
Antioxidant and Anti-Inflammatory Effects of Chaenomeles sinensis Leaf Extracts on LPS-Stimulated RAW 264.7 Cells

Abstract
:1. Introduction
2. Results
2.1. Extraction Yields, Total Polyphenol and Flavonoid Contents of CSL Extracts
2.2. Antioxidant Activity of CSL Extracts
2.3. Radical Scavenging Activity by ESR Measurement
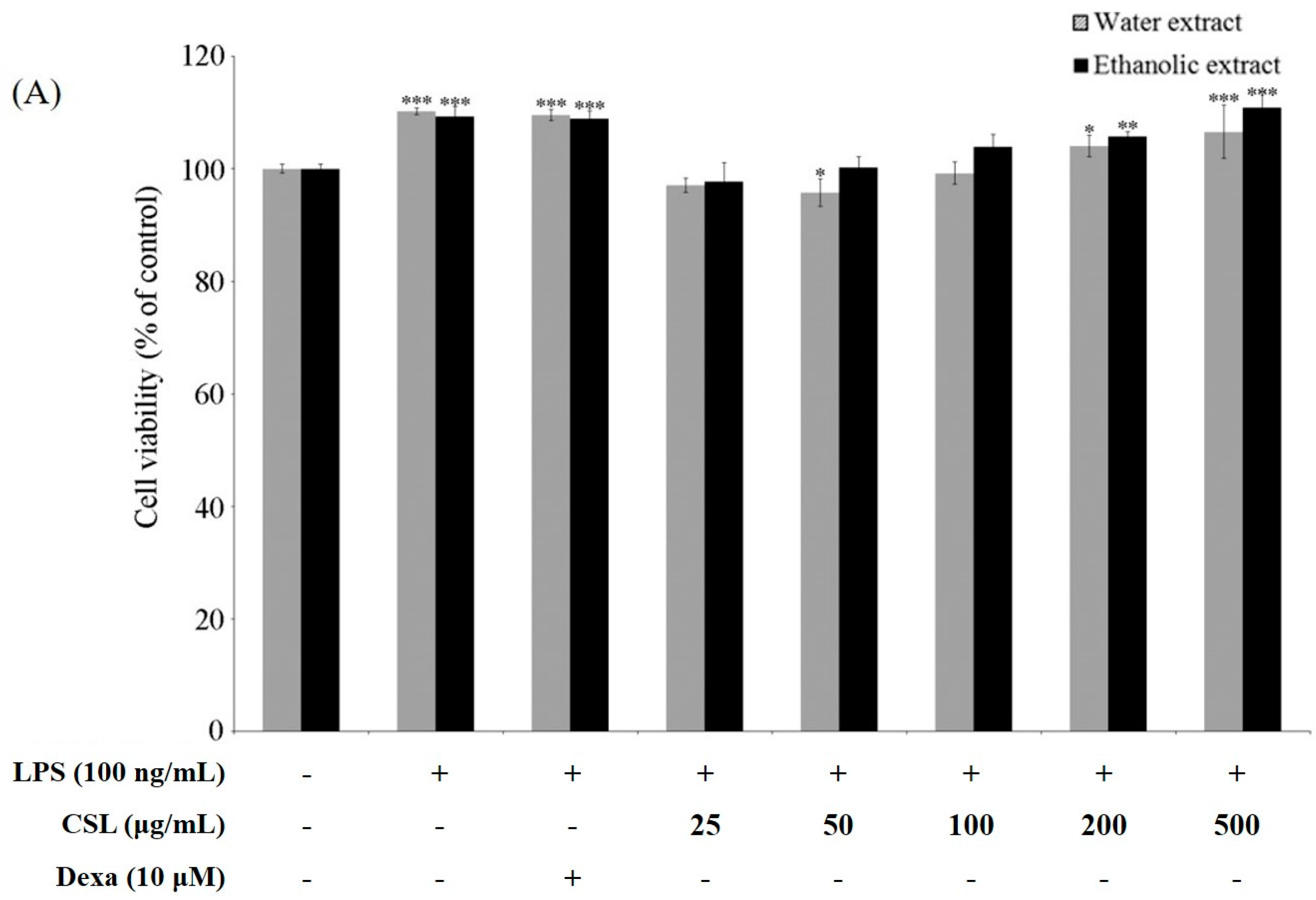
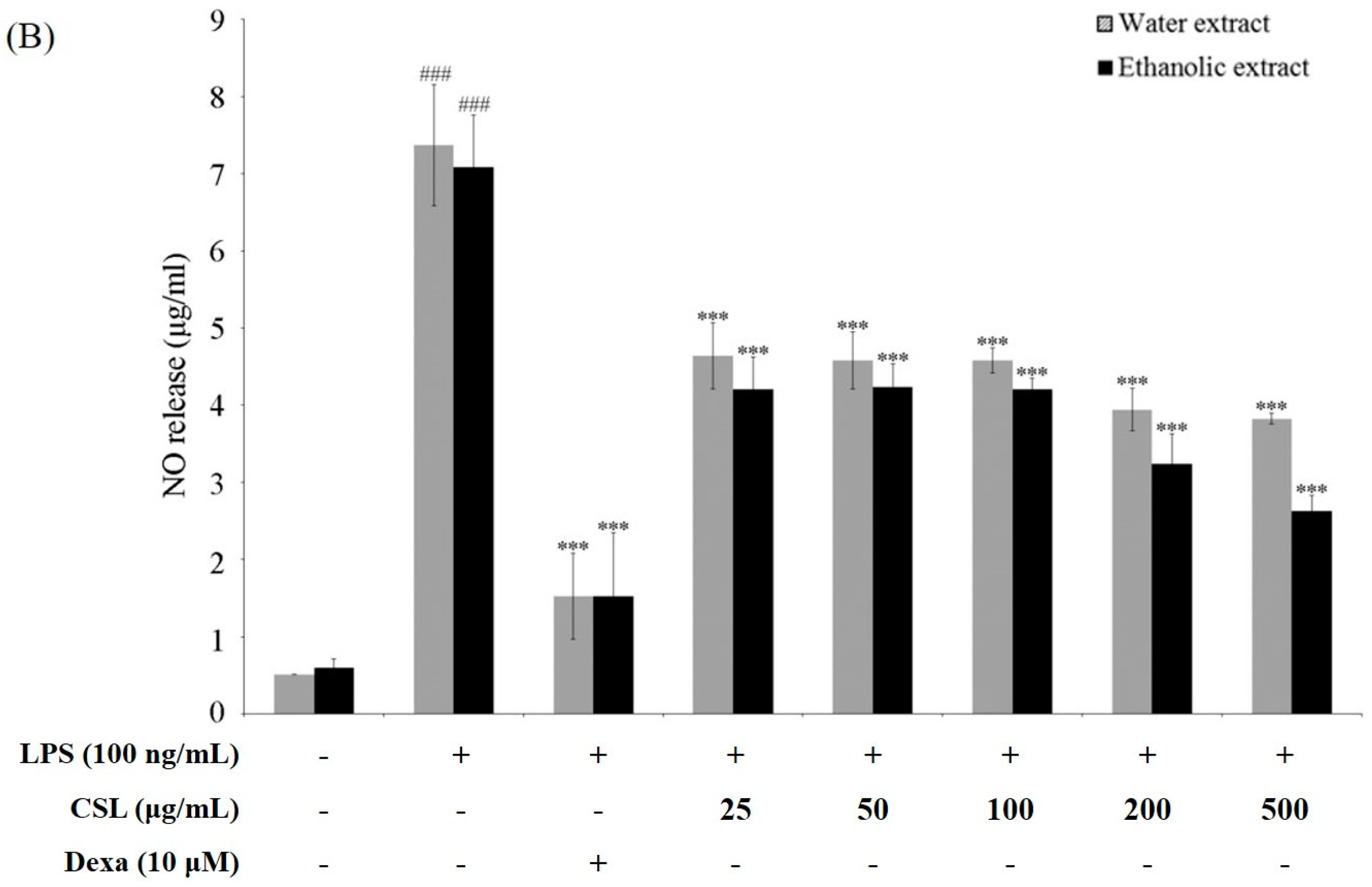
2.4. Cell Viability and Production of NO
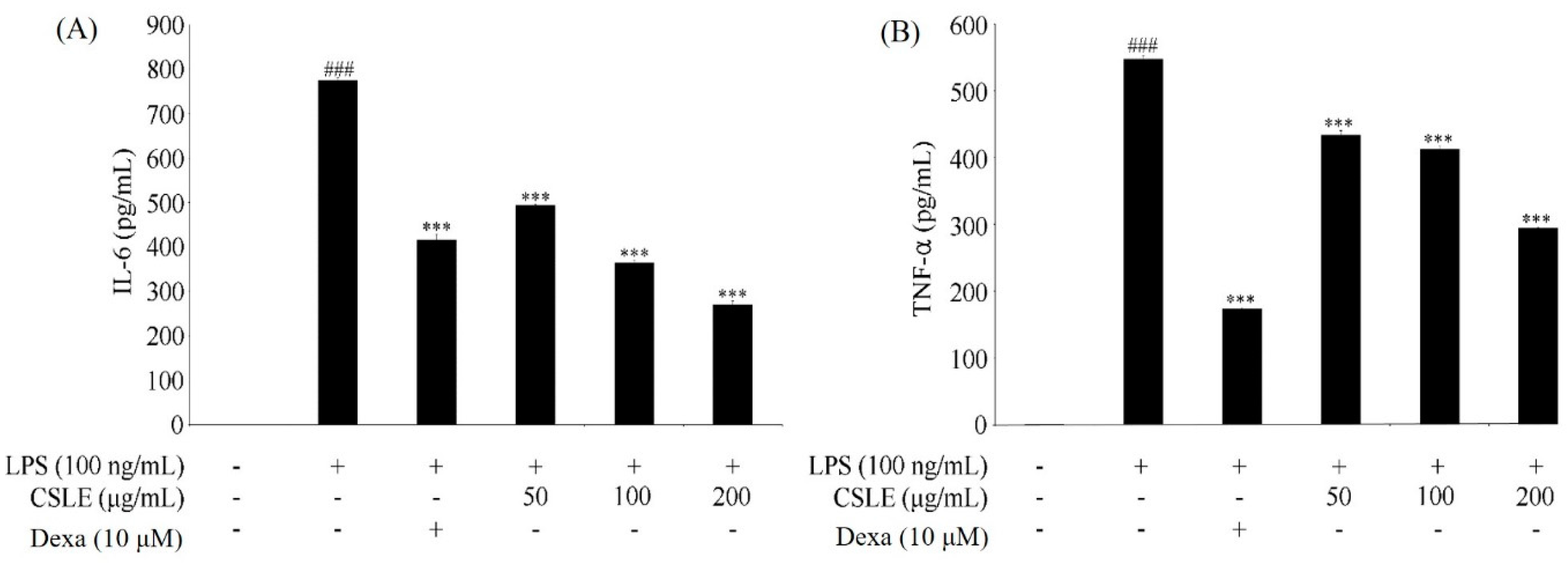
2.5. Inhibition of LPS-Induced Proinflammatory Cytokine Production by CSLE in LPS-Stimulated RAW 264.7 Cells
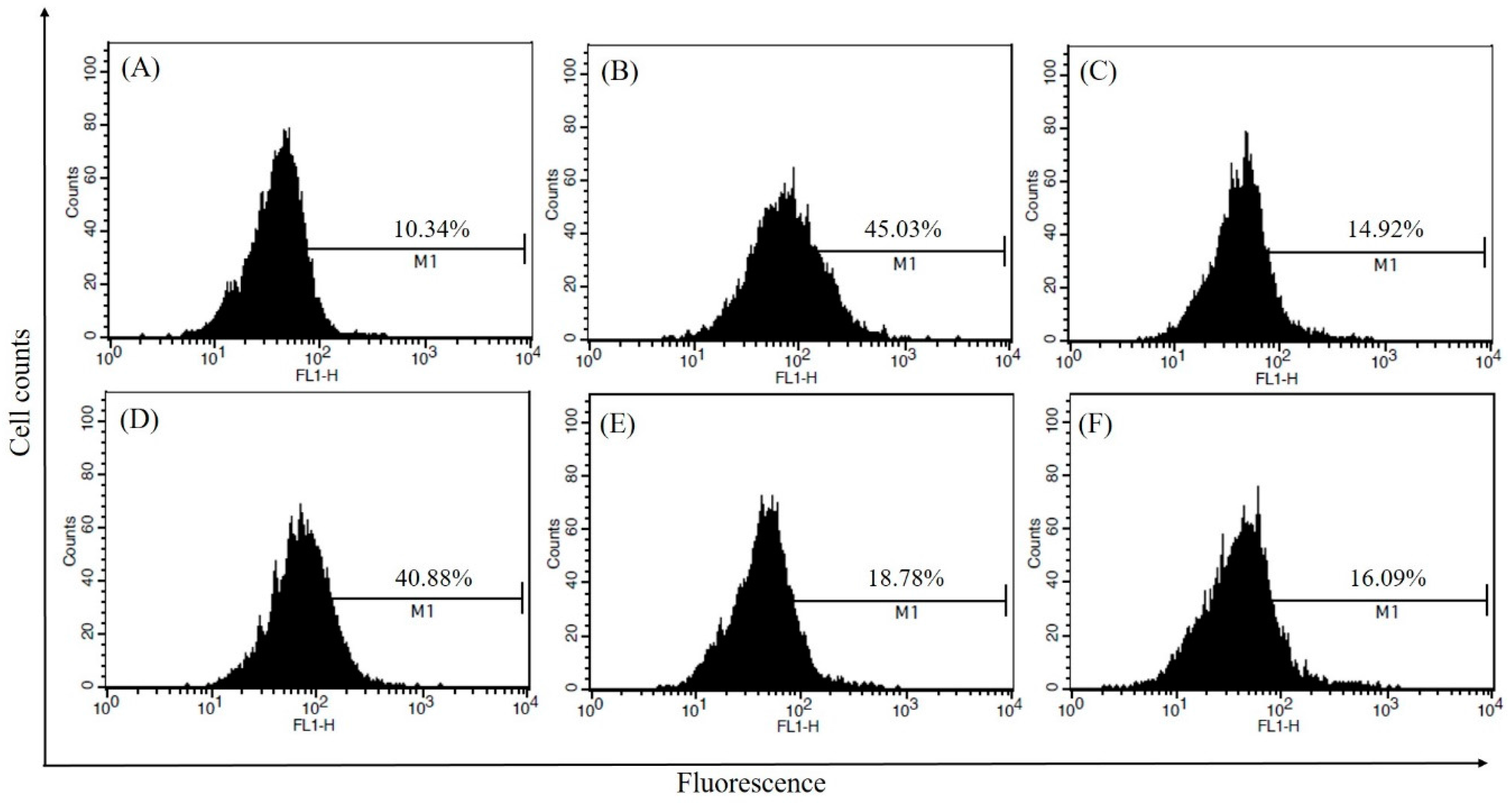
2.6. Inhibitory Effects of CSLE on ROS Generation
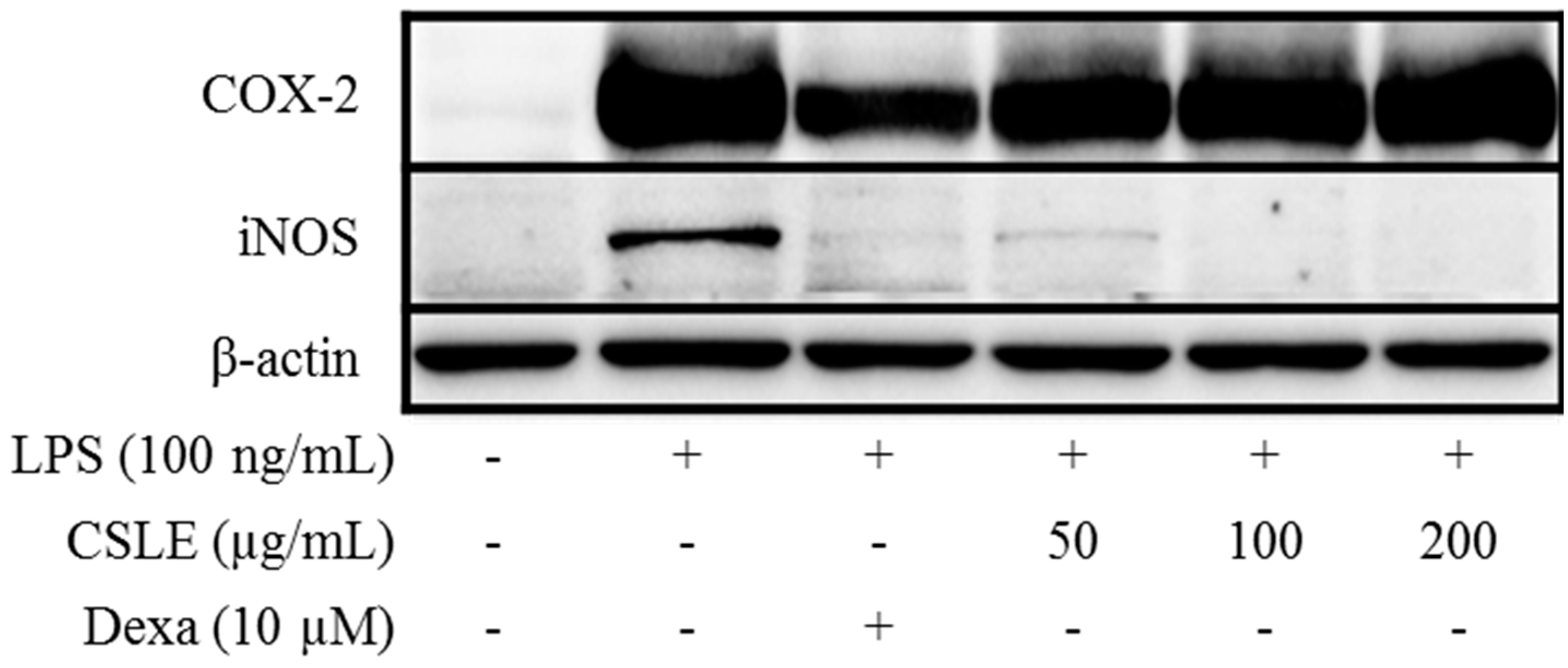
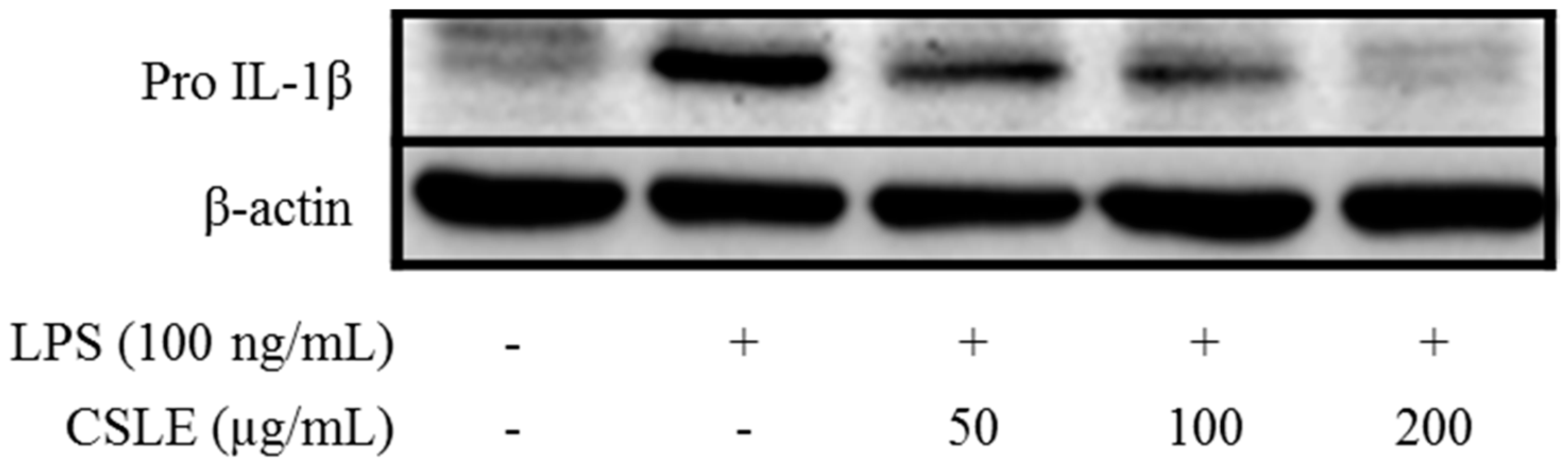
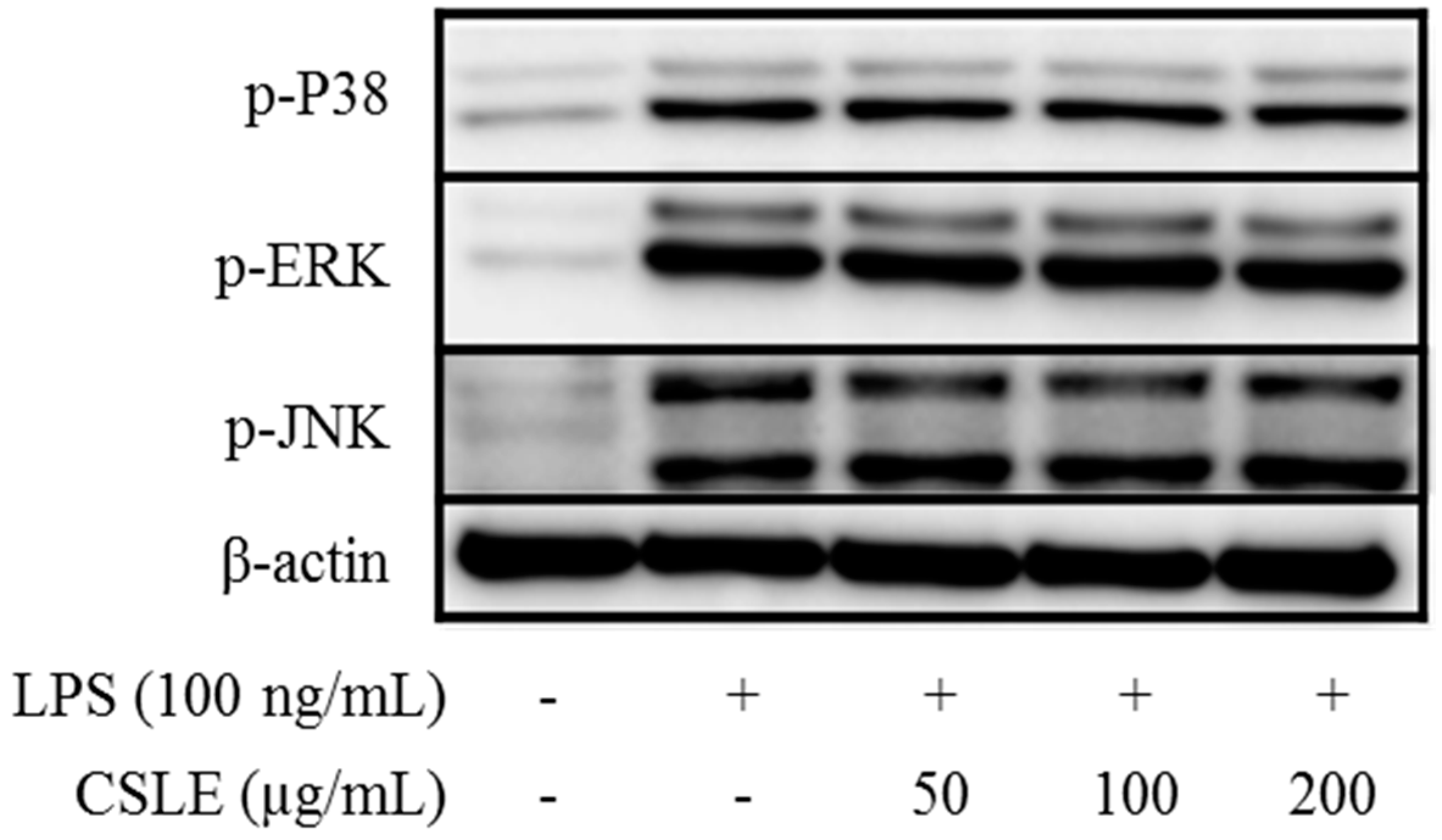
2.7. Absence of CSLE Affect on MyD-88 Dependent Signaling Pathways
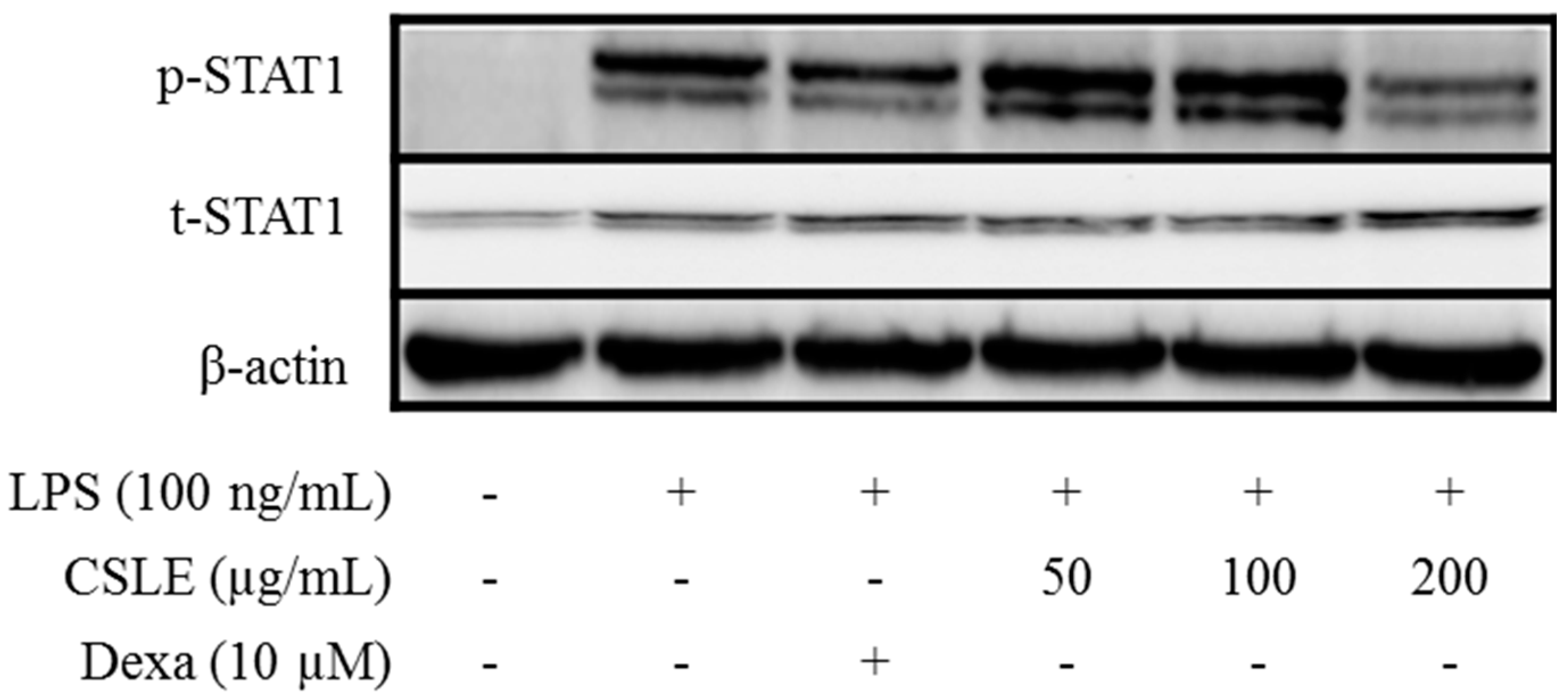
2.8. TRIF-Dependent Signaling Pathways Inhibit CSLE
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of the Extract
4.3. Antioxidant Activity of the Chaenomeles Sinensis Leaves Extract
4.3.1. Determination of Total Polypenol Contents
4.3.2. Determination of Total Flavonoid Contents
4.3.3. ABTS Radical Scavenging Activity
4.3.4. Ferric Reducing Antioxidant Power (FRAP) Assay
4.3.5. Radical Scavenging Activity by ESR Measurement
