
The Effect of Phenazine-1-Carboxylic Acid on the Morphological, Physiological, and Molecular Characteristics of Phellinus noxius

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of PCA on Growth and Morphology of P. noxius Mycelium
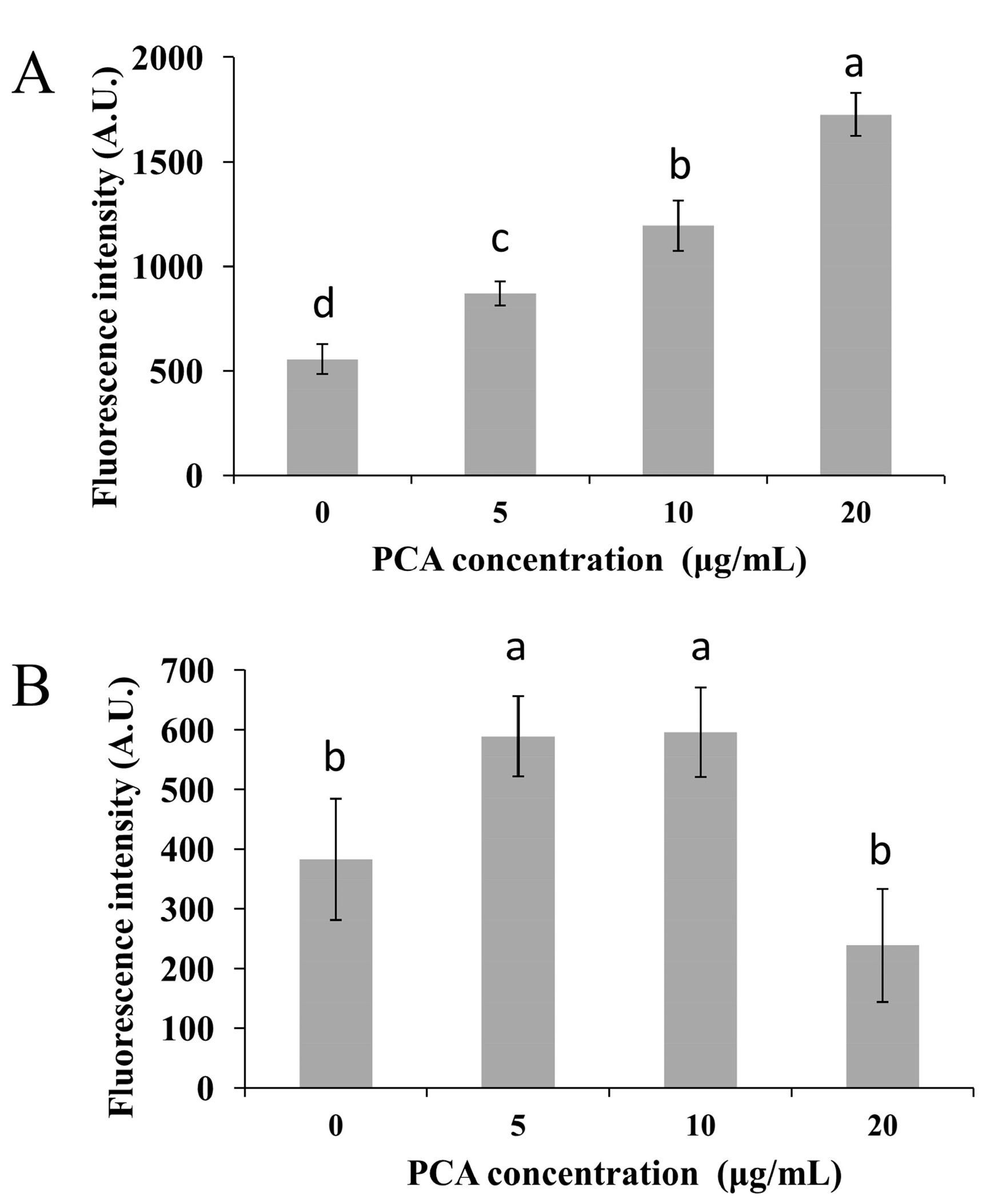
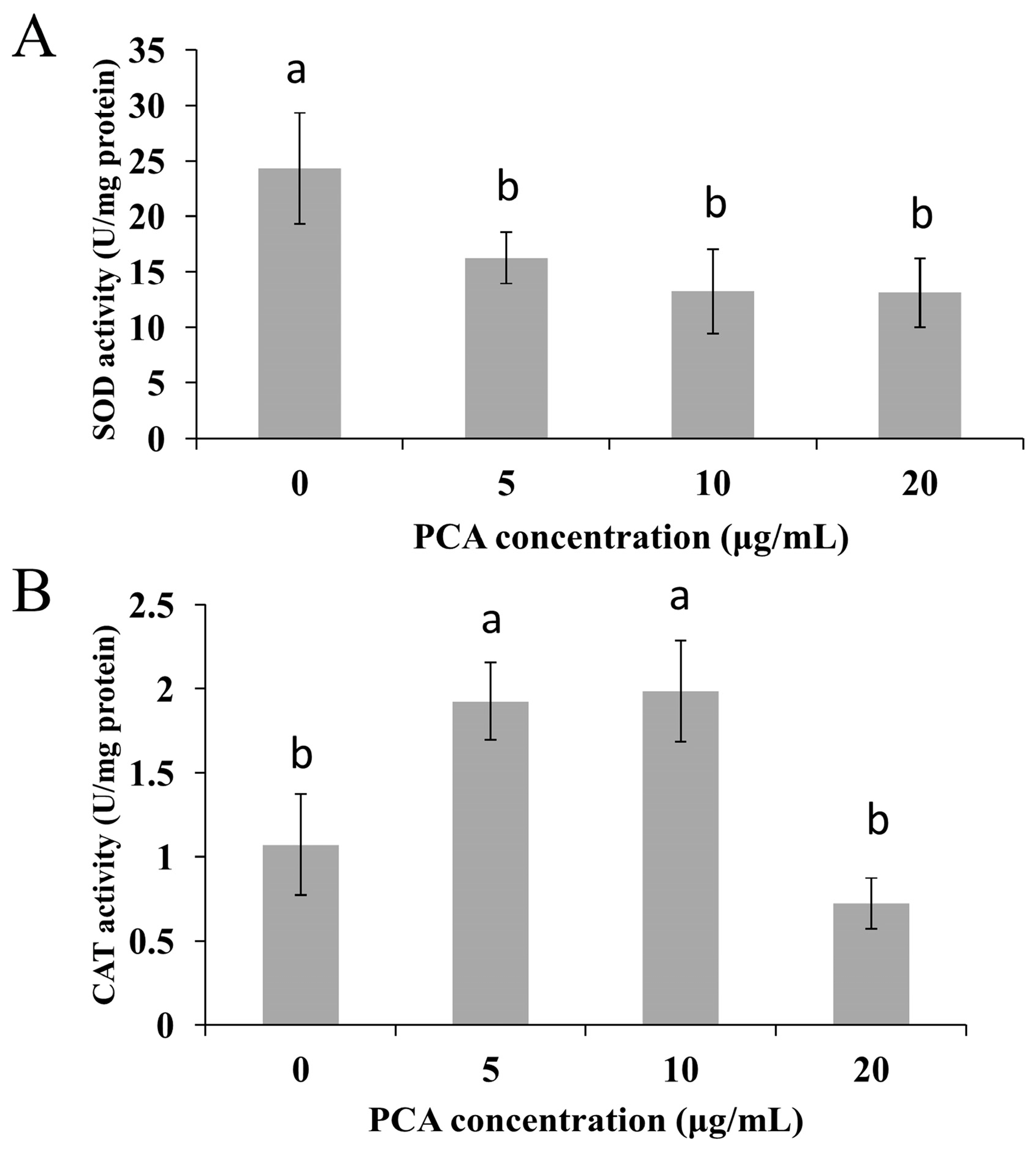
2.2. Mechanism of Action of PCA in P. noxius
2.3. Effects of PCA on Gene Expression of P. noxius
3. Materials and Methods
3.1. Fungal Strain, Culture Conditions, and Chemicals
3.2. Effects of Inhibition of PCA in P. noxius on PDA Plate and PDB Medium in Vitro
3.3. Effect of PCA on Intracellular Reactive Oxygen Species (ROS) Accumulation in P. noxius
3.4. Effect of PCA on Mitochondrial Membrane Potential (MMP)
3.5. Effects of PCA on Activities of Superoxide Dismutase (SOD) and Catalase (CAT) in P. noxius
3.6. Quantitative Real-Time PCR
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Akiba, M.; Ota, Y.; Tsai, I.J.; Hattori, T.; Sahashi, N.; Kikuchi, T. Genetic differentiation and spatial structure of Phellinus noxius, the causal agent of brown root rot of woody plants in Japan. PLoS ONE 2015, 10, e0141792. [Google Scholar]
- Sahashi, N.; Akiba, M.; Takemoto, S.; Yokoi, T.; Ota, Y.; Kanzaki, N. Phellinus noxius causes brown root rot on four important conifer species in Japan. Eur. J. Plant Pathol. 2014, 140, 869–873. [Google Scholar] [CrossRef]
- Ann, P.J.; Chang, T.T.; Ko, W.H. Phellinus noxius brown root rot of fruit and ornamental trees in Taiwan. Plant Dis. 2002, 86, 820–826. [Google Scholar] [CrossRef]
- Chung, C.L.; Huang, S.Y.; Huang, Y.C.; Tzean, S.S.; Ann, P.J.; Tsai, J.N.; Yang, C.C.; Lee, H.H.; Huang, T.W.; Huang, H.Y.; et al. The Genetic structure of Phellinus noxius and dissemination pattern of brown root rot disease in Taiwan. PLoS ONE 2015, 10, e0139445. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, F.W.; Jauss, F.; Spencer, C.; Hallam, C.; Schubert, M. Evaluation of an antagonistic Trichoderma strain for reducing the rate of wood decomposition by the white rot fungus Phellinus noxius. Biol. Control. 2012, 61, 160–168. [Google Scholar] [CrossRef]
