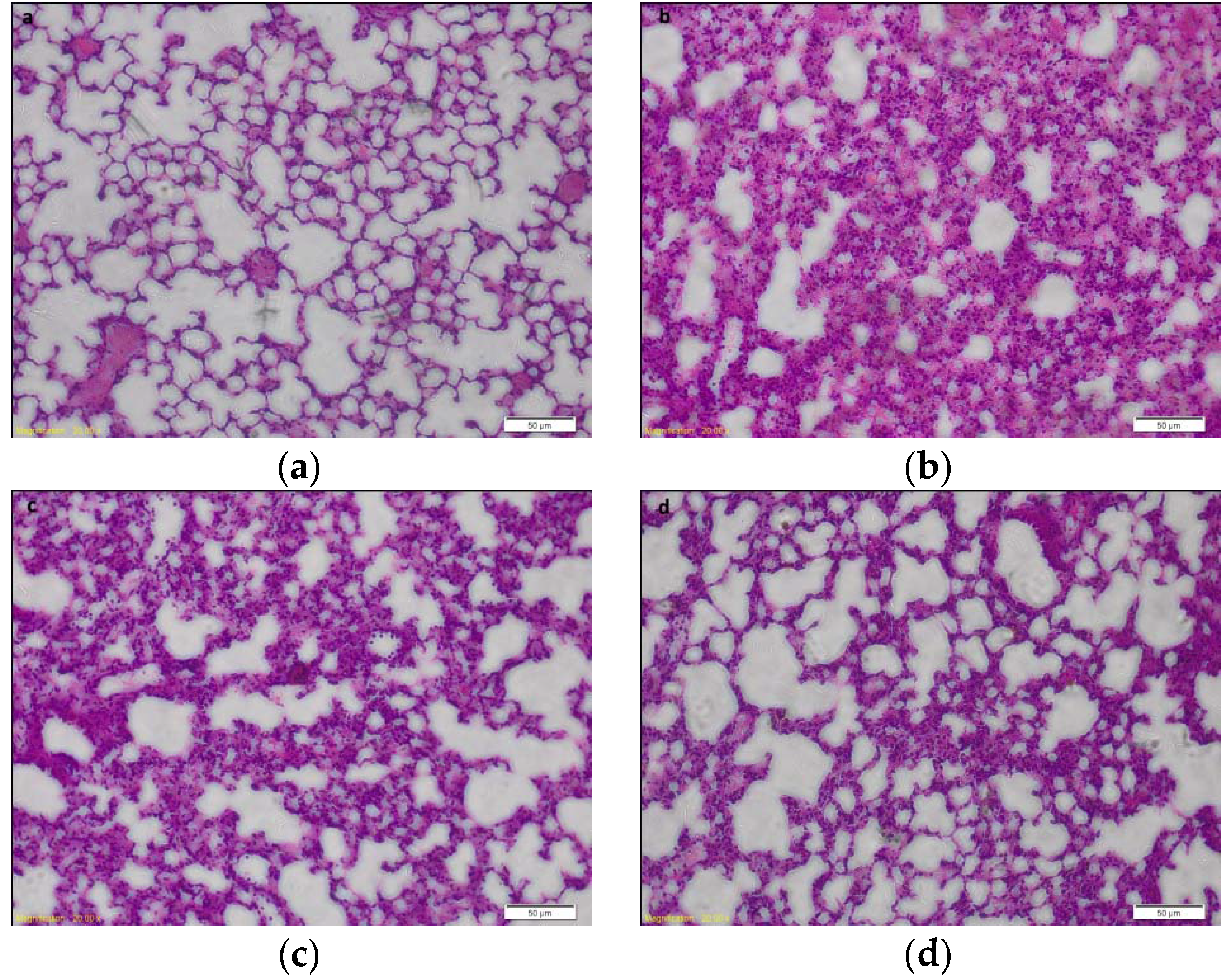
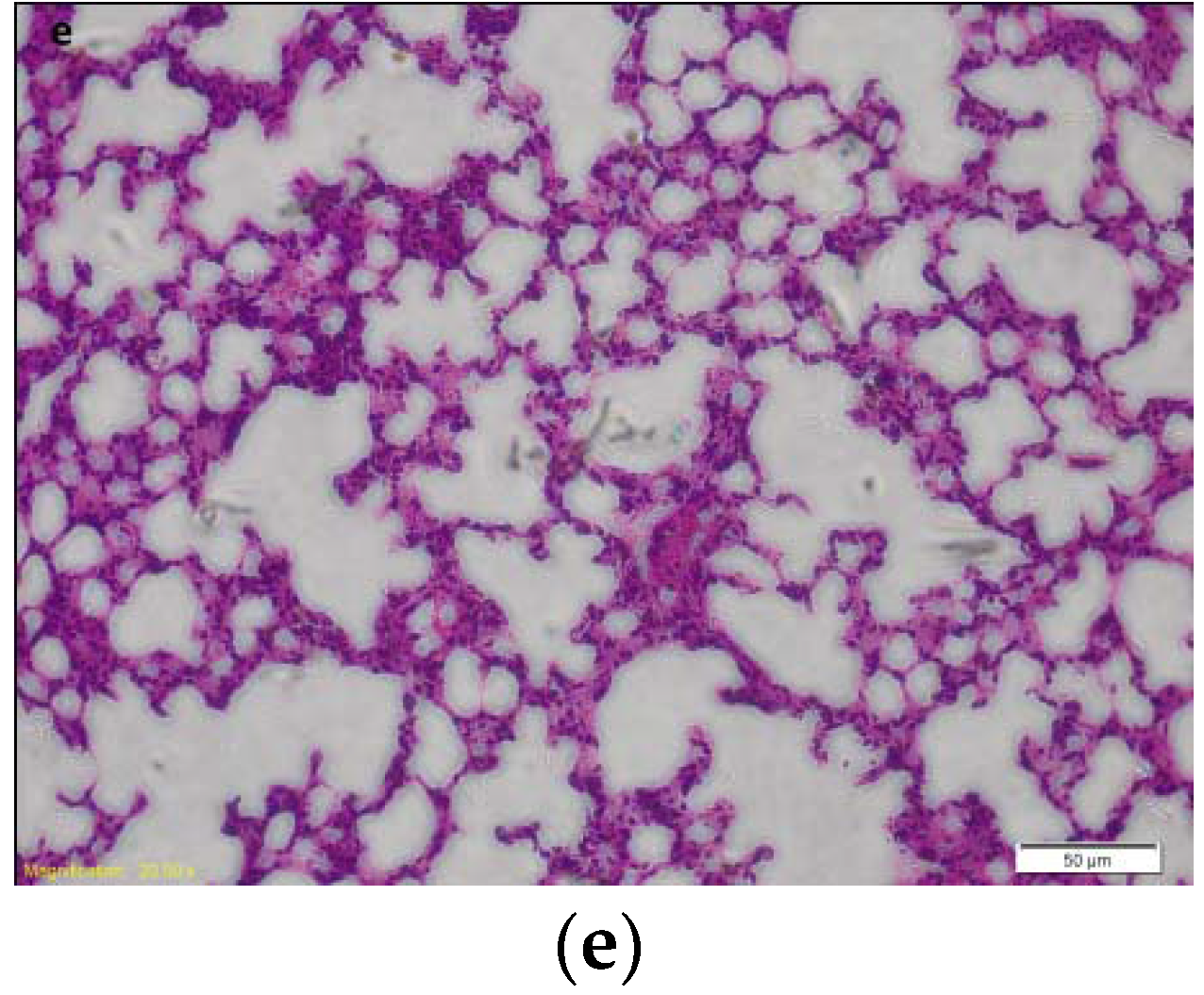
2.1. Effects of HJB-1 on LPS-Induced Lung Histopathologic Changes



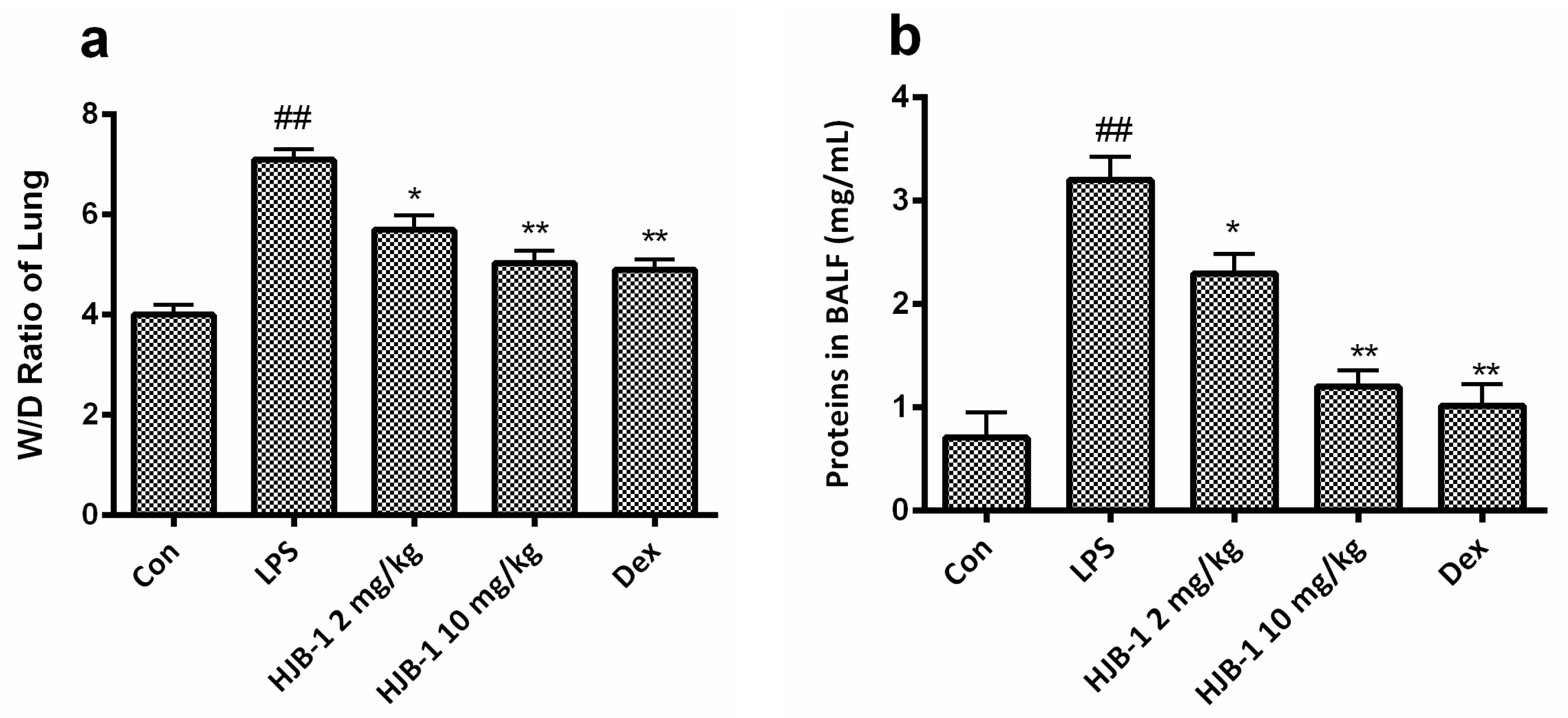
2.2. HJB-1 Attenuated LPS-Induced Pulmonary Vascular Permeability and Lung Edema

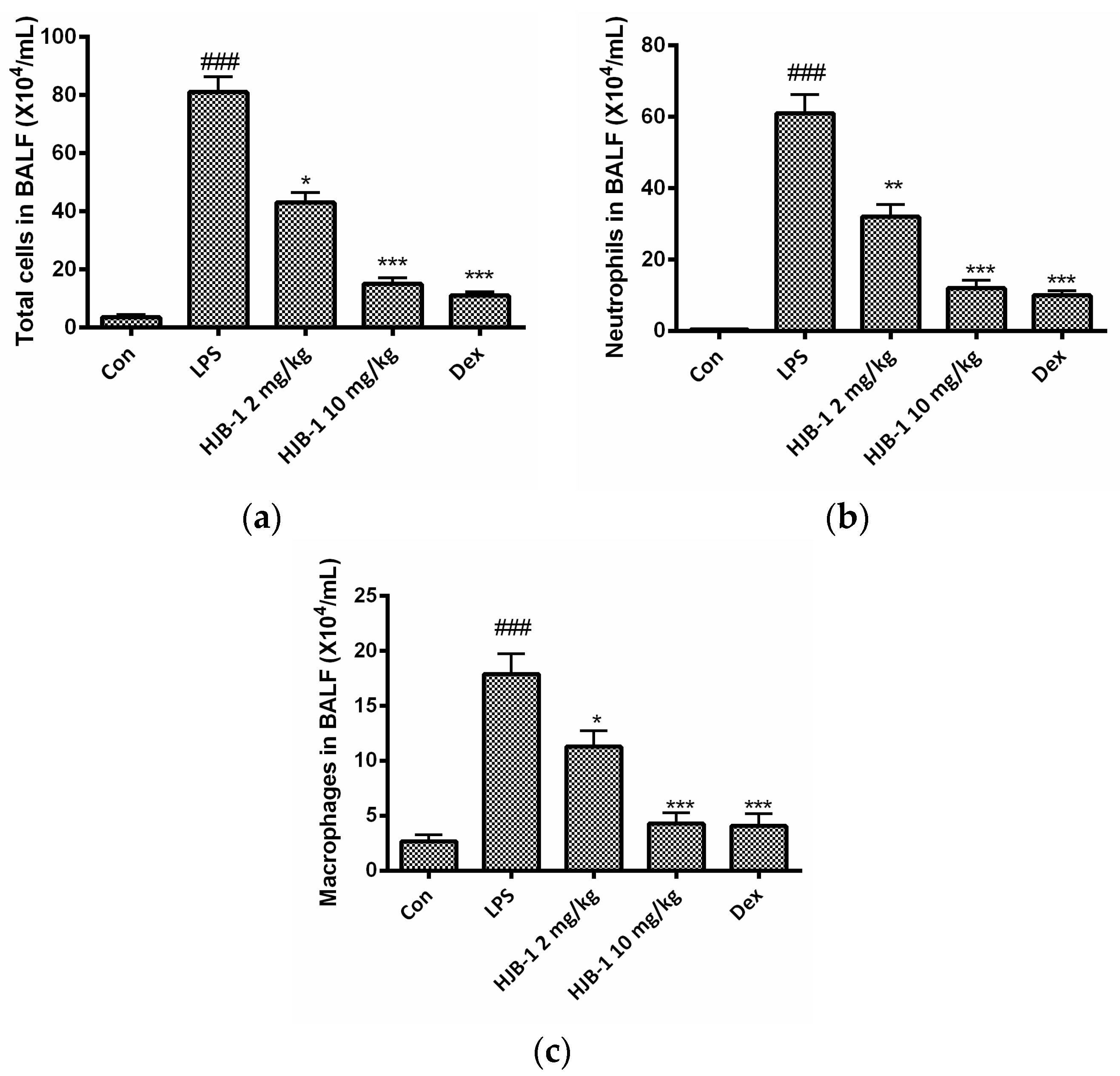
2.3. HJB-1 Reduced Total Cells Number in the BALF of LPS-Induced ARDS Mice

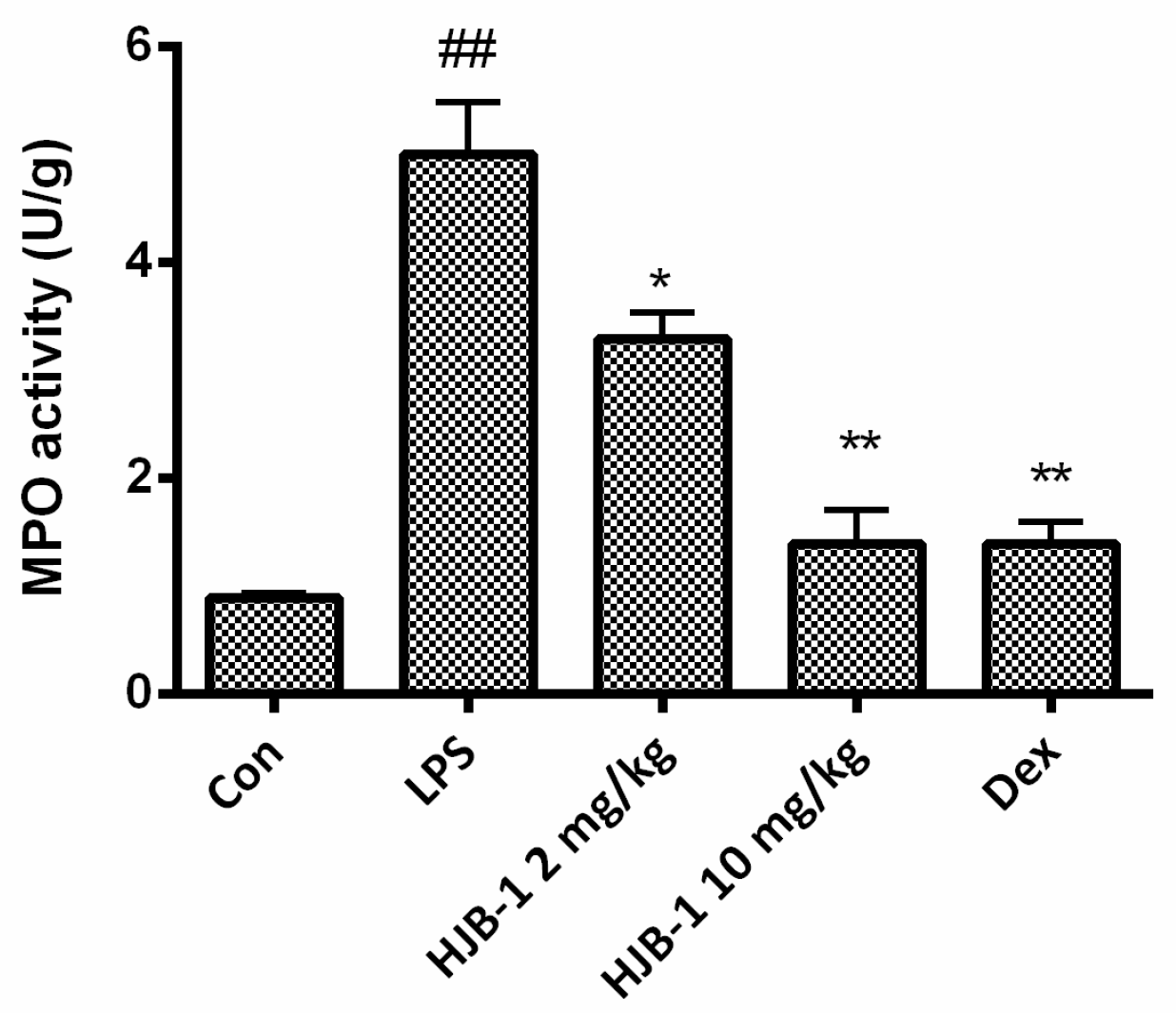
2.4. HJB-1 Alleviated the MPO Activity of Lung Tissues in LPS-Induced ARDS Mice

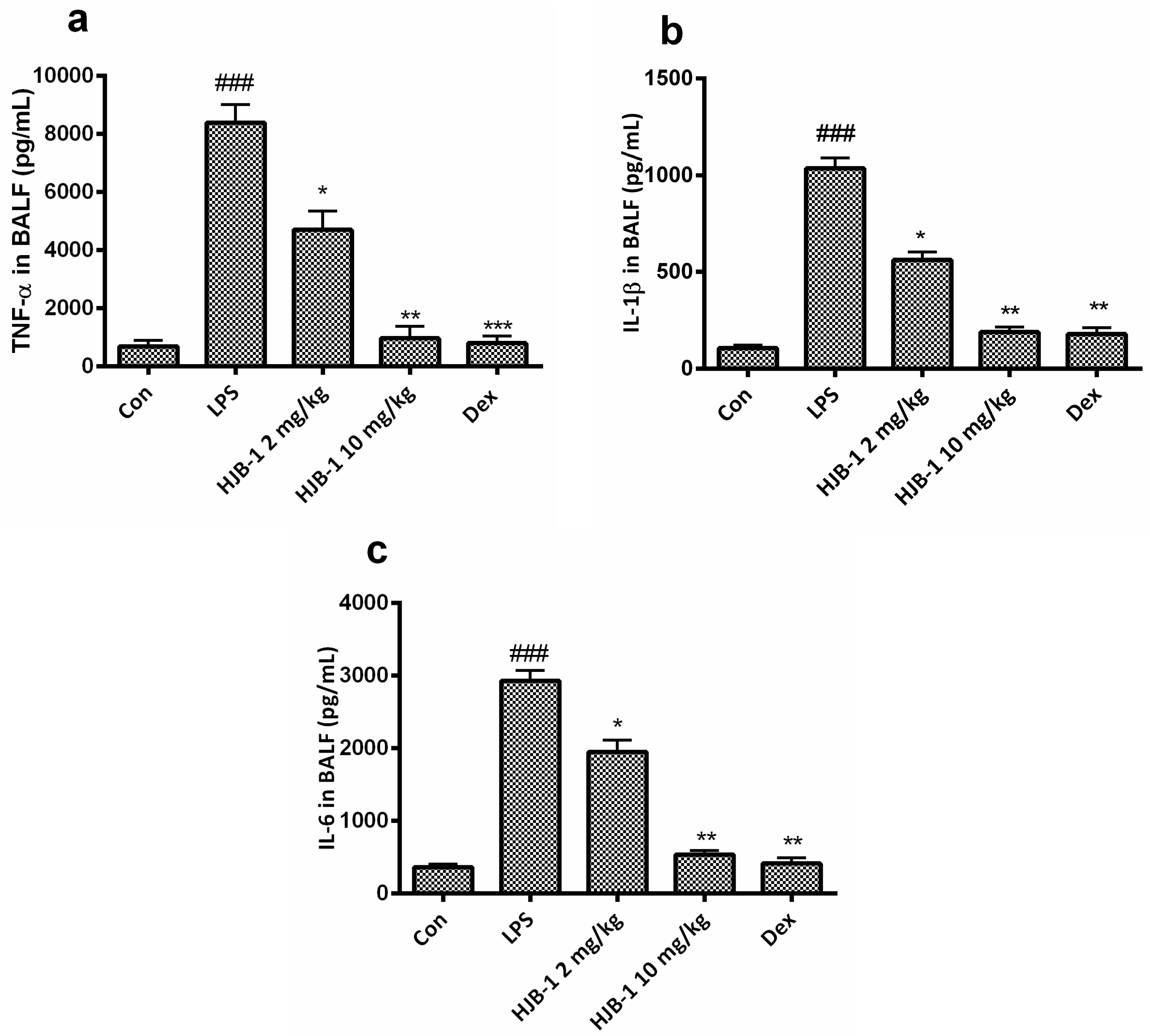
2.5. Effects of HJB-1 on Inflammatory Cytokines Production in BALF of LPS-Induced ARDS Mice

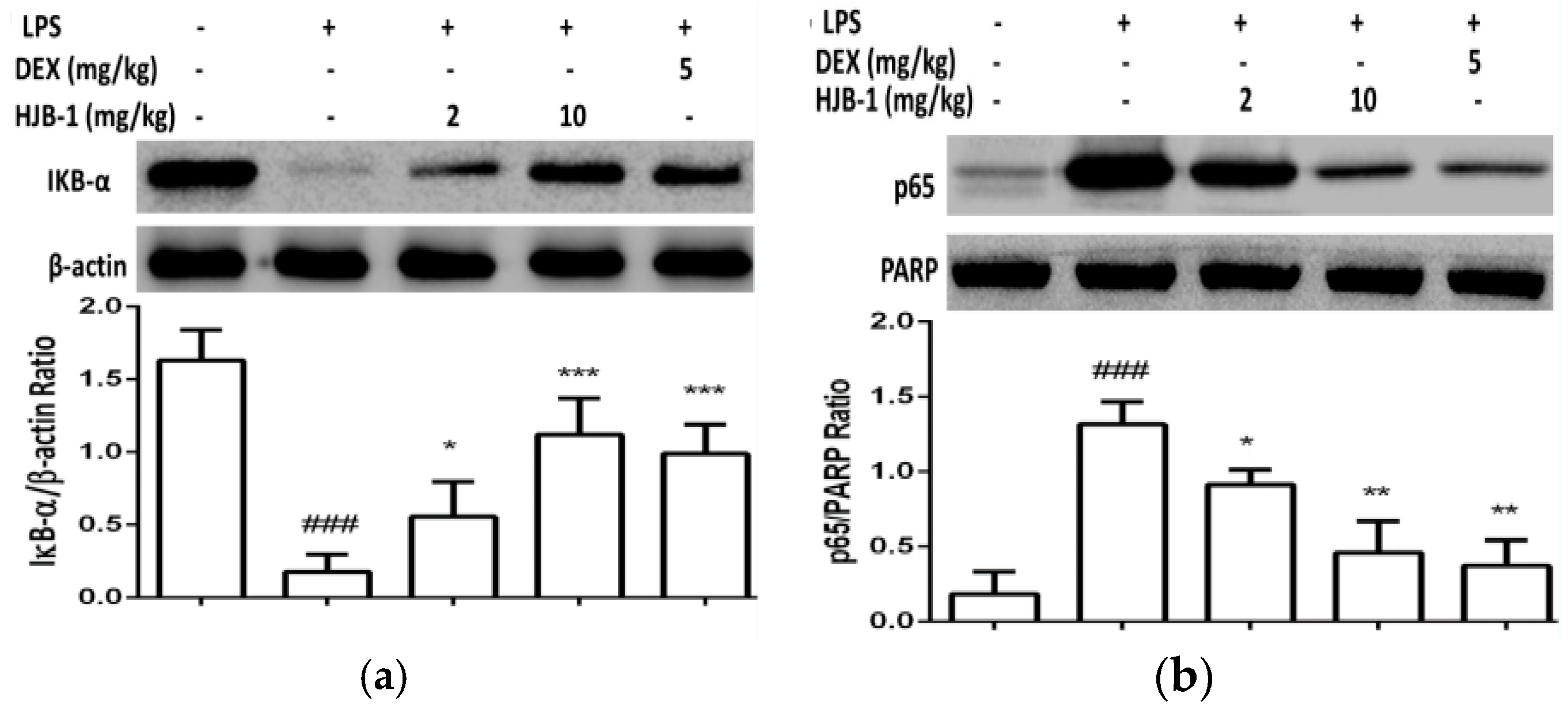
2.6. Effects of HJB-1 on NF-κB Activation in LPS-Induced ARDS Mice
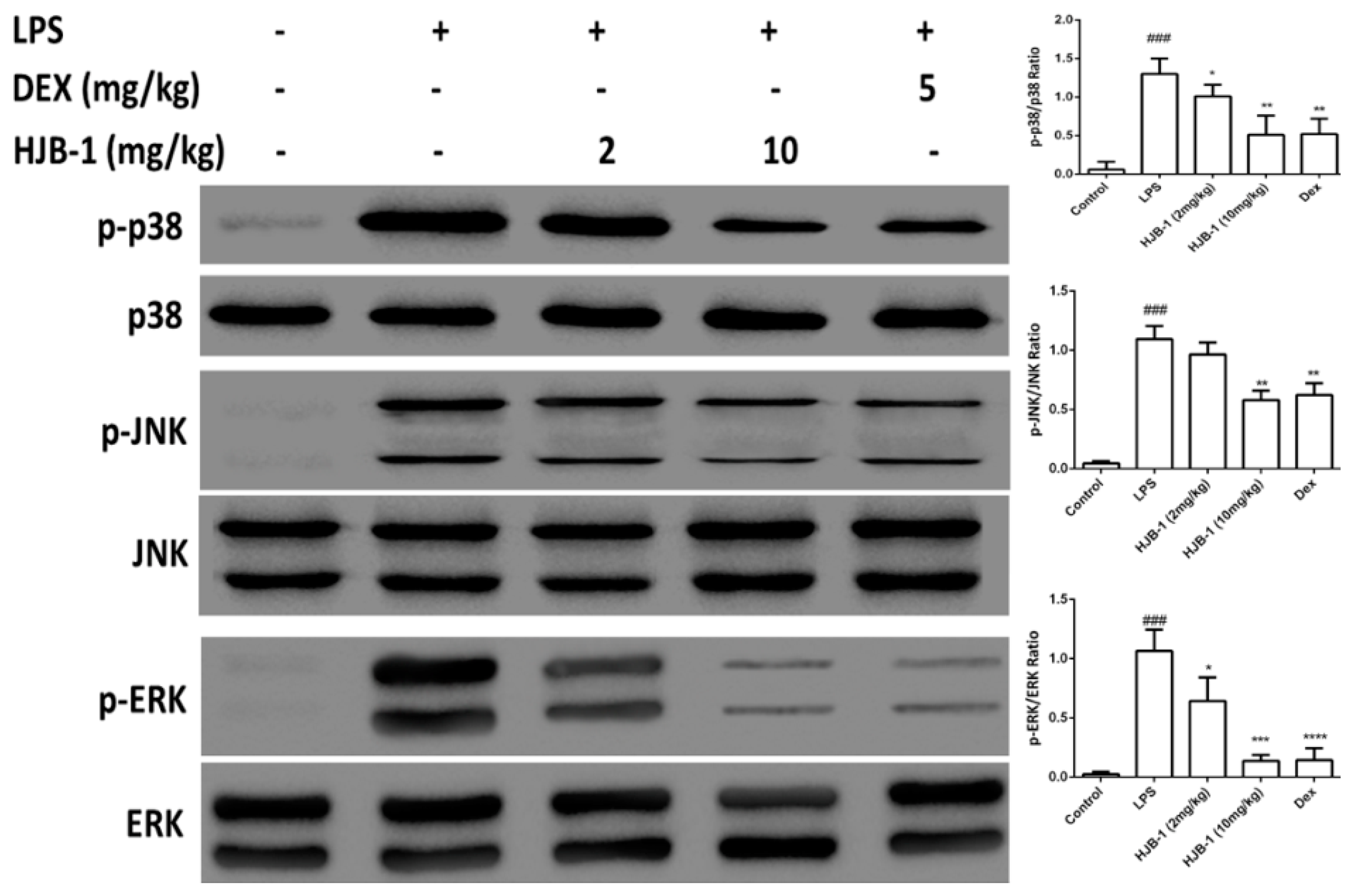
2.7. Effects of HJB-1 on MAPK Phosphorylation in LPS-Induced ARDS Mice


3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals
4.3. Histopathologic Evaluation of the Lung Tissue
4.4. Wet-to-Dry Lung Weight Ratio (W/D Ratio)
4.5. BALF Collection and Cell Count
4.6. MPO Activity Assay