DPPH Radical Scavenging Activity
Alkyl Radical Scavenging Activity
Hydroxyl (OH) Radical Scavenging Activity
4.4. Cell Culture
4.5. Determination of Cell Viability Assay
4.6. Determination of Nitric Oxide (NO) Production
4.7. Determination of Intracellular ROS Using Flow Cytometry
4.8. Measurement of CYtokine (IL-6 and TNF-α)
4.9. Western Blot Analysis
4.10. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sawai, R.; Kuroda, K.; Shibata, T.; Gomyou, R.; Osawa, K.; Shimizu, K. Anti-influenza virus activity of Chaenomeles sinensis. J. Ethnopharmacol. 2008, 118, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Osawa, K.; Yasuda, H.; Morita, H.; Takeya, K.; Itokawa, H. Antibacterial and Antihemolytic Activity of Triterpenes and beta-Sitosterol Isolated from Chinese Quince (Chaenomeles sinensis). Nat. Med. 1997, 51, 365–367. [Google Scholar]
- Osawa, K.; Miyazaki, K.; Imai, H.; Arakawa, T.; Yasuda, H.; Takeya, K. Inhibitory effects of Chinese quince (Chaenomeles sinensis) on hyaluronidase and histamine release from rat mast cells. Nat. Med. 1999, 53, 188–193. [Google Scholar]
- Osawa, K.; Arakawa, T.; Shimura, S.; Takeya, K. New quinic acid derivatives from the fruits of Chaenomeles sinensis (Chinese quince). Jpn. J. Pharm. 2001, 55, 255–257. [Google Scholar]