- Chang, T.T. Survival of Phellinus noxius in soil and in the roots of dead host plants. Phytopathology 1996, 86, 272–276. [Google Scholar] [CrossRef]
- Chang, T.T.; Chang, R.J. Generation of volatile ammonia from urea fungicidal to Phellinus noxius in infested wood in soil under controlled conditions. Plant Pathol. 1999, 48, 337–344. [Google Scholar] [CrossRef]
- Fu, C.H.; Hu, B.Y.; Chang, T.T.; Hsueh, K.L.; Hsu, W.T. Evaluation of dazomet as fumigant for the control of brown root rot disease. Pest Manag. Sci. 2012, 68, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Puopolo, G.; Masi, M.; Raio, A.; Andolfi, A.; Zoina, A.; Cimmino, A.; Evidente, A. Insights on the susceptibility of plant pathogenic fungi to phenazine-1-carboxylic acid and its chemical derivatives. Nat. Prod. Res. 2013, 27, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Pan, X.; Luo, J.; Wu, J.; Zhou, Z.; Liang, X.; He, Y.; Zhou, M. Effects of phenazine-1-carboxylic acid on the biology of the plant-pathogenic bacterium Xanthomonas oryzae pv. oryzae. Pest Biochem. Physiol. 2015, 117, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Cezairliyan, B.; Vinayavekhin, N.; Grenfell-Lee, D.; Yuen, G.J.; Saghatelian, A.; Ausubel, F.M. Identification of Pseudomonas aeruginosa phenazines that kill Caenorhabditis elegans. PLoS Pathog. 2013, 9, e1003101. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.L.; Li, Y.Q.; Wang, Y.; Zhang, X.H.; Xu, Y.Q. Optimization of critical medium components using response surface methodology for phenazine-1 -carboxylic acid production by pseudomonas sp. M-18Q. J. Biosci. Bioeng. 2008, 105, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Bonsall, R.F.; Mavrodi, D.V.; Weller, D.M.; Thomashow, L.S. Transformation of Pseudomonas fluorescens with genes for biosynthesis of phenazine-1-carboxylic acid improves biocontrol of rhizoctonia root rot and in situ antibiotic production. FEMS Microbiol. Ecol. 2004, 49, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, L.S.; Weller, D.M.; Bonsall, R.F.; Pierson, L.S. Production of the antibiotic phenazine-1-carboxylic acid by fluorescent pseudomonas species in the rhizosphere of wheat. Appl. Environ. Microbiol. 1990, 56, 908–912. [Google Scholar] [PubMed]
