
Identification and Characterisation of the Antimicrobial Peptide, Phylloseptin-PT, from the Skin Secretion of Phyllomedusa tarsius, and Comparison of Activity with Designed, Cationicity-Enhanced Analogues and Diastereomers

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
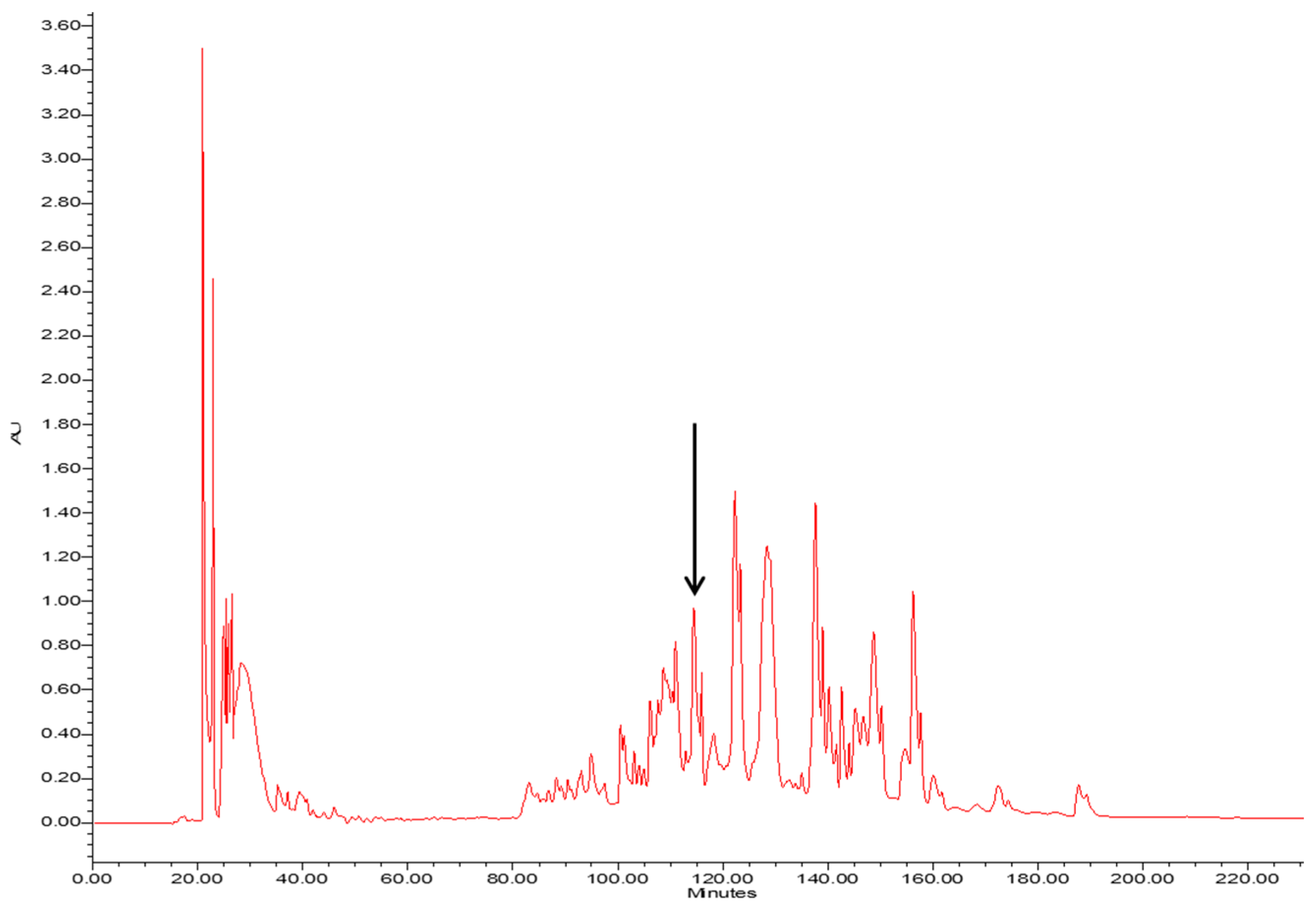
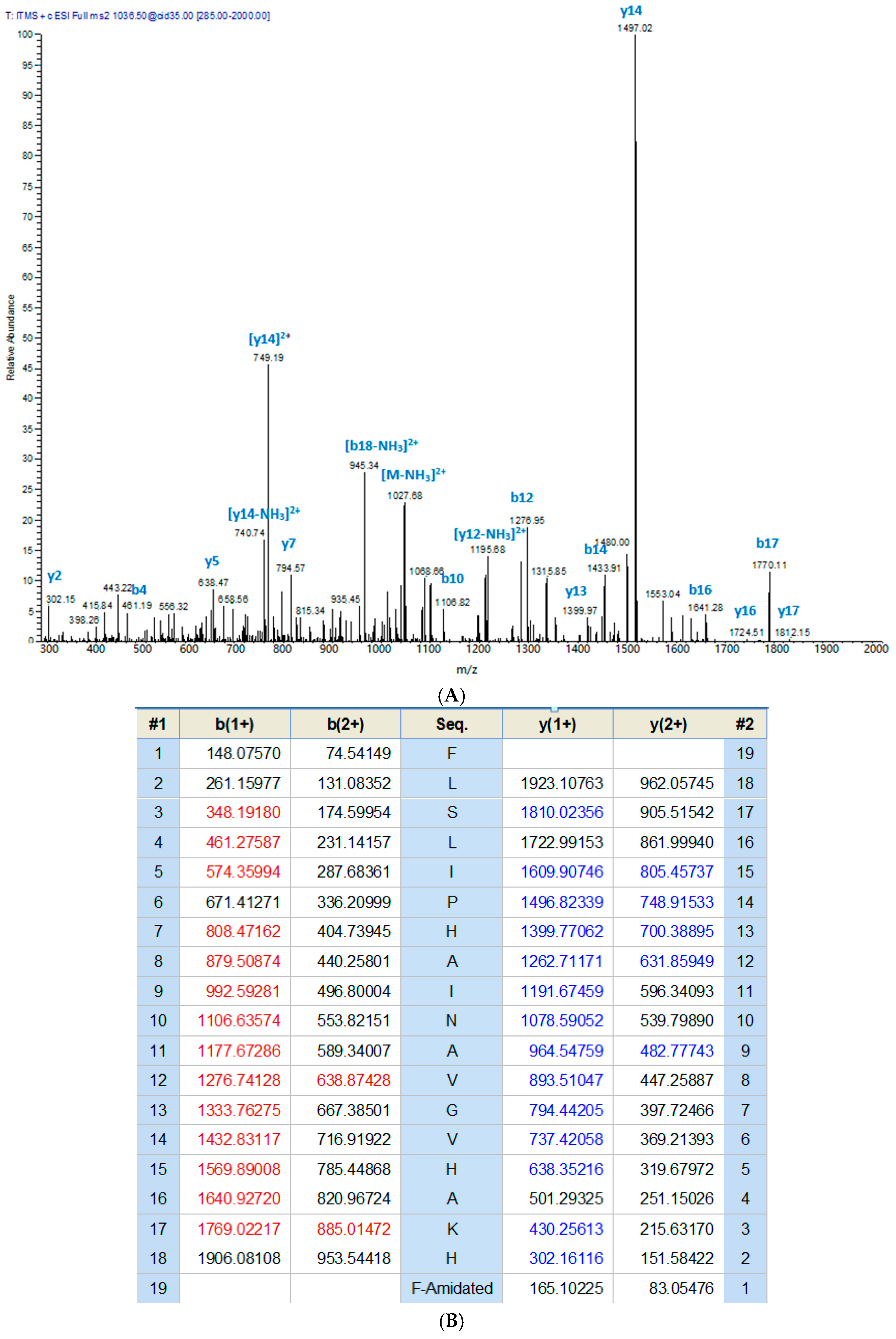
2.1. Molecular Cloning and Identification of Phylloseptin-PT Precursor-Encoding cDNA from the Skin Secretion of P. tarsius
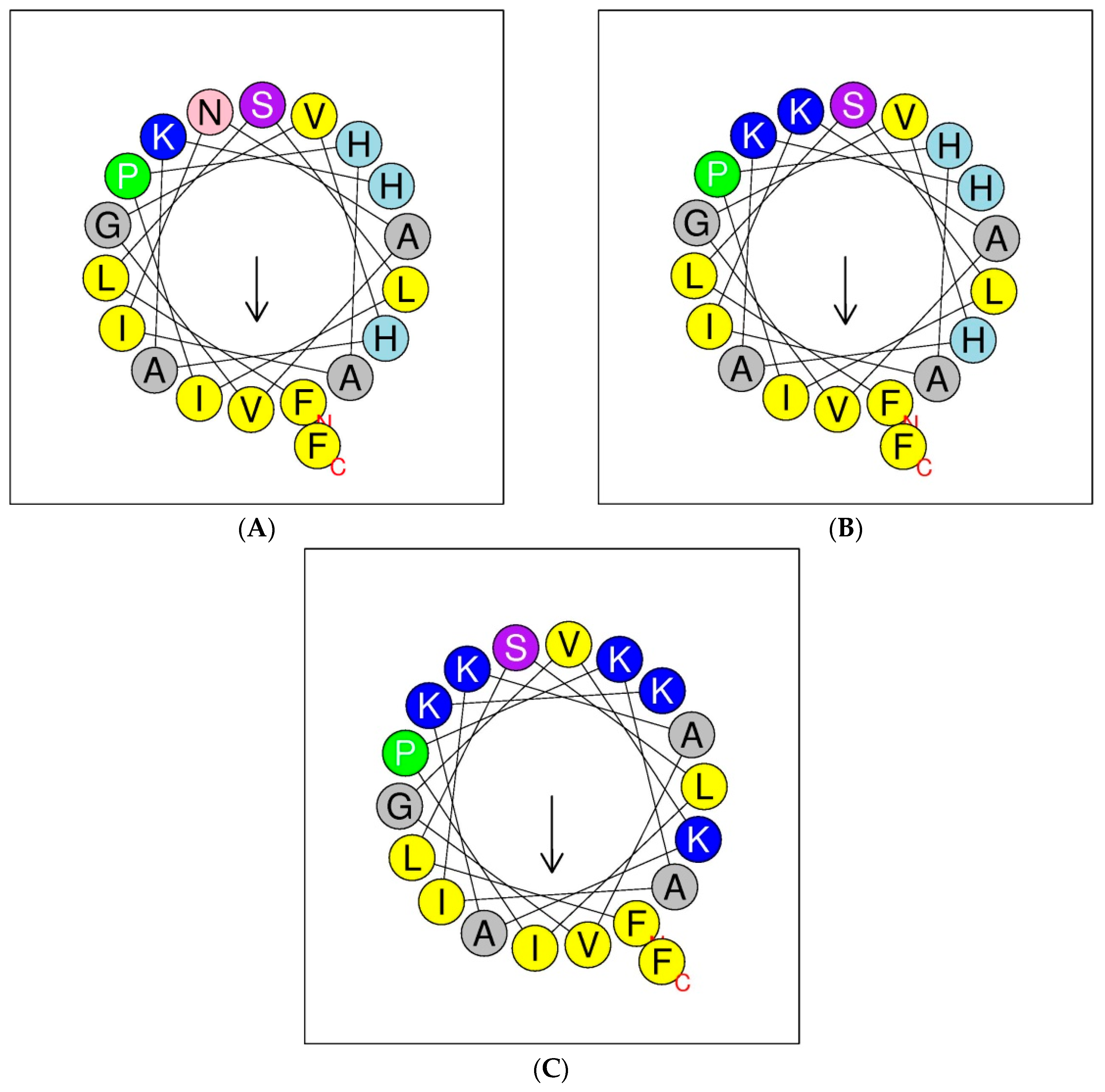
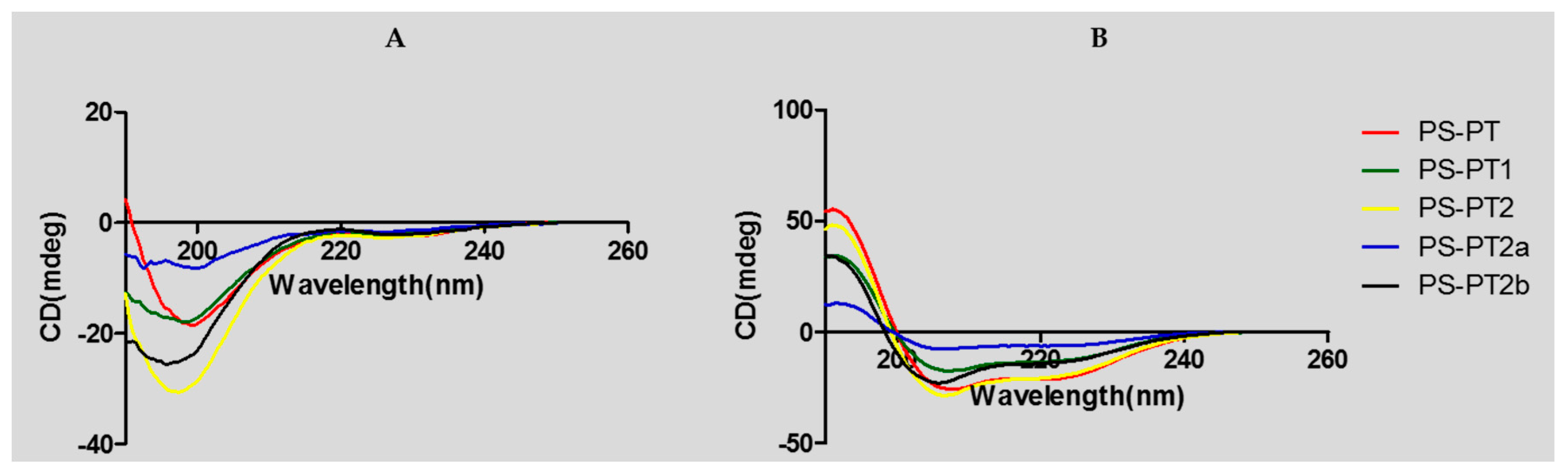
2.2. Design, Synthesis, Predicted Physicochemical Parameters, and Secondary Structures of Peptides
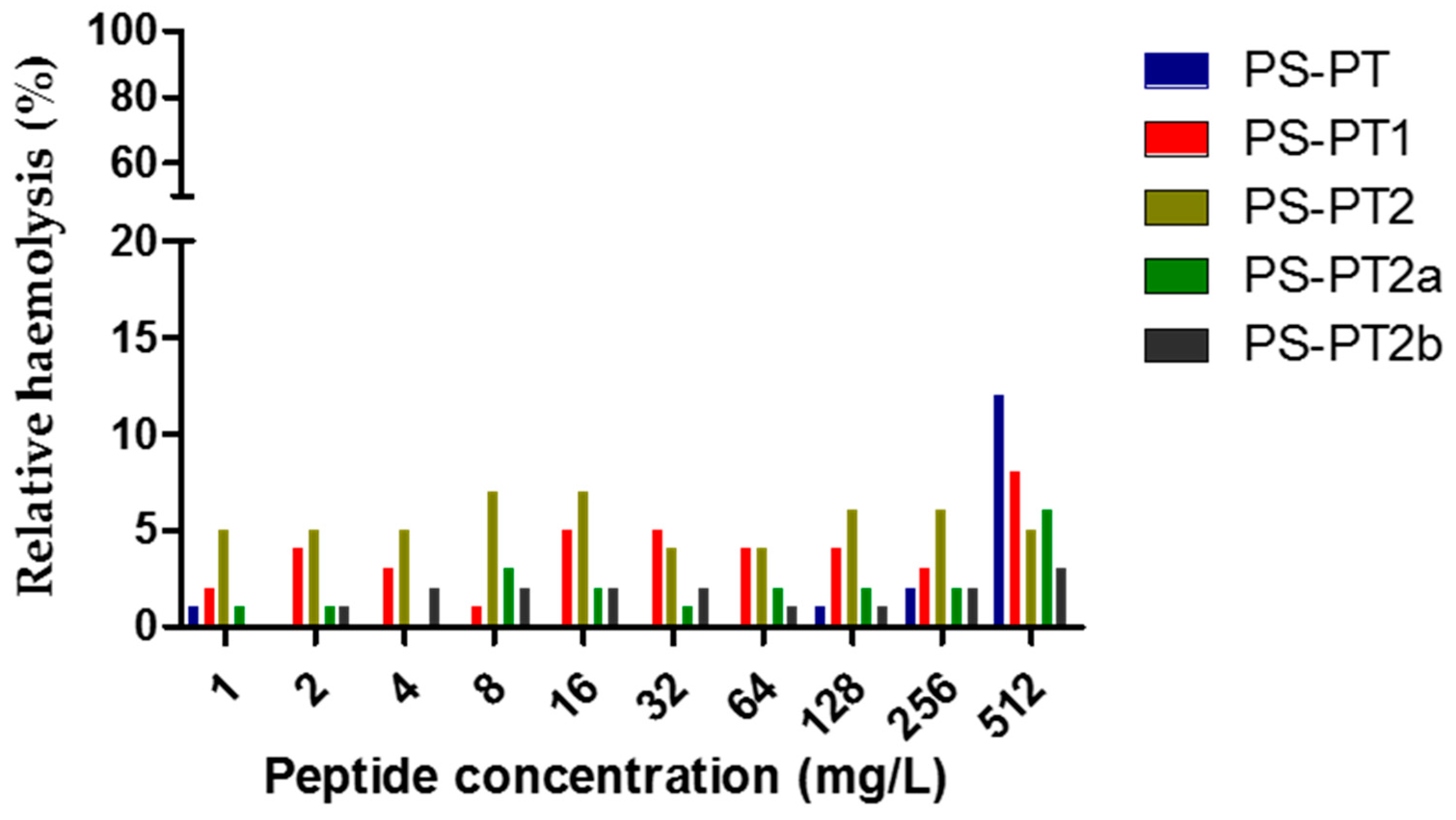
2.3. Antimicrobial and Haemolysis Assays
2.4. Comparison of the Stability of Diastereomers
3. Discussion
4. Materials and Methods
4.1. Acquisition of Phyllomedusa Tarsius Skin Secretions
4.2. Molecular Cloning of Phylloseptin-PT Precursor-Encoding cDNA from a Skin Secretion-Derived cDNA Library of P. tarsius