- Oku, H.; Ueda, Y.; Ishiguro, K. Antipruritic effects of the fruits of Chaenomeles sinensis. Biol. Pharm. Bull. 2003, 26, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Majno, G. The Healing Hand: Man and wound in the ancient world. Plast. Reconstr. Surg. 1976, 57, 230. [Google Scholar] [CrossRef]
- Cheng, B.C.Y.; Ma, X.Q.; Kwan, H.Y.; Tse, K.W.; Cao, H.H.; Su, T.; Shu, X.; Wu, Z.-Z.; Yu, Z.-L. A herbal formula consisting of Rosae Multiflorae Fructus and Lonicerae Japonicae Flos inhibits inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. J. Ethnopharmacol. 2014, 153, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Willoughby, D.A.; Gilroy, D.W. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat. Rev. Immunol. 2002, 2, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Boscá, L.; Zeini, M.; Través, P.G.; Hortelano, S. Nitric oxide and cell viability in inflammatory cells: A role for NO in macrophage function and fate. Toxicology 2005, 208, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, S.; Legumsky, M.; Kohen, R.; Kanner, J. A novel function of red wine polyphenols in humans: Prevention of absorption of cytotoxic lipid peroxidation products. FASEB J. 2008, 22, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Schaich, K.M.; Tian, X.; Xie, J. Reprint of “Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays”. J. Funct. Foods 2015, 18, 782–796. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Toshchakov, V.; Jones, B.W.; Perera, P.-Y.; Thomas, K.; Cody, M.J.; Zhang, S.; Williams, B.R.; Major, J.; Hamilton, T.A.; Fenton, M.J. TLR4, but not TLR2, mediates IFN-β-induced STAT1α/β-dependent gene expression in macrophages. Nat. Immunol. 2002, 3, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Samavati, L.; Rastogi, R.; Du, W.; Hüttemann, M.; Fite, A.; Franchi, L. STAT3 tyrosine phosphorylation is critical for interleukin 1 beta and interleukin-6 production in response to lipopolysaccharide and live bacteria. Mol. Immunol. 2009, 46, 1867–1877. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Hokari, R.; Miura, S.; Shigematsu, T.; Hirokawa, M.; Akiba, Y.; Kurose, I.; Higuchi, H.; Fujimori, H.; Tsuzuki, Y. Increased expression of an inducible isoform of nitric oxide synthase and the formation of peroxynitrite in colonic mucosa of patients with active ulcerative colitis. Gut 1998, 42, 180–187. [Google Scholar] [PubMed]
- Osawa, T. Novel natural antioxidants for utilization in food and biological systems. In Postharvest Biochemistry of Plant Food-Materials in the Tropics; Uritani, I., Garcia, V.V., Mendoza, E.M., Eds.; Japan Scientific Societies Press: Tokyo, Japan, 1994; pp. 241–251. [Google Scholar]