- Raio, A.; Puopolo, G.; Cimmino, A.; Danti, R.; Della Rocca, G.; Evidente, A. Biocontrol of cypress canker by the phenazine producer Pseudomonas chlororaphis subsp. Aureofasciens strain M71. Biol. Control. 2011, 58, 133–138. [Google Scholar] [CrossRef]
- Abraham, A.; Philip, S.; Jacob, M.K.; Narayanan, S.P.; Jacob, C.K.; Kochupurackal, J. Phenazine-1-carboxylic acid mediated anti-oomycete activity of the endophytic Alcaligenes sp. EIL-2 against Phytophthora meadii. Microbiol. Res. 2015, 170, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Thid, D.; Benkoski, J.J.; Svedhem, S.; Kasemo, B.; Gold, J. DHA-induced changes of supported lipid membrane morphology. Langmuir 2007, 23, 5878–5881. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Cho, N.J. Spectrum of membrane morphological responses to antibacterial fatty acids and related surfactants. Langmuir 2015, 31, 10223–10232. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Baracca, A.; Sgarbi, G.; Solaini, G.; Lenaz, G. Rhodamine 123 as a probe of mitochondrial membrane potential: evaluation of proton flux through F0 during ATP synthesis. BBA-Bioenergetics 2003, 1606, 137–146. [Google Scholar] [CrossRef]
- Tupe, S.G.; Kulkarni, R.R.; Shirazi, F.; Sant, D.G.; Joshi, S.P.; Deshpande, M.V. Possible mechanism of antifungal phenazine-1-carboxamide from Pseudomonas sp. against dimorphic fungi Benjaminiella poitrasii and human pathogen Candida albicans. J. Appl. Microbiol. 2015, 118, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Yano, T.; Kassovska-Bratinova, S.; The, J.S.; Winkler, J.; Sullivan, K.; Isaacs, A.; Schechter, N.M.; Rubin, H. Reduction of clofazimine by mycobacterial type 2 NADH:quinone oxidoreductase: A pathway for the generation of bactericidal levels of reactive oxygen species. J. Biol. Chem. 2011, 286, 10276–10287. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M.; Fridowich, I. Mechanism of antibiotic action of pyocyanin. J. Bacteriol. 1980, 141, 156–163. [Google Scholar] [PubMed]
- Muller, M. Pyocyanin induces oxidative stress in human endothelial cells and modulates the glutathione redox cycle. Free Radic. Biol. Med. 2002, 33, 1527–1533. [Google Scholar] [CrossRef]
- Morales, D.K.; Jacobs, N.J.; Rajamani, S.; Krishnamurthy, M.; Cubillos-Ruiz, J.R.; Hogan, D.A. Antifungal mechanisms by which a novel Pseudomonas aeruginosa phenazine toxin kills Candida albicans in biofilms. Mol. Microbiol. 2010, 78, 1379–1392. [Google Scholar] [CrossRef] [PubMed]
- Del Sorbo, G.; Schoonbeek, H.; De Waard, M.A. Fungal transporters involved in efflux of natural toxic compounds and fungicides. Fungal Genet. Biol. 2000, 30, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cervelatti, E.P.; Fachin, A.L.; Ferreira-Nozawa, M.S.; Martinez-Rossi, N.M. Molecular cloning and characterization of a novel ABC transporter gene in the human pathogen Trichophyton rubrum. Med. Mycol. 2006, 44, 141–147. [Google Scholar] [CrossRef]
- Li, D.; Alic, M.; Brown, J.A.; Gold, M.H. Regulation of manganese peroxidase gene transcription by hydrogen peroxide, chemical stress, and molecular oxygen. Appl. Environ. Microbiol. 1995, 61, 341–345. [Google Scholar] [PubMed]
- Kawasaki, L.; Aguirre, J. Multiple catalase genes are differentially regulated in Aspergillus nidulans. J. Bacteriol. 2001, 183, 1434–1440. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Klotz, M.G.; York, J.L.; Kruft, V.; McEwen, J.E. Redundancy, phylogeny and differential expression of Histoplasma capsulatum catalases. Microbiology 2002, 148, 1129–1142. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.K.; Grahl, N.; Okegbe, C.; Dietrich, L.E.; Jacobs, N.J.; Hogan, D.A. Control of Candida albicans metabolism and biofilm formation by Pseudomonas aeruginosa phenazines. mBio 2013, 4, e00526–e00512. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Jiang, H.X.; Sun, S.; Yang, D.D.; Jin, K.M.; Zhang, W.; He, Y.W. Biotechnological potential of a rhizosphere Pseudomonas aeruginosa strain producing phenazine-1-carboxylic acid and phenazine-1-carboxamide. World J. Microbiol. Biotechnol. 2016, 32. [Google Scholar] [CrossRef] [PubMed]
- Churchill, A.; Ciuffetti, L.; Hansen, D.; van Etten, H.; van Alfen, N. Transformation of the fungal pathogen Cryphonectria parasitica with a variety of heterologous plasmids. Curr. Genet. 1990, 17, 25–31. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chung, R.S.; Chen, C.C.; Ng, L.T. Nitrogen fertilization affects the growth performance, betaine and polysaccharide concentrations of Lycium barbarum. Ind. Crops Prod. 2010, 32, 650–655. [Google Scholar] [CrossRef]