4.3. Identification and Structural Characterization of the Putative Antimicrobial Peptide
4.4. Determination of Peptide Secondary Structures and Prediction of Antimicrobial and Physicochemical Properties
4.5. Peptide Synthesiss
4.6. Antimicrobial Assay
4.7. Haemolysis Assay
4.8. Stability and Degradation of Peptides
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| RP-HPLC: | Reverse-phase high-performance liquid chromatography |
| MALDI-TOF/MS: | Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry |
| CHCA: | α-cyano-4-hydroxycinnamic acid |
| RACE: | Rapid Amplification of cDNA Ends |
| Fmoc: | 9-fluorenylmethyloxycarbonyl |
| TFA: | Trifluoroacetic Acid |
References
- Kaspar, A.A.; Reichert, J.M. Future directions for peptide therapeutics development. Drug Discov. Today 2013, 18, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, T.; Kyprianou, T.; Martinelli, F.G.; Oppici, C.A.; Heiligers, D.; Hills, D.; Calvo, X.R.; Verhaert, P. The emergence of peptides in the pharmaceutical business: From exploration to exploitation. EuPA Open Proteom. 2014, 4, 58–69. [Google Scholar] [CrossRef]
- Antosova, Z.; Mackova, M.; Kral, V.; Macek, T. Therapeutic application of peptides and proteins: parenteral forever? Trends Biotechnol. 2009, 27, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Pirri, G.; Nicoletto, S.F. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Cézard, C.; Silva-Pires, V.; Mullié, C.; Sonnet, P. Antibacterial peptides: A review. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; pp. 926–937. [Google Scholar]
- Wang, G.; Mishra, B.; Lau, K.; Lushnikova, T.; Golla, R.; Wang, X. Antimicrobial peptides in 2014. Pharmaceuticals 2015, 8, 123–150. [Google Scholar] [CrossRef] [PubMed]
- Bruno, B.J.; Miller, G.D.; Lim, C.S. Basics and recent advances in peptide and protein drug delivery. Ther. Deliv. 2013, 4, 1443–1467. [Google Scholar] [CrossRef] [PubMed]