- Jones, B.W.; Means, T.K.; Heldwein, K.A.; Keen, M.A.; Hill, P.J.; Belisle, J.T.; Fenton, M.J. Different Toll-like receptor agonists induce distinct macrophage responses. J. Leukoc. Biol. 2001, 69, 1036–1044. [Google Scholar] [PubMed]
- Rhee, S.H.; Hwang, D. Murine Toll-like receptor 4 confers lipopolysaccharide responsiveness as determined by activation of NFκB and expression of the inducible cyclooxygenase. J. Biol. Chem. 2000, 275, 34035–34040. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.J.; Moon, J.; Song, G.; Lee, Y.; Han, M.; Lee, J.; Ihm, B.; Lee, W.; Lee, N.; Hyun, C. Artemisia fukudo essential oil attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.B.; Reddanna, P. Chebulagic acid (CA) attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 381, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Bae, D.S.; Pan, C.H.; Samdan, J.; Lee, J.K. Protopine reduces the inflammatory activity of lipopolysaccharide-stimulated murine macrophages. BMB Rep. 2012, 45, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Byun, M.W. Anti-inflammatory activity of austroinulin from Stevia rebaudiana in LPS-induced RAW264. 7 cells. J. Korean Soc. Food Sci. Nutr. 2012, 41, 456–461. [Google Scholar] [CrossRef]