- Sample Availability: Not Available.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Putative Function | Sequence (5′–3′) 1 | Amplicon (bp.) |
|---|---|---|---|
| MRP4 | multidrug resistance protein 4 | GGGTTTGACCTCTATCCGAA (F) | 105 |
| CCAGCAACATCGACCAATAC (R) | |||
| ABC1 | ATP-binding cassette protein | AGTGCTCGTGAAATGAAACG (F) | 96 |
| ACTCCGAATGGTGGCTAATC (R) | |||
| MFS1 | MFS transporter | CTCCGGTTCAACATCACATC (F) | 105 |
| AAGGGTCATCTGGTCCATTC (R) | |||
| CAT1 | catalase | CCTTACGATTCTCCACCGTT (F) | 107 |
| CCAATCAAGATTTCCGTCCT (R) | |||
| MNP1 | manganese peroxidase 1 | TCTACGATCTTGCTGATGCC (F) | 91 |
| TCGTGGAAAGTGAGACGAAG (R) | |||
| HEME1 | heme peroxidase | ATCGGATCAAATGGGTTCAT (F) | 98 |
| GGCCATTTCATGGCTCTTAT (R) | |||
| COX1 | cytochrome c oxidase subunit 1 | TTTAACCTTGGTTTCACTGATGA (F) | 85 |
| GGTCACTGATAGGTGGCTGA (R) | |||
| EF1α | elongation factor 1-alpha | ACGGCGGATATCCTTAACTG (F) | 107 |
| CTGAAGTGAAGTCCGTCGAA (R) |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Sun, L.; Bi, K.; Zhong, G.; Hu, M. The Effect of Phenazine-1-Carboxylic Acid on the Morphological, Physiological, and Molecular Characteristics of Phellinus noxius. Molecules 2016, 21, 613. https://doi.org/10.3390/molecules21050613
Huang H, Sun L, Bi K, Zhong G, Hu M. The Effect of Phenazine-1-Carboxylic Acid on the Morphological, Physiological, and Molecular Characteristics of Phellinus noxius. Molecules. 2016; 21(5):613. https://doi.org/10.3390/molecules21050613
Chicago/Turabian StyleHuang, Huazhi, Longhua Sun, Keke Bi, Guohua Zhong, and Meiying Hu. 2016. "The Effect of Phenazine-1-Carboxylic Acid on the Morphological, Physiological, and Molecular Characteristics of Phellinus noxius" Molecules 21, no. 5: 613. https://doi.org/10.3390/molecules21050613
APA StyleHuang, H., Sun, L., Bi, K., Zhong, G., & Hu, M. (2016). The Effect of Phenazine-1-Carboxylic Acid on the Morphological, Physiological, and Molecular Characteristics of Phellinus noxius. Molecules, 21(5), 613. https://doi.org/10.3390/molecules21050613