- Proaño-Bolaños, C.; Zhou, M.; Wang, L.; Coloma, L.A.; Chen, T.; Shaw, C. Peptidomic approach identifies cruzioseptins, a new family of potent antimicrobial peptides in the splendid leaf frog, Cruziohyla calcarifer. J. Proteom. 2016, 146, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Ma, C.; Zhou, M.; Xi, X.; Li, L.; Wu, D.; Wang, L.; Lin, C.; Lopez, J.C.; Chen, T.; et al. Phylloseptin-PBa—A novel broad-spectrum antimicrobial peptide from the skin secretion of the Peruvian purple-sided leaf frog (Phyllomedusa baltea) which exhibits cancer cell cytotoxicity. Toxins (Basel). 2015, 7, 5182–5193. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.H.; Bjourson, A.J.; Orr, D.F.; Shaw, C.; McClean, S. A combined mass spectrometric and cDNA sequencing approach to the isolation and characterization of novel antimicrobial peptides from the skin secretions of Phyllomedusa hypochondrialis azurea. Peptides 2007, 28, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.J. Peptide vaccines for viral diseases. Sci. Prog. 1990, 74, 115–130. [Google Scholar] [PubMed]
- Choonara, B.F.; Choonara, Y.E.; Kumar, P.; Bijukumar, D.; du Toit, L.C.; Pillay, V. A review of advanced oral drug delivery technologies facilitating the protection and absorption of protein and peptide molecules. Biotechnol. Adv. 2014, 32, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Antimicrobial Peptides: Discovery, Design and Novel Therapeutic Strategies. In Advances in Molecular and Cellular Microbiology; The Centre for Agriculture and Bioscience International: Wallingford, UK, 2010; Volume 18. [Google Scholar]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef] [PubMed]
- Leite, J.R.S.A.; Silva, L.P.; Rodrigues, M.I.S.; Prates, M.V.; Brand, G.D.; Lacava, B.M.; Azevedo, R.B.; Bocca, A.L.; Albuquerque, S.; Bloch, C. Phylloseptins: A novel class of anti-bacterial and anti-protozoan peptides from the Phyllomedusa genus. Peptides 2005, 26, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.Y.; Lee, Y.T.; Park, C.B.; Lee, K.H.; Kim, S.C.; Choi, B.S. Structural and functional implications of a proline residue in the antimicrobial peptide gaegurin. Eur. J. Biochem. 1999, 266, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Cordes, F.S.; Bright, J.N.; Sansom, M.S.P. Proline-induced distortions of transmembrane helices. J. Mol. Biol. 2002, 323, 951–960. [Google Scholar] [CrossRef]
- Resende, J.M.; Moraes, C.M.; Prates, M.V.; Cesar, A.; Almeida, F.C.L.; Mundim, N.C.C.R.; Valente, A.P.; Bemquerer, M.P.; Piló-Veloso, D.; Bechinger, B. Solution NMR structures of the antimicrobial peptides phylloseptin-1, -2, and -3 and biological activity: The role of charges and hydrogen bonding interactions in stabilizing helix conformations. Peptides 2008, 29, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Simmaco, M.; Mignogna, G.; Barra, D. Antimicrobial peptides from amphibian skin: What do they tell us? Pept. Sci. 1998, 47, 435–450. [Google Scholar] [CrossRef]
- ENZYME entry 1. Available online: http://enzyme.expasy.org/EC/3.4.21.4 (accessed on 25 February 2016).