- Kim, K.N.; Heo, S.J.; Yoon, W.J.; Kang, S.M.; Ahn, G.; Yi, T.H.; Jeon, Y.J. Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. Eur. J. Pharmacol. 2010, 649, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Maity, B.; Yadav, S.K.; Patro, B.S.; Tyagi, M.; Bandyopadhyay, S.K.; Chattopadhyay, S. Molecular mechanism of the anti-inflammatory activity of a natural diarylnonanoid, malabaricone C. Free Radic. Biol. Med. 2012, 52, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.S.; Kim, W.J.; Kim, J.J.; Kim, T.J.; Ye, S.K.; Song, M.D.; Kang, H.; Kim, D.W.; Moon, W.K.; Lee, K.H. Gold nanoparticles attenuate LPS-induced NO production through the inhibition of NF-κB and IFN-β/STAT1 pathways in RAW264. 7 cells. Nitric Oxide 2010, 23, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Aline, M.; Charles, E.L.; Marco, R.; Jeanne, M.; Odile, G.N. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar]
- Quettier-Deleu, C.; Gressier, B.; Vasseur, J.; Dine, T.; Brunet, C.; Luyckx, M.; Cazin, M.; Cazin, J.C.; Bailleul, F.; Trotin, F. Phenolic compounds and antioxidant activities of buckwheat (Fagopyrum esculentum Moench) hulls and flour. J. Ethnopharmacol. 2000, 72, 35–42. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Lee, S.J.; Lim, B.O.; Jeon, Y.J.; Song, M.D.; Park, T.K.; Lee, K.H.; Kim, B.; Lee, S.R.; Moon, S.H.; et al. Antioxidative and neuroprotective effects of enzymatic extracts from leaves of Perilla frutescens var. japonica. Food Sci. Biotechnol. 2008, 17, 279–286. [Google Scholar]