4.7. Measurement of Inflammatory Cytokines in BALF
4.8. Western Blotting
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Standiford, T.J.; Ward, P.A. Therapeutic targeting of acute lung injury and acute respiratory distress syndrome. Transl. Res. 2015, 167, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Ware, L.B.; Zimmerman, G.A. The acute respiratory distress syndrome. J. Clin. Investig. 2012, 122, 2731–2740. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Zemans, R.L. The acute respiratory distress syndrome: pathogenesis and treatment. Annu. Rev. Pathol. 2011, 6, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Ware, L.B.; Matthay, M.A. The acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1334–1349. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Kewley, E.M.; Brodsky, K.S.; Tak, E.; Bonney, S.; Gobel, M.; Anderson, D.; Glover, L.E.; Riegel, A.K.; Colgan, S.P.; et al. Identification of hypoxia-inducible factor HIF-1A as transcriptional regulator of the A2B adenosine receptor during acute lung injury. J. Immunol. 2014, 192, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Szarka, R.J.; Wang, N.; Gordon, L.; Nation, P.N.; Smith, R.H. A murine model of pulmonary damage induced by lipopolysaccharide via intranasal instillation. J. Immunol. Methods 1997, 202, 49–57. [Google Scholar] [CrossRef]
- Lin, M.H.; Chen, M.C.; Chen, T.H.; Chang, H.Y.; Chou, T.C. Magnolol ameliorates lipopolysaccharide-induced acute lung injury in rats through PPAR-gamma-dependent inhibition of NF-kB activation. Int. Immunopharmacol. 2015, 28, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Liu, G.; Sun, R.; Wang, L.; Wang, J.; Wang, H. Fucosterol attenuates lipopolysaccharide-induced acute lung injury in mice. J. Surg. Res. 2015, 195, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, Y.; Huo, P.; Li, X.O.; Kong, D.; Mu, W.; Fang, W.; Li, L.; Liu, N.; Fang, L.; et al. Protective effect of Jolkinolide B on LPS-induced mouse acute lung injury. Int. Immunopharmacol. 2015, 26, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Wang, D.X.; Zhang, W.; Liao, X.Q.; Guan, X.; Bo, H.; Sun, J.Y.; Huang, N.W.; He, J.; Zhang, Y.K.; et al. Andrographolide protects against LPS-induced acute lung injury by inactivation of NF-kappaB. PLoS ONE 2013, 8, e56407. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, X.; Yan, S.; Shen, S.; Zhu, H.; Gu, Y.; Wang, H.; Qin, G.; Yu, Q. 17-hydroxy-jolkinolide B inhibits signal transducers and activators of transcription 3 signaling by covalently cross-linking Janus kinases and induces apoptosis of human cancer cells. Cancer Res. 2009, 69, 7302–7310. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.C.; Xu, X.H.; Zhang, N.N.; Liu, N.; Wu, D.L.; Wang, Y.; Peng, Q.S.; Vandenplas, M.; Wang, H.B.; Sun, W.C. HJB-1, a 17-hydroxy-jolkinolide B derivative, inhibits LPS-induced inflammation in mouse peritoneal macrophages. Int. Immunopharmacol. 2014, 21, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Soromou, L.W.; Chen, N.; Jiang, L.; Huo, M.; Wei, M.; Chu, X.; Millimouno, F.M.; Feng, H.; Sidime, Y.; Deng, X. Astragalin attenuates lipopolysaccharide-induced inflammatory responses by down-regulating NF-kappaB signaling pathway. Biochem. Biophys. Res. Commun. 2012, 419, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Herold, S.; Gabrielli, N.M.; Vadasz, I. Novel concepts of acute lung injury and alveolar-capillary barrier dysfunction. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L665–L681. [Google Scholar] [CrossRef] [PubMed]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol. Med. 2011, 17, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Sun, Y.; Shen, Y.; Li, F.; Song, X.; Zhou, E.; Zhao, F.; Liu, Z.; Fu, Y.; Guo, M.; et al. Shikonin exerts anti-inflammatory effects in a murine model of lipopolysaccharide-induced acute lung injury by inhibiting the nuclear factor-kappaB signaling pathway. Int. Immunopharmacol. 2013, 16, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Piao, T.; Wang, M.; Zhang, J.; Jiang, J.; Wang, X.; Liu, H. Protective effect of catalpol on lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2014, 23, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Joh, E.H.; Gu, W.; Kim, D.H. Echinocystic acid ameliorates lung inflammation in mice and alveolar macrophages by inhibiting the binding of LPS to TLR4 in NF-kappaB and MAPK pathways. Biochem. Pharmacol. 