- Tyler, M.J.; Stone, D.J.M.; Bowie, J.H. A novel method for the release and collection of dermal, glandular secretions from the skin of frogs. J. Pharmacol. Toxicol. Methods 1992, 28, 199–200. [Google Scholar] [CrossRef]
- Pierre, T.N.; Seon, A.A.; Amiche, M.; Nicolas, P. Phylloxin, a novel peptide antibiotic of the dermaseptin family of antimicrobial/opioid peptide precursors. Eur. J. Biochem. 2000, 267, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Wechselberger, C.; Severini, C.; Kreil, G.; Negri, L. A new opioid peptide predicted from cloned cDNAs from skin of Pachymedusa dacnicolor and Agalychnis annae. FEBS Lett. 1998, 429, 41–43. [Google Scholar] [CrossRef]
- Louis-Jeune, C.; Andrade-Navarro, M.A.; Perez-Iratxeta, C. Prediction of protein secondary structure from circular dichroism using theoretically derived spectra. Proteins Struct. Funct. Bioinform. 2012, 80, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Rhone-Alpes Bioinformatic Pole Gerland Site (PRABI)-Institute of Biology and Protein Chemisty NPS@: GOR4 Secondary Structure Prediction. Available online: https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html (accessed on 24 February 2016).
- HeliQuest ComputParam form version2. Available online: http://heliquest.ipmc.cnrs.fr/cgi-bin/ComputParamsV2.py (accessed on 24 February 2016).
- Armstrong, D.; Zidovetzki, R. Helical Wheel Projections. Available online: http://rzlab.ucr.edu/scripts/wheel/wheel.cgi (accessed on 25 February 2016).
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific α-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef] [PubMed]
- Dathe, M.; Wieprecht, T. Structural features of helical antimicrobial peptides: Their potential to modulate activity on model membranes and biological cells. Biochim. Biophys. Acta (BBA)-Biomembr. 1999, 1462, 71–87. [Google Scholar] [CrossRef]
- Dathe, M.; Wieprecht, T.; Nikolenko, H.; Handel, L.; Maloy, W.L.; MacDonald, D.L.; Beyermann, M.; Bienert, M. Hydrophobicity, hydrophobic moment and angle subtended by charged residues modulate antibacterial and haemolytic activity of amphipathic helical peptides. FEBS Lett. 1997, 403, 208–212. [Google Scholar] [CrossRef]
- Chen, Y.; Mant, C.T.; Farmer, S.W.; Hancock, R.E.W.; Vasil, M.L.; Hodges, R.S. Rational design of α-helical antimicrobial peptides with enhanced activities and specificity/therapeutic index. J. Biol. Chem. 2005, 280, 12316–12329. [Google Scholar] [CrossRef] [PubMed]
- Tiantang, D.; Zigang, T.; Jianhua, W. The structural parameters-functional activity relationship of alpha-helical antimicrobial peptides. China Biotechnol. 2007, 27, 116–119. [Google Scholar]
- Sample Availability: Samples of the compounds PS-PT, PS-PT1, PS-PT2, PS-PT2a and PS-PT2b are available from the authors.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Hydrophobicity (H) | Hydrophobic Moment (μH) | % Helix | Net Charge |
|---|---|---|---|---|---|