- Hiramoto, K.; Johkoh, H.; Sako, K.I.; Kikugawa, K. DNA breaking activity of the carbon-centered radical generated from 2,2-azobis-(2-amidinopropane)-hydrochloride (AAPH). Free Radic. Res. Commun. 1993, 19, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Rosen, G.M.; Rauckman, E.J. Spin trapping of superoxide and hydroxyl radicals. Methods Enzymol. 1984, 105, 198–205. [Google Scholar] [PubMed]
- Bass, D.; Parce, J.W.; Dechatelet, L.R.; Szejda, P.; Seeds, M.; Thomas, M. Flow cytometric studies of oxidative product formation by neutrophils: A graded response to membrane stimulation. J. Immunol. 1983, 130, 1910–1917. [Google Scholar] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Extraction Yield (%) | Total Polyphenol (mg GAE/g Extract) | Total Flavonoid (mg CE/g Extract) | ABTS (mM Trolox eq./mg Extract) | FRAP (mM FeSO4 eq./mg Extract) |
|---|---|---|---|---|---|
| Water extract | 28.36 | 165.9 ± 2.5 | 174.3 ± 3.2 | 0.99 ± 0.04 | 2.45 ± 0.08 |
| Ethanol extract | 26.7 | 132.4 ± 1.9 | 155 ± 4.2 | 0.94 ± 0.03 | 2.16 ± 0.07 |
| Sample | DPPH Radical | Alkyl Radical | Hydroxyl Radical |
|---|---|---|---|
| Water extract | 3.06 ± 0.12 | 0.85 ± 0.02 | 205.45 ± 5.98 |
| Ethanol extract | 5.75 ± 0.24 | 7.00 ± 0.36 | 328.58 ± 6.47 |