2012, 84, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Moochhala, S. Role of inflammatory mediators in the pathophysiology of acute respiratory distress syndrome. J. Pathol. 2004, 202, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Hoidal, J.R.; Mukherjee, T.K. Role of TNFalpha in pulmonary pathophysiology. Respir. Res. 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- Grailer, J.J.; Canning, B.A.; Kalbitz, M.; Haggadone, M.D.; Dhond, R.M.; Andjelkovic, A.V.; Zetoune, F.S.; Ward, P.A. Critical role for the NLRP3 inflammasome during acute lung injury. J. Immunol. 2014, 192, 5974–5983. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Gerondakis, S. Coordinating TLR-activated signaling pathways in cells of the immune system. Immunol. Cell Biol. 2007, 85, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.B.; Reddanna, P. Chebulagic acid (CA) attenuates LPS-induced inflammation by suppressing NF-kappaB and MAPK activation in RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 381, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, X.; Liu, T.; Guan, M.; Feng, X.; Dong, W.; Chu, X.; Liu, J.; Tian, X.; Ci, X.; et al. Kaempferol regulates MAPKs and NF-kappaB signaling pathways to attenuate LPS-induced acute lung injury in mice. Int. Immunopharmacol. 2012, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.M.; Tan, L.; Wang, Z.R.; Liu, S.X.; Wang, Y.L.; Liang, S.Y.; Zhong, J.B.; Lin, H.S. Preventive and therapeutic effects of Danhong injection on lipopolysaccharide induced acute lung injury in mice. J. Ethnopharmacol. 2013, 149, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.C.; Moore, J.N.; Hurley, D.J.; Vandenplas, M.L.; Linden, J.; Cao, Z.; Murray, T.F. Adenosine A2A receptor agonists inhibit lipopolysaccharide-induced production of tumor necrosis factor-alpha by equine monocytes. Vet. Immunol. Immunopathol. 2008, 121, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compound HJB-1 are available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Liu, N.; Zhang, Y.-X.; Cao, J.; Wu, D.; Peng, Q.; Wang, H.-B.; Sun, W.