| PS-PT | FLSLIPHAINAVGVHAKHF-NH2 | 0.707 | 0.418 | 37.92 | 2 |
| PS-PT1 | FLSLIPHAIKAVGVHAKHF-NH2 | 0.686 | 0.438 | 19.48 | 3 |
| PS-PT2 | FLSLIPKAIKAVGVKAKKF-NH2 | 0.509 | 0.497 | 37.91 | 6 |
| PS-PT2a | FLSLIPKAIKAVGVKAKKF-NH2 | 0.509 | 0.497 | 1.06 | 6 |
| PS-PT2b | FLSLIPKAIKAVGVKAKKF-NH2 | 0.509 | 0.497 | 23.92 | 6 |
| Peptide | MIC (mg/L) | MBC (mg/L) | ||||
|---|---|---|---|---|---|---|
| S. aureus | E. coli | C. albicans | S. aureus | E. coli | C. albicans | |
| PS-PT | 512 | >512 | 512 | >512 | >512 | >512 |
| PS-PT1 | 128 | 512 | 128 | 256 | >512 | 256 |
| PS-PT2 | 64 | 512 | 16 | 64 | >512 | 32 |
| PS-PT2a | >512 | >512 | 256 | >512 | >512 | 256 |
| PS-PT2b | 256 | 256 | 32 | 512 | 256 | 64 |
| Conditions | PS-PT2 | PS-PT2a | PS-PT2b |
|---|---|---|---|
| Trypsin | 1–19 FLSLIPKAIKAVGVKAKKF | 1–19 FLSLIPKAIKAVGVKAKKF | 1–19 FLSLIPKAIKAVGVKAKKF |
| 1–7 FLSLIPK | 1–10 FLSLIPKAIK | ||
| 8–19 AIKAVGVKAKKF | 11–19 AVGVKAKKF | ||
| 1–17 FLSLIPKAIKAVGVKAK | |||
| Serum | 1–19 FLSLIPKAIKAVGVKAKKF | 1–19 FLSLIPKAIKAVGVKAKKF | 1–19 FLSLIPKAIKAVGVKAKKF |
| 1–7 FLSLIPK | 3–19 SLIPKAIKAVGVKAKKF | 1–14 FLSLIPKAIKAVGVK | |
| 8–19 AIKAVGVKAKKF | 2–7 LSLIPK | 1–15 FLSLIPKAIKAVGVKA | |
| 1–16 FLSLIPKAIKAVGVKA | 1–17 FLSLIPKAIKAVGVKAK | ||
| 3–19 SLIPKAIKAVGVKAKKF |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Wu, D.; Xi, X.; Wu, Y.; Ma, C.; Zhou, M.; Wang, L.; Yang, M.; Chen, T.; Shaw, C. Identification and Characterisation of the Antimicrobial Peptide, Phylloseptin-PT, from the Skin Secretion of Phyllomedusa tarsius, and Comparison of Activity with Designed, Cationicity-Enhanced Analogues and Diastereomers. Molecules 2016, 21, 1667. https://doi.org/10.3390/molecules21121667
Gao Y, Wu D, Xi X, Wu Y, Ma C, Zhou M, Wang L, Yang M, Chen T, Shaw C. Identification and Characterisation of the Antimicrobial Peptide, Phylloseptin-PT, from the Skin Secretion of Phyllomedusa tarsius, and Comparison of Activity with Designed, Cationicity-Enhanced Analogues and Diastereomers. Molecules. 2016; 21(12):1667. https://doi.org/10.3390/molecules21121667
Chicago/Turabian StyleGao, Yitian, Di Wu, Xinping Xi, Yue Wu, Chengbang Ma, Mei Zhou, Lei Wang, Mu Yang, Tianbao Chen, and Chris Shaw. 2016. "Identification and Characterisation of the Antimicrobial Peptide, Phylloseptin-PT, from the Skin Secretion of Phyllomedusa tarsius, and Comparison of Activity with Designed, Cationicity-Enhanced Analogues and Diastereomers" Molecules 21, no. 12: 1667. https://doi.org/10.3390/molecules21121667
APA StyleGao, Y., Wu, D., Xi, X., Wu, Y., Ma, C., Zhou, M., Wang, L., Yang, M., Chen, T., & Shaw, C. (2016). Identification and Characterisation of the Antimicrobial Peptide, Phylloseptin-PT, from the Skin Secretion of Phyllomedusa tarsius, and Comparison of Activity with Designed, Cationicity-Enhanced Analogues and Diastereomers. Molecules, 21(12), 1667. https://doi.org/10.3390/molecules21121667