| Vitamin C | 4.79 ± 0.25 | 5.70 ± 0.32 | 42.41 ± 2.31 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.-K.; Kim, Y.-S.; Natarajan, S.B.; Kim, W.-S.; Hwang, J.-W.; Jeon, N.-J.; Jeong, J.-H.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Antioxidant and Anti-Inflammatory Effects of Chaenomeles sinensis Leaf Extracts on LPS-Stimulated RAW 264.7 Cells. Molecules 2016, 21, 422. https://doi.org/10.3390/molecules21040422
Han Y-K, Kim Y-S, Natarajan SB, Kim W-S, Hwang J-W, Jeon N-J, Jeong J-H, Moon S-H, Jeon B-T, Park P-J. Antioxidant and Anti-Inflammatory Effects of Chaenomeles sinensis Leaf Extracts on LPS-Stimulated RAW 264.7 Cells. Molecules. 2016; 21(4):422. https://doi.org/10.3390/molecules21040422
Chicago/Turabian StyleHan, Young-Ki, Yon-Suk Kim, Sithranga Boopathy Natarajan, Won-Suk Kim, Jin-Woo Hwang, Nam-Joo Jeon, Jae-Hyun Jeong, Sang-Ho Moon, Byong-Tae Jeon, and Pyo-Jam Park. 2016. "Antioxidant and Anti-Inflammatory Effects of Chaenomeles sinensis Leaf Extracts on LPS-Stimulated RAW 264.7 Cells" Molecules 21, no. 4: 422. https://doi.org/10.3390/molecules21040422
APA StyleHan, Y. -K., Kim, Y. -S., Natarajan, S. B., Kim, W. -S., Hwang, J. -W., Jeon, N. -J., Jeong, J. -H., Moon, S. -H., Jeon, B. -T., & Park, P. -J. (2016). Antioxidant and Anti-Inflammatory Effects of Chaenomeles sinensis Leaf Extracts on LPS-Stimulated RAW 264.7 Cells. Molecules, 21(4), 422. https://doi.org/10.3390/molecules21040422