
Glycyrrhizic Acid Reduces Heart Rate and Blood Pressure by a Dual Mechanism


Abstract
:1. Introduction
2. Results
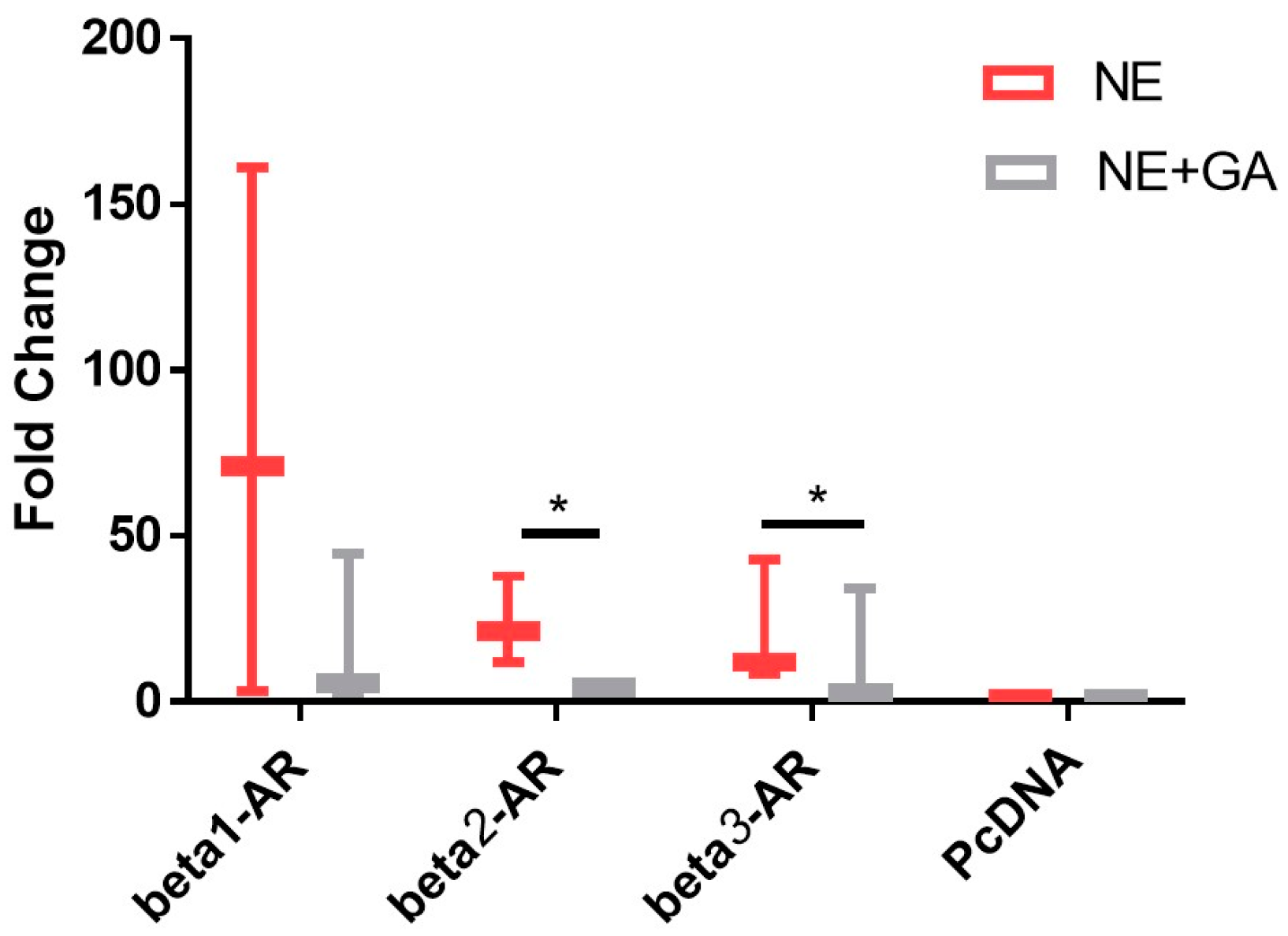
2.1. In Vitro cAMP Assay
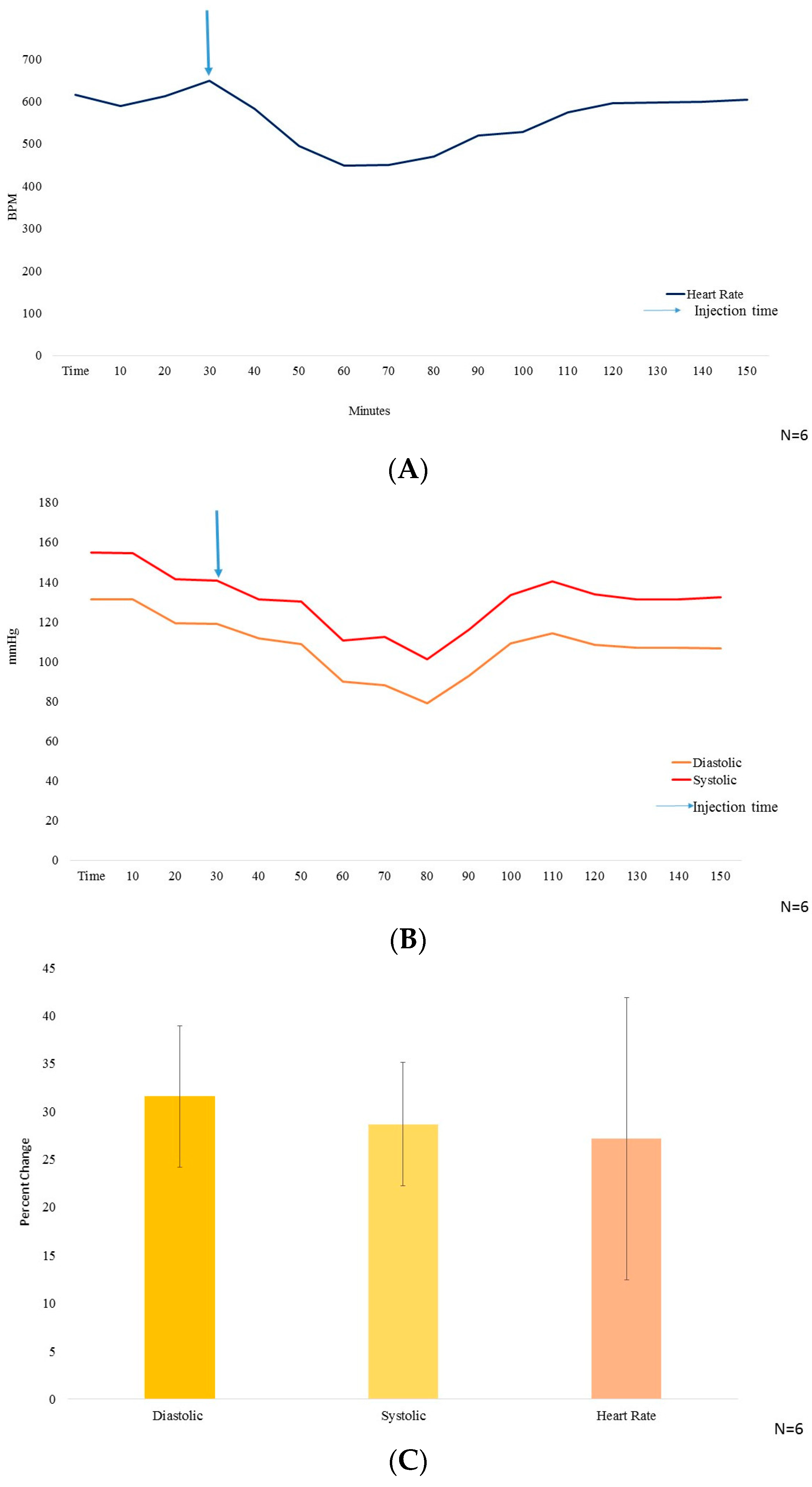
2.2. Acute Effect of GA on Heart Rate & Blood Pressure
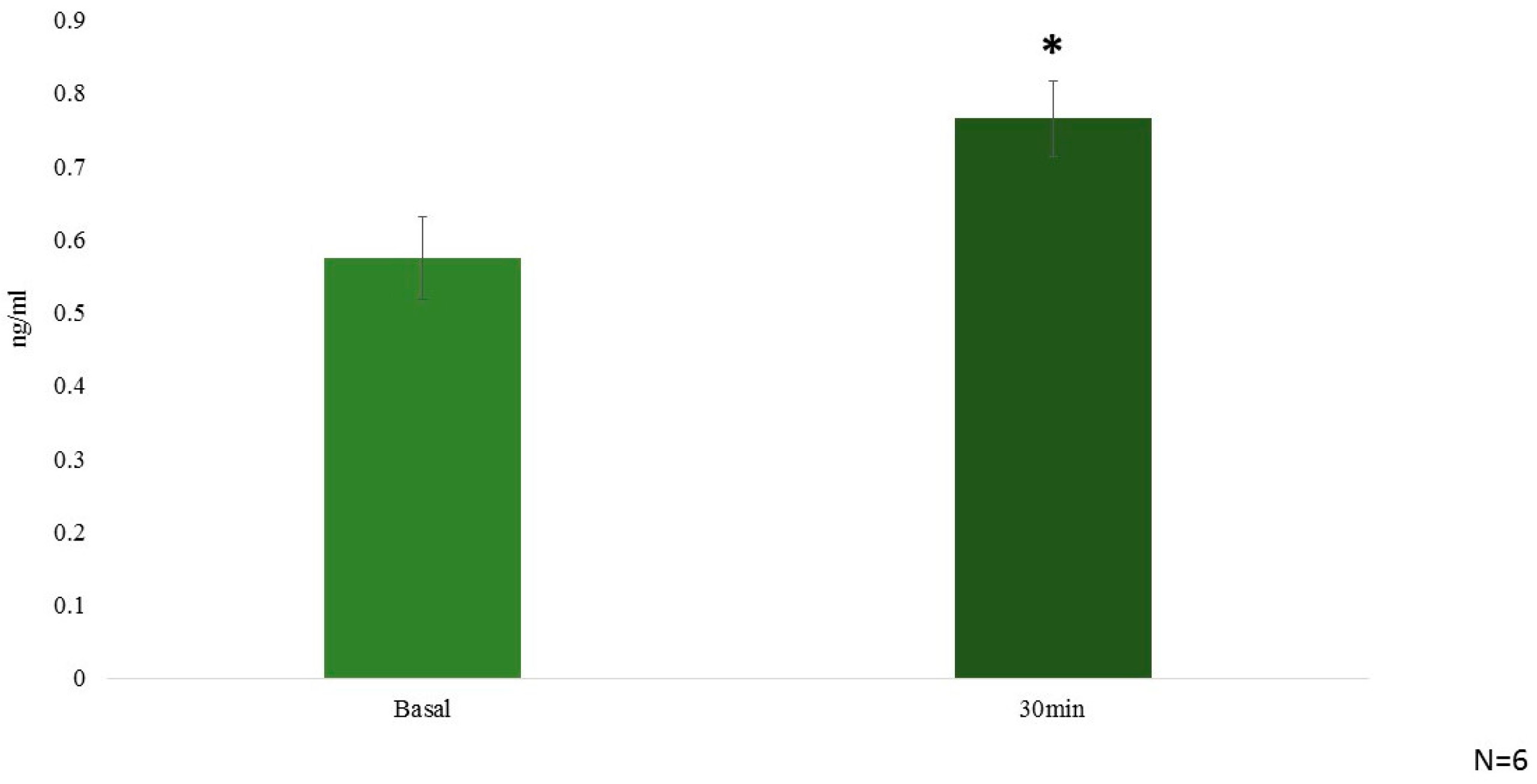
2.3. In Vivo Hormonal Effect
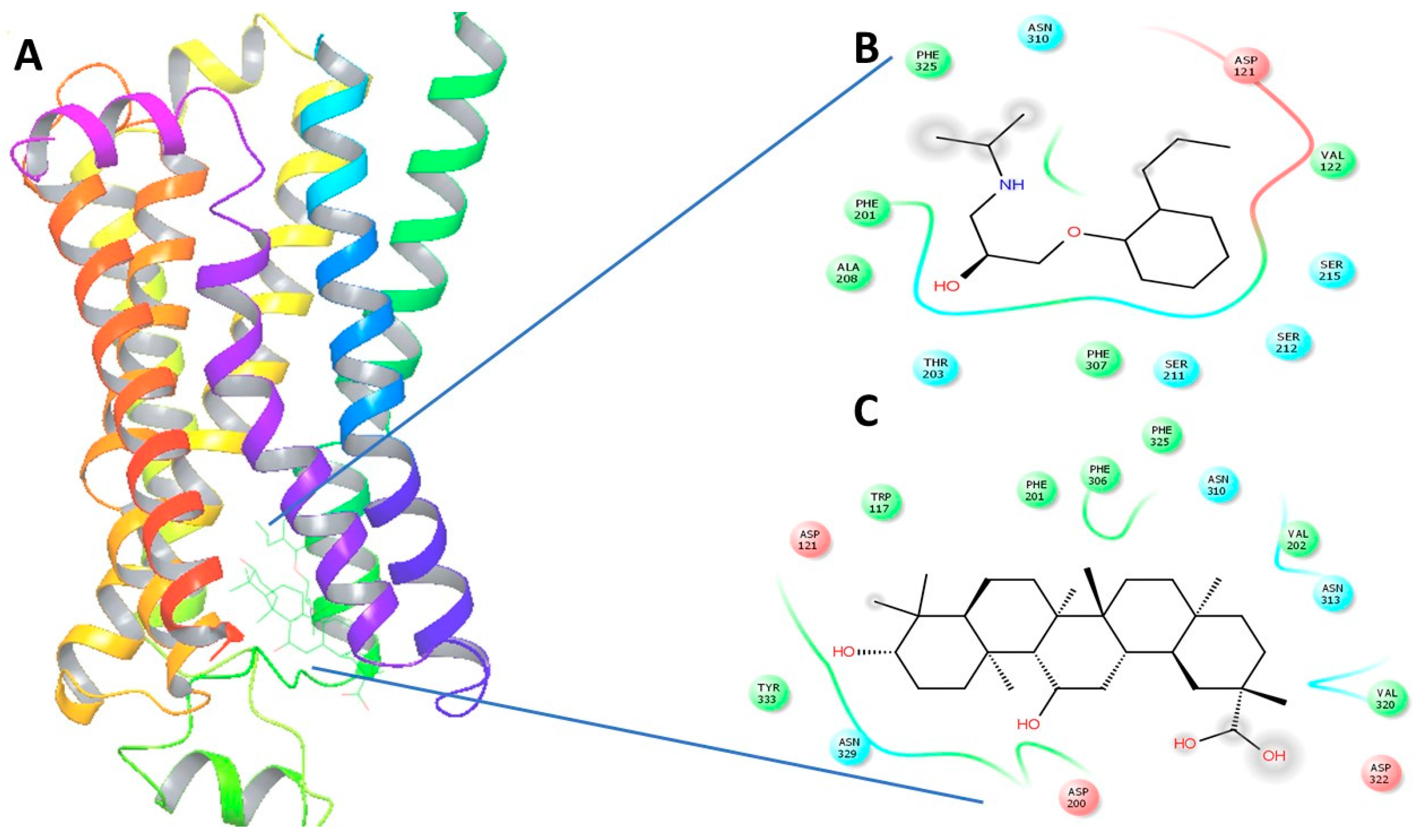
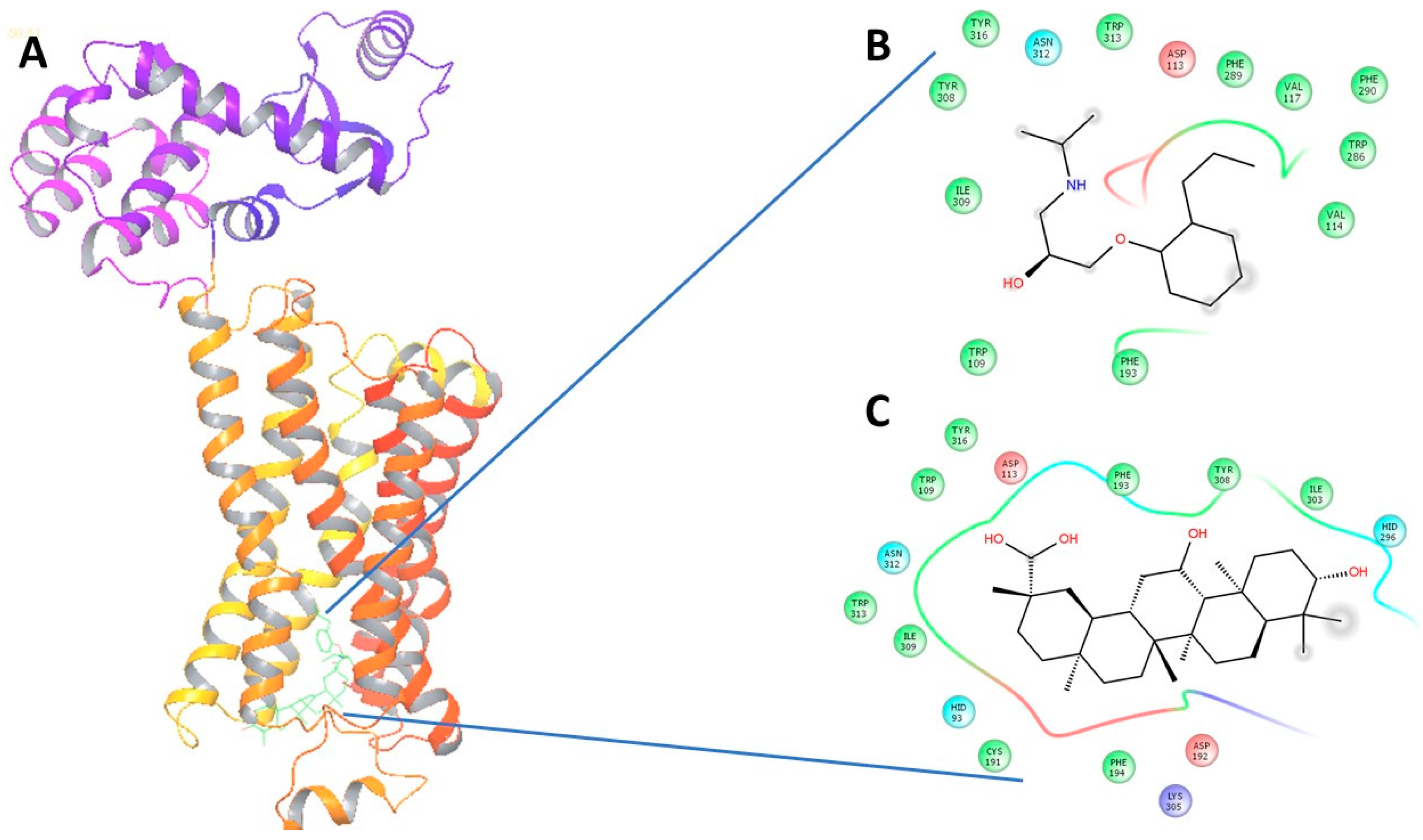
2.4. In Silico Virtual Docking
3. Discussion
4. Experimental Section
4.1. In Vitro Transfected Cell and Effect of GA on cAMP Levels
4.2. Animal Grouping (Randomization)
4.3. Serum Hormonal Level Study
4.4. DSI Implantation
4.5. Heart Rate and Blood Pressure Study
4.6. Virtual Docking
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Madamanchi, A. Beta-adrenergic receptor signaling in cardiac function and heart failure. McGill J. Med. 2007, 10, 99–104. [Google Scholar] [PubMed]
- Insel, P.A. Adrenergic receptors—Evolving concepts and clinical implications. N. Engl. J. Med. 1996, 334, 580–585. [Google Scholar] [PubMed]
- Zipes, D.P. Sympathetic stimulation and arrhythmias. N. Engl. J. Med. 1991, 325, 656–657. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.P. Beta-adrenergic signaling in the heart: Dual coupling of the beta2-adrenergic receptor to g(s) and g(i) proteins. Sci. STKE 2001, 2001, re15. [Google Scholar] [PubMed]
- Taira, C.A.; Carranza, A.; Mayer, M.; Di Verniero, C.; Opezzo, J.A.; Hocht, C. Therapeutic implications of beta-adrenergic receptor pharmacodynamic properties. Curr. Clin. Pharmacol. 2008, 3, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Aidietis, A.; Laucevicius, A.; Marinskis, G. Hypertension and cardiac arrhythmias. Curr. Pharm. Des. 2007, 13, 2545–2555. [Google Scholar] [CrossRef] [PubMed]
- Frielle, T.; Collins, S.; Daniel, K.W.; Caron, M.G.; Lefkowitz, R.J.; Kobilka, B.K. Cloning of the cdna for the human beta 1-adrenergic receptor. Proc. Natl. Acad. Sci. USA 1987, 84, 7920–7924. [Google Scholar] [CrossRef] [PubMed]
- Granneman, J.G. The putative beta4-adrenergic receptor is a novel state of the beta1-adrenergic receptor. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E199–E202. [Google Scholar] [PubMed]
- Milano, C.A.; Allen, L.F.; Rockman, H.A.; Dolber, P.C.; McMinn, T.R.; Chien, K.R.; Johnson, T.D.; Bond, R.A.; Lefkowitz, R.J. Enhanced myocardial function in transgenic mice overexpressing the beta 2-adrenergic receptor. Science 1994, 264, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Chruscinski, A.J.; Rohrer, D.K.; Schauble, E.; Desai, K.H.; Bernstein, D.; Kobilka, B.K. Targeted disruption of the beta2 adrenergic receptor gene. J. Biol. Chem. 1999, 274, 16694–16700. [Google Scholar] [CrossRef] [PubMed]
- Freedman, N.J.; Lefkowitz, R.J. Anti-beta(1)-adrenergic receptor antibodies and heart failure: Causation, not just correlation. J. Clin. Investig. 2004, 113, 1379–1382. [Google Scholar] [CrossRef] [PubMed]
- Goldie, R.G.; Papadimitriou, J.M.; Paterson, J.W.; Rigby, P.J.; Spina, D. Autoradiographic localization of beta-adrenoceptors in pig lung using [125i]-iodocyanopindolol. Br. J. Pharmacol. 1986, 88, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Pourageaud, F.; Leblais, V.; Bellance, N.; Marthan, R.; Muller, B. Role of beta2-adrenoceptors (beta-ar), but not beta1-, beta3-ar and endothelial nitric oxide, in beta-ar-mediated relaxation of rat intrapulmonary artery. Naunyn Schmiedebergs Arch. Pharmacol. 2005, 372, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Brodde, O.E.; Michel, M.C. Adrenergic and muscarinic receptors in the human heart. Pharmacol. Rev. 1999, 51, 651–690. [Google Scholar] [PubMed]
- Bristow, M.R.; Ginsburg, R.; Umans, V.; Fowler, M.; Minobe, W.; Rasmussen, R.; Zera, P.; Menlove, R.; Shah, P.; Jamieson, S.; et al. Beta 1- and beta 2-adrenergic-receptor subpopulations in nonfailing and failing human ventricular myocardium: Coupling of both receptor subtypes to muscle contraction and selective beta 1-receptor down-regulation in heart failure. Circ. Res. 1986, 59, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Fowler, M.B.; Laser, J.A.; Hopkins, G.L.; Minobe, W.; Bristow, M.R. Assessment of the beta-adrenergic receptor pathway in the intact failing human heart: Progressive receptor down-regulation and subsensitivity to agonist response. Circulation 1986, 74, 1290–1302. [Google Scholar] [CrossRef] [PubMed]
- Altschuld, R.A.; Starling, R.C.; Hamlin, R.L.; Billman, G.E.; Hensley, J.; Castillo, L.; Fertel, R.H.; Hohl, C.M.; Robitaille, P.M.; Jones, L.R.; et al. Response of failing canine and human heart cells to beta 2-adrenergic stimulation. Circulation 1995, 92, 1612–1618. [Google Scholar] [CrossRef] [PubMed]
- Freemantle, N.; Cleland, J.; Young, P.; Mason, J.; Harrison, J. Beta blockade after myocardial infarction: Systematic review and meta regression analysis. BMJ 1999, 318, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Ploeger, B.; Mensinga, T.; Sips, A.; Seinen, W.; Meulenbelt, J.; DeJongh, J. The pharmacokinetics of glycyrrhizic acid evaluated by physiologically based pharmacokinetic modeling. Drug Metab. Rev. 2001, 33, 125–147. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.G. Inhibitory actions of glycyrrhizic acid on arylamine N-acetyltransferase activity in strains of helicobacter pylori from peptic ulcer patients. Drug Chem. Toxicol. 1998, 21, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Kao, T.C.; Shyu, M.H.; Yen, G.C. Glycyrrhizic acid and 18beta-glycyrrhetinic acid inhibit inflammation via pi3k/akt/gsk3beta signaling and glucocorticoid receptor activation. J. Agric. Food Chem. 2010, 58, 8623–8629. [Google Scholar] [CrossRef] [PubMed]
- Polyakov, N.E.; Leshina, T.V.; Salakhutdinov, N.F.; Konovalova, T.A.; Kispert, L.D. Antioxidant and redox properties of supramolecular complexes of carotenoids with beta-glycyrrhizic acid. Free Radic. Biol. Med. 2006, 40, 1804–1809. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yin, J.; Sui, X.; Li, G.; Ren, X. Comparative analysis of the effect of glycyrrhizin diammonium and lithium chloride on infectious bronchitis virus infection in vitro. Avian Pathol. 2009, 38, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Yoke Yin, C.; So Ha, T.; Abdul Kadir, K. Effects of glycyrrhizic acid on peroxisome proliferator-activated receptor gamma (ppargamma), lipoprotein lipase (lpl), serum lipid and homa-ir in rats. PPAR Res. 2010, 2010, 530265. [Google Scholar] [CrossRef] [PubMed]
- Chia, Y.Y.; Liong, S.Y.; Ton, S.H.; Kadir, K.B. Amelioration of glucose homeostasis by glycyrrhizic acid through gluconeogenesis rate-limiting enzymes. Eur. J. Pharmacol. 2012, 677, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Cherng, J.M.; Tsai, K.D.; Yu, Y.W.; Lin, J.C. Molecular mechanisms underlying chemopreventive activities of glycyrrhizic acid against uvb-radiation-induced carcinogenesis in skh-1 hairless mouse epidermis. Radiat. Res. 2011, 176, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Raphael, T.J.; Kuttan, G. Effect of naturally occurring triterpenoids ursolic acid and glycyrrhizic acid on the cell-mediated immune responses of metastatic tumor-bearing animals. Immunopharmacol. Immunotoxicol. 2008, 30, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.T.; Chan, C.; Ho, J.W. Inhibition of glycyrrhizic acid on aflatoxin b1-induced cytotoxicity in hepatoma cells. Toxicology 2003, 188, 211–217. [Google Scholar] [CrossRef]
- Yoh, T.; Nakashima, T.; Sumida, Y.; Kakisaka, Y.; Nakajima, Y.; Ishikawa, H.; Sakamoto, Y.; Okanoue, T.; Mitsuyoshi, H. Effects of glycyrrhizin on glucocorticoid signaling pathway in hepatocytes. Dig. Dis. Sci. 2002, 47, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Senthil, V. Structure, Activity and Relationship Studies of Peptide and Non-Peptide Analogs with Secretin Receptor: In Search of Agonist and/or Antagonist. Ph.D. Thesis, University of Hong Kong, Hong Kong, China, 2014. [Google Scholar]
- Pompei, R.; Flore, O.; Marccialis, M.A.; Pani, A.; Loddo, B. Glycyrrhizic acid inhibits virus growth and inactivates virus particles. Nature 1979, 281, 689–690. [Google Scholar] [CrossRef] [PubMed]
- Kailash Singh, A.M.Z.; Revathi Sekar, A.A.A.; Jamaan, A.; Billy, K.C.C. Possible role of secretin in anti-ulcer activity of glycyrrhizic acid. 2016; manuscript in preparation. [Google Scholar]
- Watanabe, S.I.; Chey, W.Y.; Lee, K.Y.; Chang, T.M. Release of secretin by licorice extract in dogs. Pancreas 1986, 1, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Hickie, J.B. Alprenolol (“aptin”) in angina pectoris. A double-blind multicentre trial. Med. J. Aust. 1970, 2, 268–272. [Google Scholar] [PubMed]
- Santulli, G.; Iaccarino, G. Adrenergic signaling in heart failure and cardiovascular aging. Maturitas 2016. [Google Scholar] [CrossRef] [PubMed]
- Ciccarelli, M.; Santulli, G.; Pascale, V.; Trimarco, B.; Iaccarino, G. Adrenergic receptors and metabolism: Role in development of cardiovascular disease. Front. Physiol. 2013, 4, 265. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Ciccarelli, M.; Trimarco, B.; Iaccarino, G. Physical activity ameliorates cardiovascular health in elderly subjects: The functional role of the beta adrenergic system. Front. Physiol. 2013, 4, 209. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Lombardi, A.; Sorriento, D.; Anastasio, A.; Del Giudice, C.; Formisano, P.; Beguinot, F.; Trimarco, B.; Miele, C.; Iaccarino, G. Age-related impairment in insulin release: The essential role of beta(2)-adrenergic receptor. Diabetes 2012, 61, 692–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, C.M.; Nelson, R.; Deegan, R.; He, H.; Wood, M.; Wood, A.J. Forearm beta adrenergic receptor-mediated vasodilation is impaired, without alteration of forearm norepinephrine spillover, in borderline hypertension. J. Clin. Investig. 1995, 96, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Izzo, R.; Cipolletta, E.; Ciccarelli, M.; Campanile, A.; Santulli, G.; Palumbo, G.; Vasta, A.; Formisano, S.; Trimarco, B.; Iaccarino, G. Enhanced grk2 expression and desensitization of betaar vasodilatation in hypertensive patients. Clin. Transl. Sci. 2008, 1, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Perino, A.; Ghigo, A.; Ferrero, E.; Morello, F.; Santulli, G.; Baillie, G.S.; Damilano, F.; Dunlop, A.J.; Pawson, C.; Walser, R.; et al. Integrating cardiac pip3 and camp signaling through a pka anchoring function of p110gamma. Mol. Cell 2011, 42, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Dosa, P.I.; Amin, E.A. Tactical approaches to interconverting gpcr agonists and antagonists. J. Med. Chem. 2016, 59, 810–840. [Google Scholar] [CrossRef] [PubMed]
- Ram, C.V. Beta-blockers in hypertension. Am. J. Cardiol. 2010, 106, 1819–1825. [Google Scholar] [CrossRef] [PubMed]
- Antov, G.; Khalkova, Z.; Mikhailova, A.; Zaikov, K.; Burkova, T. The toxicological characteristics of ammonium glycyrrhizinate (glycyram). A study of its acute and subacute toxicity. Eksp. Klin. Farmakol. 1997, 60, 65–67. [Google Scholar] [PubMed]
- Bengtsson, K.; Orho-Melander, M.; Melander, O.; Lindblad, U.; Ranstam, J.; Rastam, L.; Groop, L. Beta(2)-adrenergic receptor gene variation and hypertension in subjects with type 2 diabetes. Hypertension 2001, 37, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Billman, G.E.; Castillo, L.C.; Hensley, J.; Hohl, C.M.; Altschuld, R.A. Beta2-adrenergic receptor antagonists protect against ventricular fibrillation: In vivo and in vitro evidence for enhanced sensitivity to beta2-adrenergic stimulation in animals susceptible to sudden death. Circulation 1997, 96, 1914–1922. [Google Scholar] [CrossRef] [PubMed]
- Parisella, M.L.; Angelone, T.; Gattuso, A.; Cerra, M.C.; Pellegrino, D. Glycyrrhizin and glycyrrhetinic acid directly modulate rat cardiac performance. J. Nutr. Biochem. 2012, 23, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Iaccarino, G. Pinpointing beta adrenergic receptor in ageing pathophysiology: Victim or executioner? Evidence from crime scenes. Immun. Ageing 2013, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Ciccarelli, M.; Cipolletta, E.; Santulli, G.; Campanile, A.; Pumiglia, K.; Cervero, P.; Pastore, L.; Astone, D.; Trimarco, B.; Iaccarino, G. Endothelial beta2 adrenergic signaling to akt: Role of gi and src. Cell. Signal. 2007, 19, 1949–1955. [Google Scholar] [CrossRef] [PubMed]
- Van der Westhuizen, E.T.; Breton, B.; Christopoulos, A.; Bouvier, M. Quantification of ligand bias for clinically relevant beta2-adrenergic receptor ligands: Implications for drug taxonomy. Mol. Pharmacol. 2014, 85, 492–509. [Google Scholar] [CrossRef] [PubMed]
- On, J.S.; Duan, C.; Chow, B.K.; Lee, L.T. Functional pairing of class b1 ligand-gpcr in cephalochordate provides evidence of the origin of pth and pacap/glucagon receptor family. Mol. Biol. Evol. 2015, 32, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Senthil, V.; Arokiaraj, A.W.; Leprince, J.; Lefranc, B.; Vaudry, D.; Allam, A.A.; Ajarem, J.; Chow, B.K. Structure-activity relationship studies of n- and c-terminally modified secretin analogs for the human secretin receptor. PLoS ONE 2016, 11, e0149359. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Leslie, A.G.; Warne, T.; Tate, C.G. Ligand occupancy in crystal structure of beta1-adrenergic g protein-coupled receptor. Nat. Struct. Mol. Biol. 2015, 22, 941–942. [Google Scholar] [CrossRef] [PubMed]
- Wacker, D.; Fenalti, G.; Brown, M.A.; Katritch, V.; Abagyan, R.; Cherezov, V.; Stevens, R.C. Conserved binding mode of human beta2 adrenergic receptor inverse agonists and antagonist revealed by X-ray crystallography. J. Am. Chem. Soc. 2010, 132, 11443–11445. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. Patchdock and symmdock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef] [PubMed]
- Andrusier, N.; Nussinov, R.; Wolfson, H.J. Firedock: Fast interaction refinement in molecular docking. Proteins 2007, 69, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Beard, H.; Cholleti, A.; Pearlman, D.; Sherman, W.; Loving, K.A. Applying physics-based scoring to calculate free energies of binding for single amino acid mutations in protein-protein complexes. PLoS ONE 2013, 8, e82849. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | β1-AR | β2-AR |
|---|---|---|
| PDB ID | 5F8U | 3NYA |
| NE | −28.08 | −33.36 |
| Alperenolol | −38.14 | −40.61 |
| GA | −56.27 | −59.51 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, K.; Zaw, A.M.; Sekar, R.; Palak, A.; Allam, A.A.; Ajarem, J.; Chow, B.K.C. Glycyrrhizic Acid Reduces Heart Rate and Blood Pressure by a Dual Mechanism. Molecules 2016, 21, 1291. https://doi.org/10.3390/molecules21101291
Singh K, Zaw AM, Sekar R, Palak A, Allam AA, Ajarem J, Chow BKC. Glycyrrhizic Acid Reduces Heart Rate and Blood Pressure by a Dual Mechanism. Molecules. 2016; 21(10):1291. https://doi.org/10.3390/molecules21101291
Chicago/Turabian StyleSingh, Kailash, Aung Moe Zaw, Revathi Sekar, Ahuja Palak, Ahmed A. Allam, Jamaan Ajarem, and Billy K. C. Chow. 2016. "Glycyrrhizic Acid Reduces Heart Rate and Blood Pressure by a Dual Mechanism" Molecules 21, no. 10: 1291. https://doi.org/10.3390/molecules21101291
APA StyleSingh, K., Zaw, A. M., Sekar, R., Palak, A., Allam, A. A., Ajarem, J., & Chow, B. K. C. (2016). Glycyrrhizic Acid Reduces Heart Rate and Blood Pressure by a Dual Mechanism. Molecules, 21(10), 1291. https://doi.org/10.3390/molecules21101291