-C. The Protective Effects of HJB-1, a Derivative of 17-Hydroxy-Jolkinolide B, on LPS-Induced Acute Distress Respiratory Syndrome Mice. Molecules 2016, 21, 77. https://doi.org/10.3390/molecules21010077
Xu X, Liu N, Zhang Y-X, Cao J, Wu D, Peng Q, Wang H-B, Sun W-C. The Protective Effects of HJB-1, a Derivative of 17-Hydroxy-Jolkinolide B, on LPS-Induced Acute Distress Respiratory Syndrome Mice. Molecules. 2016; 21(1):77. https://doi.org/10.3390/molecules21010077
Chicago/Turabian StyleXu, Xiaohan, Ning Liu, Yu-Xin Zhang, Jinjin Cao, Donglin Wu, Qisheng Peng, Hong-Bing Wang, and Wan-Chun Sun. 2016. "The Protective Effects of HJB-1, a Derivative of 17-Hydroxy-Jolkinolide B, on LPS-Induced Acute Distress Respiratory Syndrome Mice" Molecules 21, no. 1: 77. https://doi.org/10.3390/molecules21010077
APA StyleXu, X., Liu, N., Zhang, Y.-X., Cao, J., Wu, D., Peng, Q., Wang, H.-B., & Sun, W.-C. (2016). The Protective Effects of HJB-1, a Derivative of 17-Hydroxy-Jolkinolide B, on LPS-Induced Acute Distress Respiratory Syndrome Mice. Molecules, 21(1), 77. https://doi.org/10.3390/molecules21010077