
Phytoestrogen Metabolism by Adult Human Gut Microbiota


Abstract
:1. Introduction
2. Results and Discussion
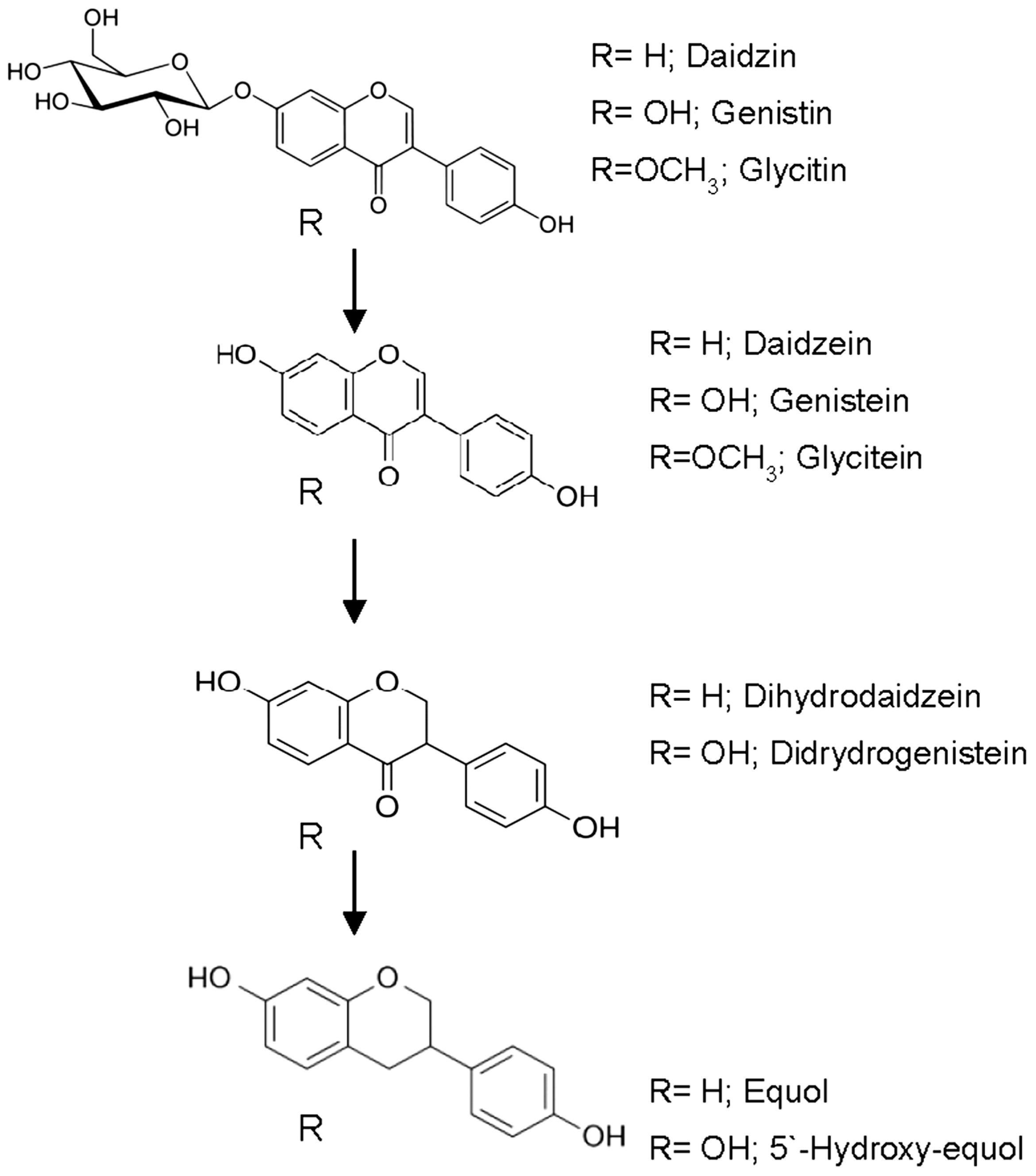
2.1. Metabolism of Isoflavones
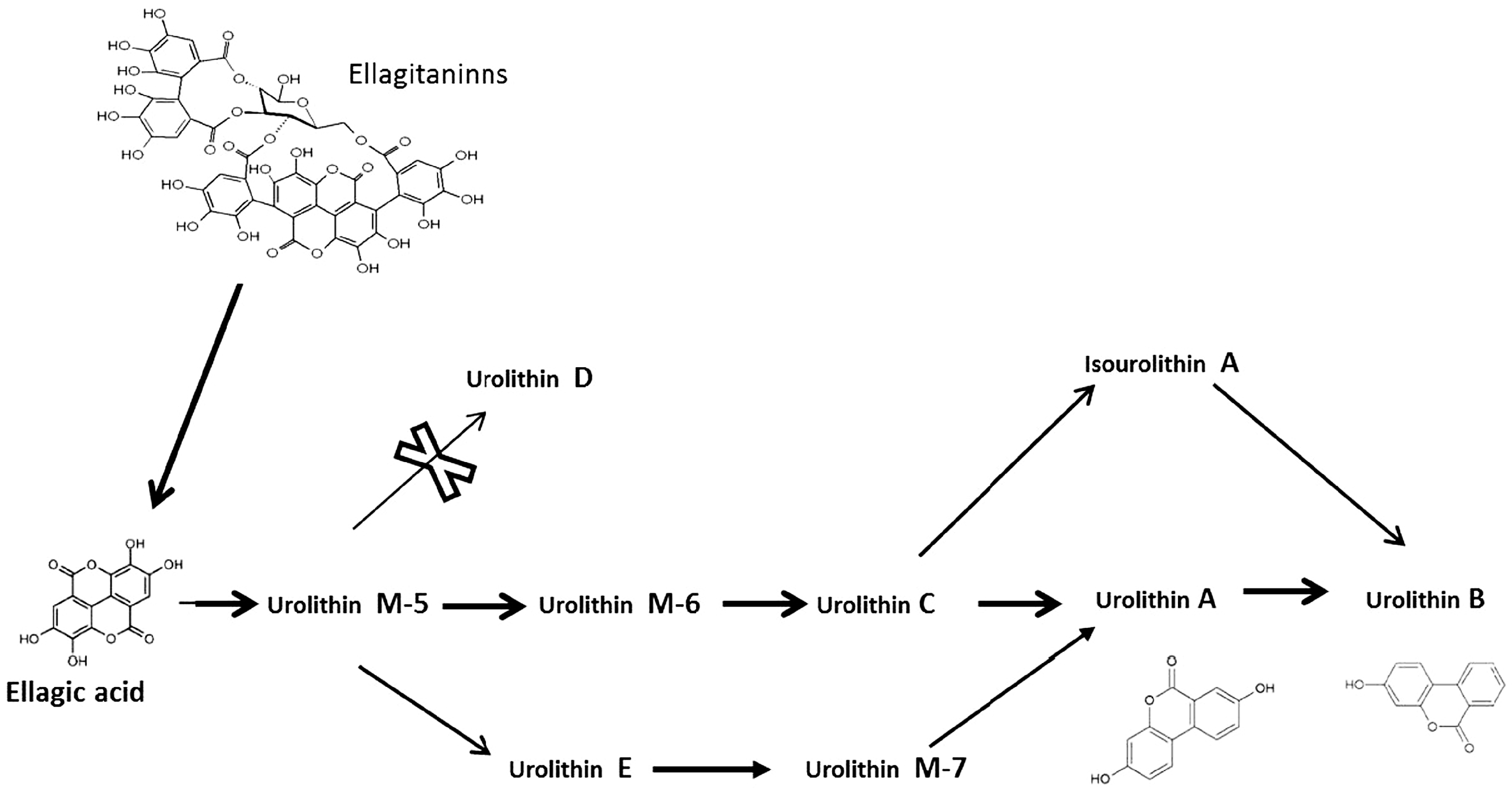
2.2. Metabolism of Ellagitannins
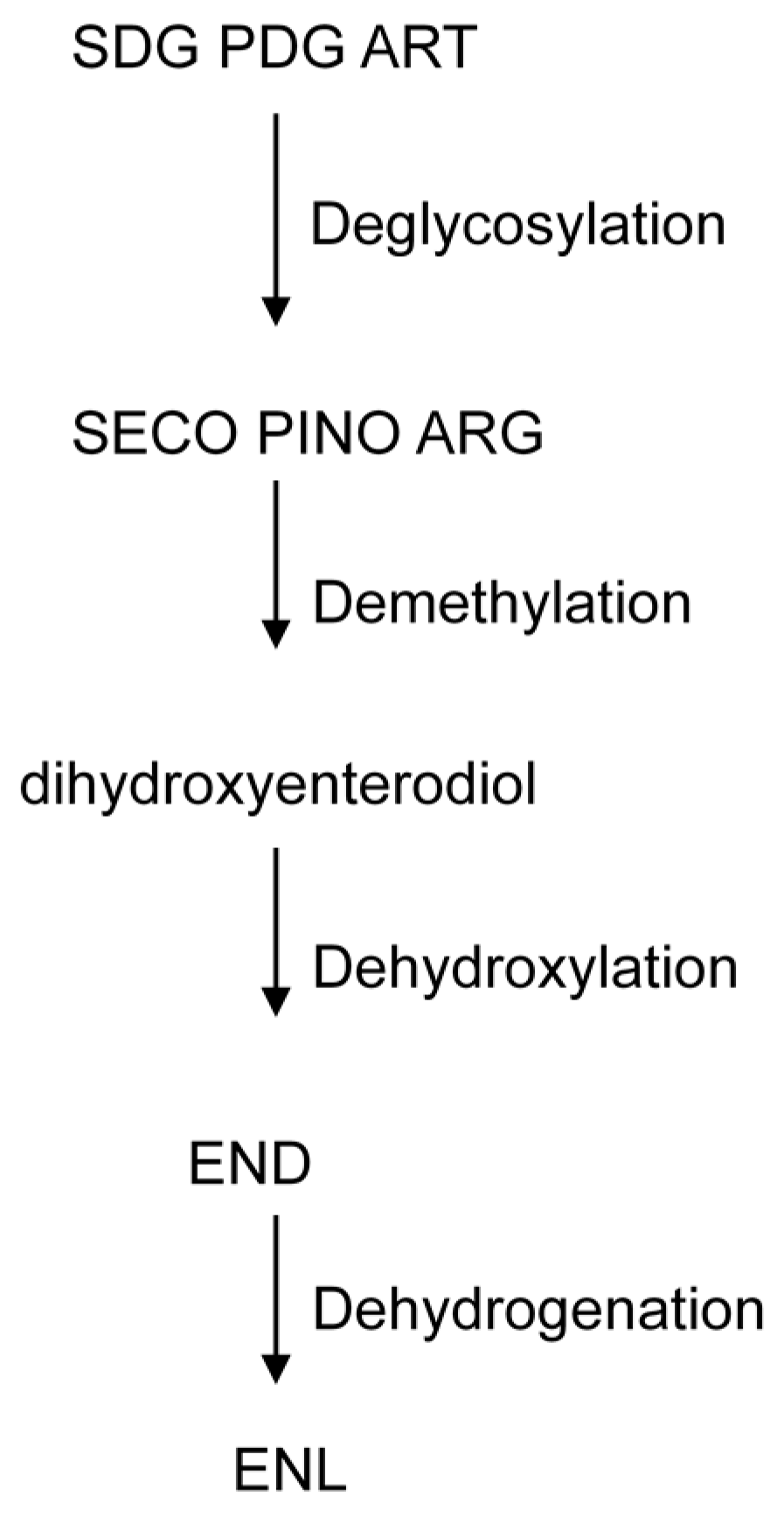
2.3. Metabolism of Lignans
2.4. Correlation between Pairs of Groups of Metabolites
3. Experimental Section
3.1. Chemicals and Solvents
3.2. Collection of Human Fecal Samples
3.3. Phytoestrogen Fermentation Assay
3.4. Extraction of Phytoestrogens
3.5. HPLC-PAD and HPLC-ESI/MS Analysis
3.6. Identification and Quantification of Phenolic Compounds
3.7. Determination of β-Glucosidase and β-Glucuronidase Activities
3.8. Correlation Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, H.; Murphy, P.A. Isoflavone content in commercial soybean foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar]
- Landete, J.M. Ellagitannins, ellagic acid and their derived metabolites: A review about source, metabolism, function and health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar]
- Landete, J.M. Plant and mammalian lignans: A review of source, intake, metabolism, intestinal bacteria and health. Food Res. Int. 2012, 46, 410–424. [Google Scholar] [CrossRef]
- Harris, D.M.; Besselink, E.; Henning, S.M.; Go, V.L.W.; Heber, D. Phytoestrogens induce differential estrogen receptor alpha- or beta-mediated responses in transfected breast cancer cells. Exp. Biol. Med. 2005, 230, 558–568. [Google Scholar]
- Landete, J.M.; Arqués, J.L.; Medina, M.; Gaya, P.; de las Rivas, B.; Muñoz, R. Bioactivation of phytoestrogens: Intestinal bacteria and health. Crit. Rev. Food Sci. Nutr. 2015, 25. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Morand, C.; Manach, C.; Rémesy, C. Absorption and metabolism of polyphenols in the gut and impact on health. Biomed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef]
- Seeram, N.P.; Aronson, W.J.; Zhang, Y.; Henning, S.M.; Moro, A.; Lee, R.P.; