
Transcriptome Profile Reveals that Pu-Erh Tea Represses the Expression of Vitellogenin Family to Reduce Fat Accumulation in Caenorhabditis elegans


Abstract
:1. Introduction
2. Results and Discussion
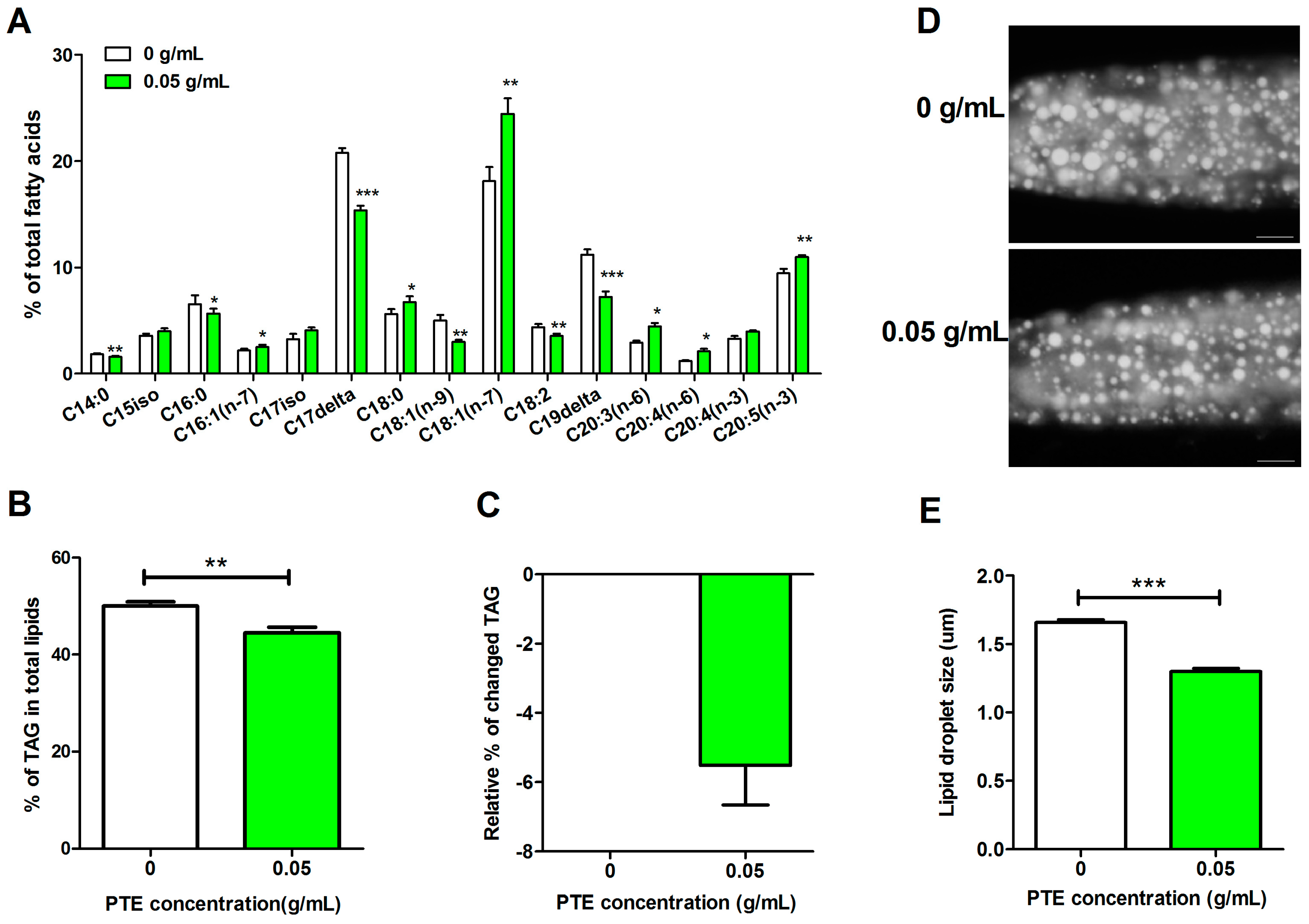
2.1. PTE Reduces Fat Accumulation and Modifies Fatty Acid Compositions
2.2. Transcriptome Results Analysis
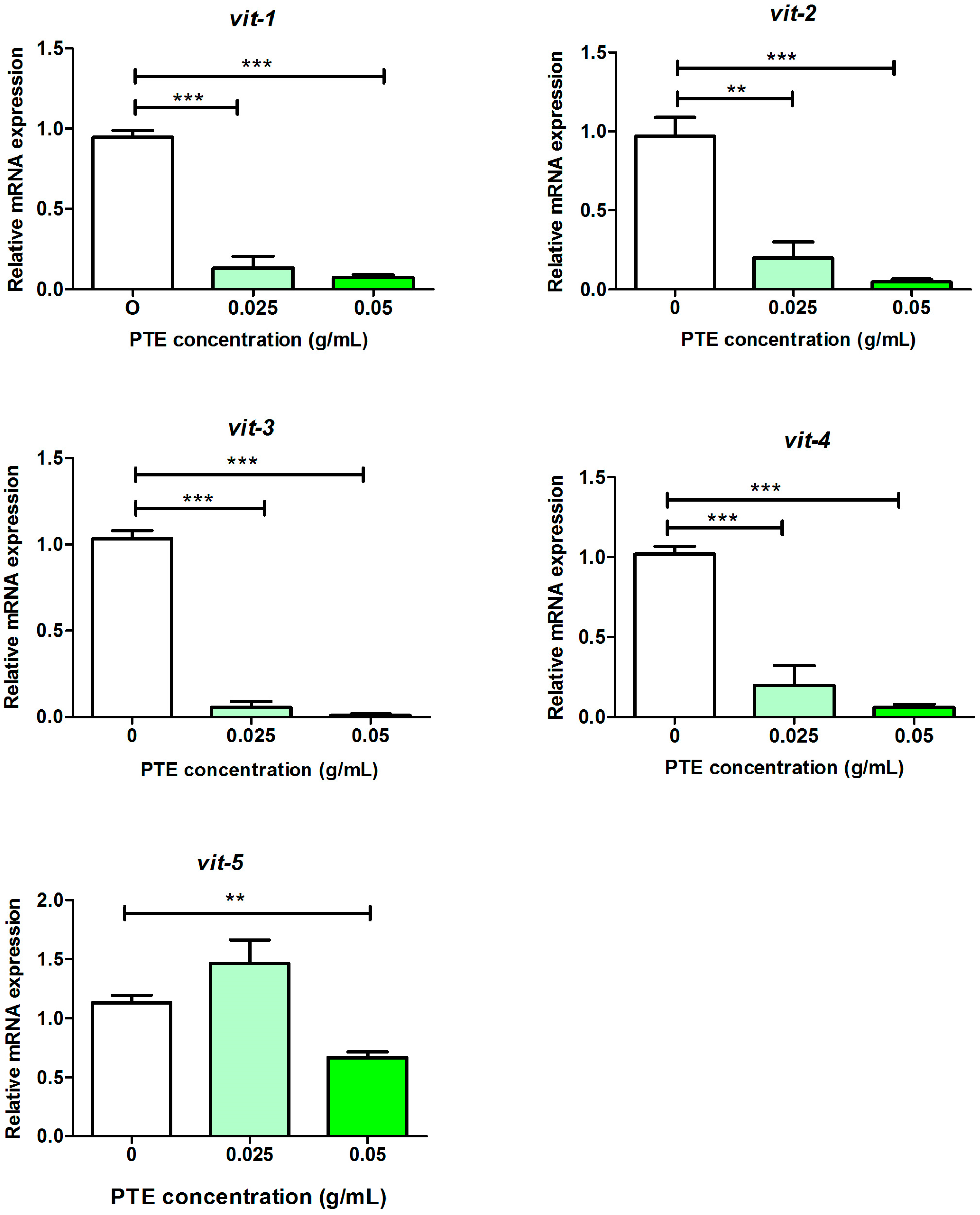
2.3. Validation of RNA-Seq Results by Quantitative PCR (qPCR)
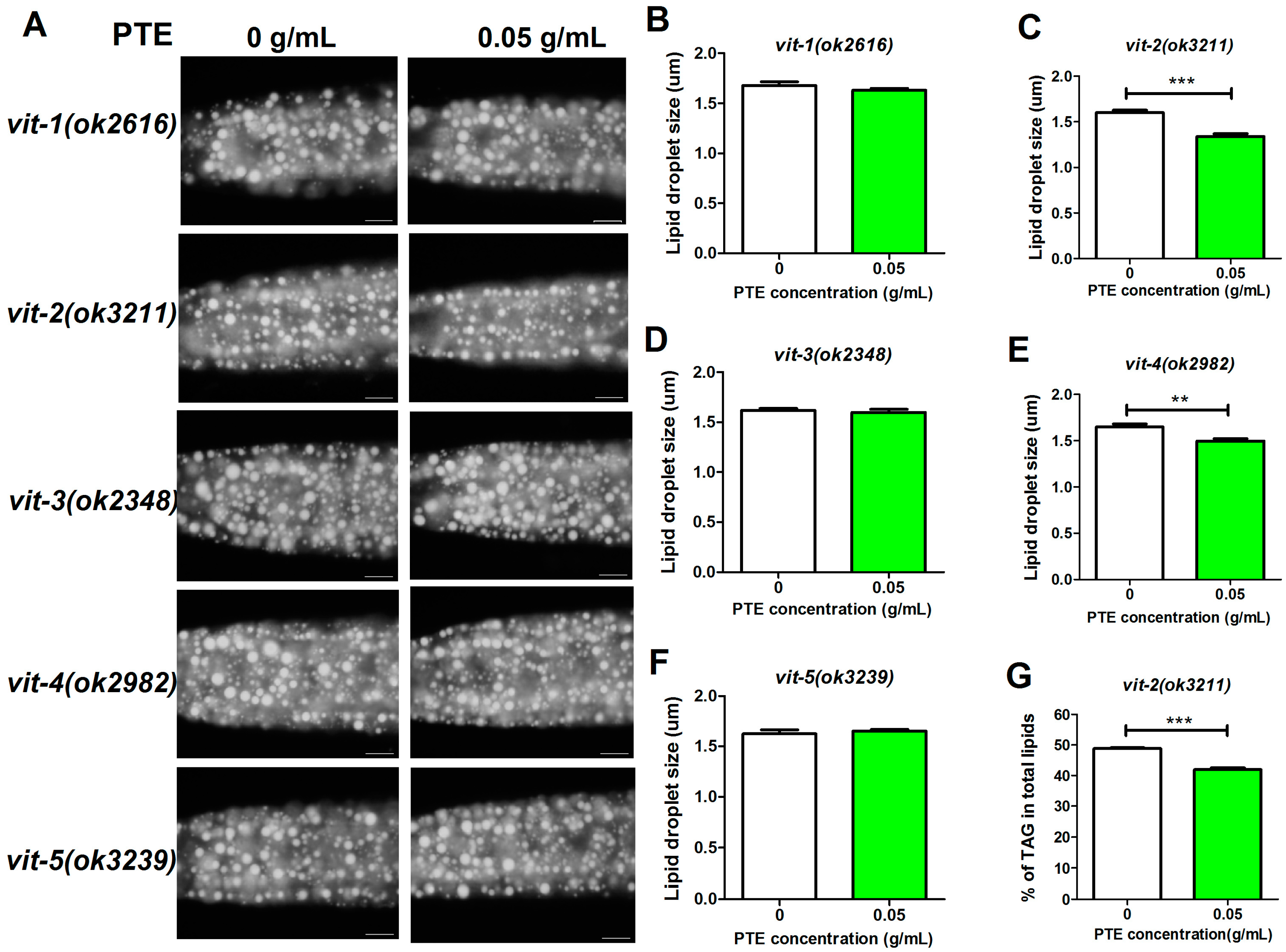
2.4. Mutants of Vitellogenin Genes are Resistant to PTE Reduced Fat Accumulation
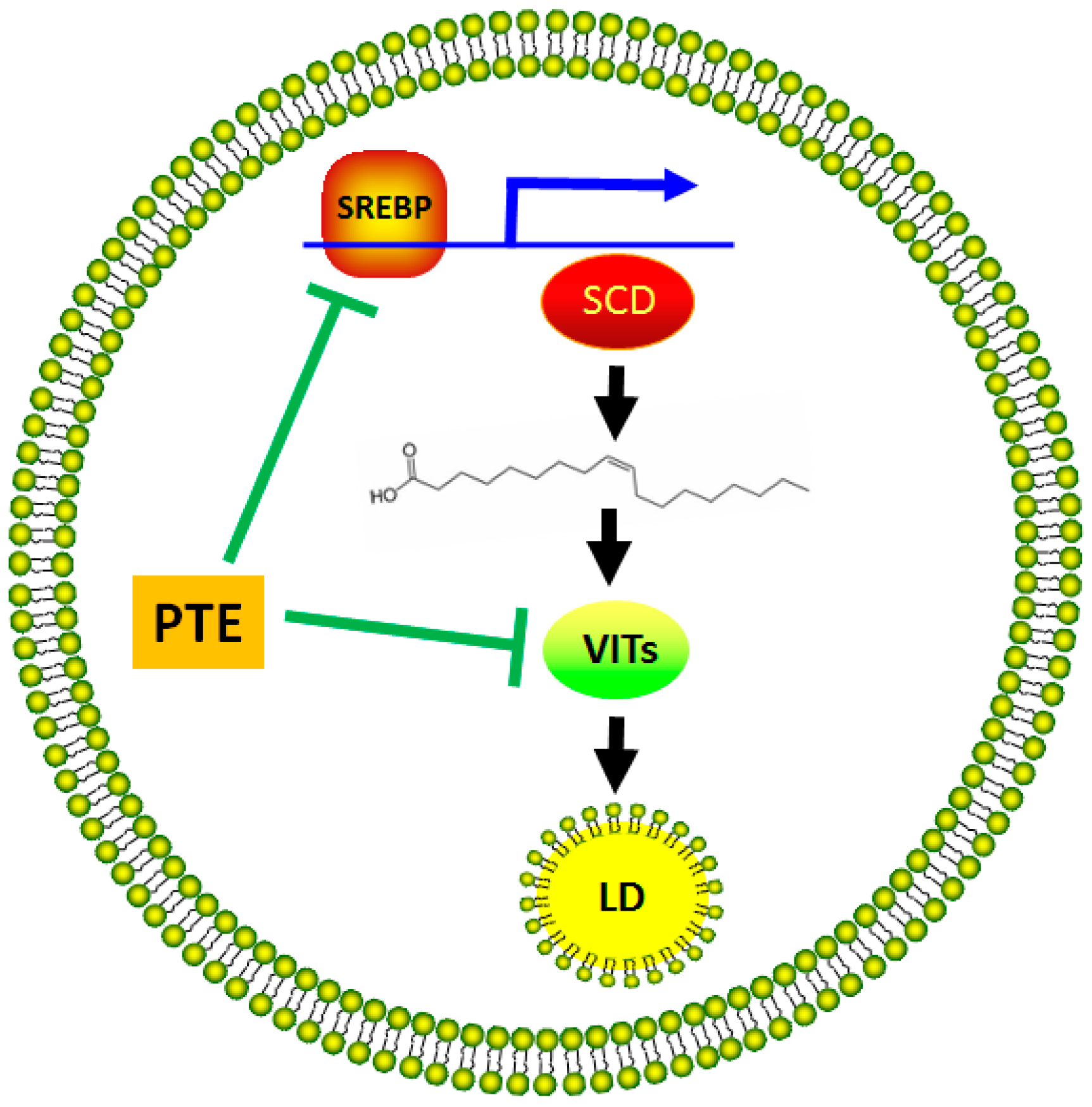
2.5. Discussion
3. Materials and Methods
3.1. Production of the Aqueous Extract of Pu-Erh Tea
3.2. Culture Conditions and Worm Strains
3.3. Samples Collection and Sequencing
3.4. Nile Red Staining of Fixation
3.5. Lipid Extraction and Analysis
3.6. RNA Isolation and qPCR Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of interest
References
- Osborn, O.; Olefsky, J.M. The cellular and signaling networks linking the immune system and metabolism in disease. Nat. Med. 2012, 18, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Hossain, P.; Kawar, B.; El Nahas, M. Obesity and diabetes in the developing world—A growing challenge. New Eng. J. Med. 2007, 356, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Raitakari, M. Weight reduction with very-low-caloric diet and endothelial function in overweight adults: Role of plasma glucose. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Everson, E.; Boles, M.; Fink, K.; Topol, R.; Fenaughty, A. Estimating the prevalence of childhood obesity in alaska using partial, nonrandom measurement data. Prev. Chron. Dis. 2016, 13, E40. [Google Scholar] [CrossRef] [PubMed]
- Gemelli, I.F.B.; Farias, E.d.S.; Souza, O.F. Age at menarche and its association with excess weight and body fat percentage in girls in the southwestern region of the brazilian amazon. J. Pediatr. Adolesc. Gynecol. 2016, 29, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Olds, T.S.; Tomkinson, G.R.; Ferrar, K.E.; Maher, C.A. Trends in the prevalence of childhood overweight and obesity in australia between 1985 and 2008. Int. J. Obes. 2009, 34, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Dietrich, E.S.; Venetz, W.P. Trends and stabilization up to 2022 in overweight and obesity in switzerland, comparison to France, UK, US and Australia. Int. J. Environ. Res. Publ. Health 2010, 7, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, M.; Bentham, J.; Stevens, G.A.; Zhou, B.; Danaei, G.; Lu, Y.; Bixby, H.; Cowan, M.J.; Riley, L.M.; Hajifathalian, K.; et al. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar]
- Padwal, R.S.; Majumdar, S.R. Drug treatments for obesity: Orlistat, sibutramine, and rimonabant. Lancet 2007, 369, 71–77. [Google Scholar] [CrossRef]
- Cooke, D.; Bloom, S. The obesity pipeline: Current strategies in the development of anti-obesity drugs. Nat. Rev. Drug Discov. 2006, 5, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Balaji, M.; Ganjayi, M.S.; Hanuma Kumar, G.E.; Parim, B.N.; Mopuri, R.; Dasari, S. A review on possible therapeutic targets to contain obesity: The role of phytochemicals. Obes. Res. Clin. Pract. 2016, 10, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Chen, S.; Chen, H.; Wang, Y.; Wang, Y.; Hochstetter, D.; Xu, P. Studies on the bioactivity of aqueous extract of pu-erh tea and its fractions: In vitro antioxidant activity and α-glycosidase inhibitory property, and their effect on postprandial hyperglycemia in diabetic mice. Food Chem. Toxicol. 2013, 53, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Lin, J.-K. Pu-erh tea, green tea, and black tea suppresses hyperlipidemia, hyperleptinemia and fatty acid synthase through activating ampk in rats fed a high-fructose diet. Food Funct. 2012, 3, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Shao, W.; Xiao, R.; Xu, K.; Ma, Z.; Johnstone, B.H.; Du, Y. Pu-erh tea aqueous extracts lower atherosclerotic risk factors in a rat hyperlipidemia model. Experim. Gerontol. 2009, 44, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Duh, P.D.; Yen, G.C.; Yen, W.J.; Wang, B.S.; Chang, L.W. Effects of pu-erh tea on oxidative damage and nitric oxide scavenging. J. Agric. Food Chem. 2004, 52, 8169–8176. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jia, S.; Tang, W.; Sheng, J.; Luo, Y. Pu-erh tea inhibits tumor cell growth by down-regulating mutant p53. Int. J. Mol. Sci. 2011, 12, 7581–7593. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, Y.; Yoda, M.; Sakakibara, H.; Matsunaga, K.; Masuda, S. Pu-erh tea suppresses diet-induced body fat accumulation in c57bl/6j mice by down-regulating srebp-1c and related molecules. Biosci. Biotechnol. Biochem. 2014, 77, 1455–1460. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.-H.; Gu, D.-H.; Lin, Q.-Y.; Xu, Z.-Q.; Huang, Q.-C.; Rao, H.; Liu, E.-W.; Jia, J.-J.; Ge, C.-R. Effect of pu-erh tea on body fat and lipid profiles in rats with diet-induced obesity. Phytother. Res. 2010, 25, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Kuo, K.L.; Weng, M.S.; Chiang, C.T.; Tsai, Y.J.; Lin-Shiau, S.Y.; Lin, J.K. Comparative studies on the hypolipidemic and growth suppressive effects of oolong, black, pu-erh, and green tea leaves in rats. J. Agric. Food Chem. 2005, 53, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ding, Y.; Zou, X.; Jiang, X.; Wu, J.; Zhang, Y.; Chen, D.; Liang, B. Pu-erh tea down-regulates sterol regulatory element-binding protein and stearyol-coa desaturase to reduce fat storage in caenorhaditis elegans. PLoS ONE 2015, 10, e0113815. [Google Scholar]
- Cai, X.; Fang, C.; Hayashi, S.; Hao, S.; Zhao, M.; Tsutsui, H.; Nishiguchi, S.; Sheng, J. Pu-erh tea extract ameliorates high-fat diet-induced nonalcoholic steatohepatitis and insulin resistance by modulating hepatic IL-6/STAT3 signaling in mice. J. Gastroenterol. 2016, 51, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Way, T.D.; Lin, H.Y.; Kuo, D.H.; Tsai, S.J.; Shieh, J.C.; Wu, J.C.; Lee, M.R.; Lin, J.K. Pu-erh tea attenuates hyperlipogenesis and induces hepatoma cells growth arrest through activating amp-activated protein kinase (ampk) in human hepg2 cells. J. Agric. Food Chem. 2009, 57, 5257–5264. [Google Scholar] [CrossRef] [PubMed]
- Finotello, F.; Di Camillo, B. Measuring differential gene expression with RNA-Seq: Challenges and strategies for data analysis. Brief. Funct. Geno. 2014, 14, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, B.T.; Landry, J.-R. RNA-Seq-Quantitative measurement of expression through massively parallel RNA-Sequencing. Methods 2009, 48, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Schliebner, I.; Becher, R.; Hempel, M.; Deising, H.B.; Horbach, R. New gene models and alternative splicing in the maize pathogen colletotrichum graminicola revealed by RNA-Seq analysis. BMC Genom. 2014, 15, 842. [Google Scholar] [CrossRef] [PubMed]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAS reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Gubert, P.; Puntel, B.; Lehmen, T.; Bornhorst, J.; Avila, D.S.; Aschner, M.; Soares, F.A.A. Reversible reprotoxic effects of manganese through daf-16 transcription factor activation and vitellogenin downregulation in Caenorhabditis elegans. Life Sci. 2016, 151, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, X.; Wu, J.; Zhang, L.; Huang, J.; Zhang, Y.; Zou, X.; Liang, B. Iron overload coordinately promotes ferritin expression and fat accumulation in Caenorhabditis elegans. Genetics 2016, 203, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-K.; Lin, C.-L.; Liang, Y.-C.; Lin-Shiau, S.-Y.; Juan, I.M. Survey of catechins, gallic acid, and methylxanthines in green, oolong, pu-erh, and black teas. J. Agric. Food Chem. 1998, 46, 3635–3642. [Google Scholar] [CrossRef]
- Jang, A.; Srinivasan, P.; Lee, N.Y.; Song, H.P.; Lee, J.W.; Lee, M.; Jo, C. Comparison of hypolipidemic activity of synthetic gallic acid-linoleic acid ester with mixture of gallic acid and linoleic acid, gallic acid, and linoleic acid on high-fat diet induced obesity in C57BL/6 Cr Slc mice. Chem.-Biol. Int. 2008, 174, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Doan, K.V.; Ko, C.M.; Kinyua, A.W.; Yang, D.J.; Choi, Y.H.; Oh, I.Y.; Nguyen, N.M.; Ko, A.; Choi, J.W.; Jeong, Y.; et al. Gallic acid regulates body weight and glucose homeostasis through ampk activation. Endocrinol. 2015, 156, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Huo, T.I.; Cheng, H.Y.; Tsai, J.C.; Liao, J.W.; Lee, M.S.; Qin, X.M.; Hsieh, M.T.; Pao, L.H.; Peng, W.H. Gallic acid ameliorated impaired glucose and lipid homeostasis in high fat diet-induced nafld mice. PLoS ONE 2014, 9, e96969. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Sohn, I.; Lee, Y.S.; Lee, Y.S. Hepatic gene expression profiles are altered by genistein supplementation in mice with diet-induced obesity. J. Nutr. 2005, 135, 33–41. [Google Scholar] [PubMed]
- Liu, Z.; Li, Q.; Huang, J.; Liang, Q.; Yan, Y.; Lin, H.; Xiao, W.; Lin, Y.; Zhang, S.; Tan, B.; et al. Proteomic analysis of the inhibitory effect of epigallocatechin gallate on lipid accumulation in human HepG2 cells. Proteom. Sci. 2013, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Spieth, J.; Blumenthal, T. The Caenorhabditis elegans vitellogenin gene family includes a gene encoding a distantly related protein. Mol. Cell Biol. 1985, 5, 2495–2501. [Google Scholar] [CrossRef] [PubMed]
- Spieth, J.; Denison, K.; Kirtland, S.; Cane, J.; Blumenthal, T. The C. elegans vitellogenin genes: Short sequence repeats in the promoter regions and homology to the vertebrate genes. Nucleic Acids Res. 1985, 13, 5283–5295. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, H.; Zhang, Z.; Chang, Q.; Liao, S.; Zhang, L.; Li, Y.; Wu, D.; Liang, B. Transcriptome profiles using next-generation sequencing reveal liver changes in the early stage of diabetes in tree shrew (Tupaia belangeri chinensis). J. Diabet. Res. 2016, 2016, 6238526. [Google Scholar] [CrossRef] [PubMed]
- Brooks, K.K.; Liang, B.; Watts, J.L. The influence of bacterial diet on fat storage in C. elegans. PLoS ONE 2009, 4, e7545. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Ferguson, K.; Kadyk, L.; Watts, J.L. The role of nuclear receptor NHR-64 in fat storage regulation in Caenorhabditis elegans. PLoS ONE 2010, 5, e9869. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.J.; Browse, J.; Watts, J.L. Fatty acid desaturation and the regulation of adiposity in Caenorhabditis elegans. Genetics 2007, 176, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.L.; Browse, J. Genetic dissection of polyunsaturated fatty acid synthesis in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2002, 99, 5854–5859. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Symbol | Biological Function | Fold Change | p-Value | ||
|---|---|---|---|---|---|---|
| 0.025 g/mL | 0.05 g/mL | 0.025 g/mL | 0.05 g/mL | |||
| XLOC_000825 | col-7 | Nematode cuticle collagen | (−)17.21 | (−)86.04 | 2.6 × 10−6 | 2.6 × 10−10 |
| XLOC_010817 | col-8 | (S) | (−)68.00 | 2.11 × 10−6 | ||
| XLOC_002700 | col-62 | (S) | (−)15.32 | (−)45.24 | 3.02 × 10−6 | 2.32 × 10−8 |
| XLOC_007760 | col-81 | (S) | (−)9.33 | 2.93 × 10−6 | ||
| XLOC_014356 | col-126 | (S) | (−)19.12 | 5.21 × 10−6 | ||
| XLOC_021979 | col-127 | (S) | (−)19.12 | 5.21 × 10−6 | ||
| XLOC_028294 | col-146 | (S) | (−)13.27 | 2.88 × 10−6 | ||
| XLOC_035276 | vit-1 | Lipid transport | (−)6.53 | 2.79 × 10−5 | ||
| XLOC_034756 | vit-2 | (S) | (−)9.68 | 1.56 × 10−5 | ||
| XLOC_037318 | vit-3 | (S) | (−)7.79 | 1.72 × 10−5 | ||
| XLOC_037317 | vit-4 | (S) | (−)8.16 | (−)13.68 | 5.83 × 10−6 | 9.06 × 10−8 |
| XLOC_034472 | vit-5 | (S) | (−)18.37 | (−)73.18 | 1.81 × 10−9 | 2.86 × 10−14 |
| XLOC_034954 | C45B2.1 | Imported | (−)19.39 | 1.55 × 10−5 | ||
| XLOC_028510 | D1054.11 | (S) | (−)14.58 | 4.25 × 10−5 | ||
| XLOC_005888 | K10H10.4 | Protein binding | (−)31.48 | 5.45 × 10−6 | ||
| XLOC_006818 | dct-5 | Defence responce | (−)34.82 | 5.74 × 10−6 | ||
| XLOC_031134 | abu-7 | Protect organism | (−)12.82 | 4.41 × 10−5 | ||
| XLOC_000825 | grl-4 | Structural molecule activity | (−)13.39 | 2.27 × 10−6 | ||
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, R.-Y.; Hao, J.; Ding, Y.-H.; Che, Y.-Y.; Zou, X.-J.; Liang, B. Transcriptome Profile Reveals that Pu-Erh Tea Represses the Expression of Vitellogenin Family to Reduce Fat Accumulation in Caenorhabditis elegans. Molecules 2016, 21, 1379. https://doi.org/10.3390/molecules21101379
Xiao R-Y, Hao J, Ding Y-H, Che Y-Y, Zou X-J, Liang B. Transcriptome Profile Reveals that Pu-Erh Tea Represses the Expression of Vitellogenin Family to Reduce Fat Accumulation in Caenorhabditis elegans. Molecules. 2016; 21(10):1379. https://doi.org/10.3390/molecules21101379
Chicago/Turabian StyleXiao, Ru-Yue, Junjun Hao, Yi-Hong Ding, Yan-Yun Che, Xiao-Ju Zou, and Bin Liang. 2016. "Transcriptome Profile Reveals that Pu-Erh Tea Represses the Expression of Vitellogenin Family to Reduce Fat Accumulation in Caenorhabditis elegans" Molecules 21, no. 10: 1379. https://doi.org/10.3390/molecules21101379
APA StyleXiao, R. -Y., Hao, J., Ding, Y. -H., Che, Y. -Y., Zou, X. -J., & Liang, B. (2016). Transcriptome Profile Reveals that Pu-Erh Tea Represses the Expression of Vitellogenin Family to Reduce Fat Accumulation in Caenorhabditis elegans. Molecules, 21(10), 1379. https://doi.org/10.3390/molecules21101379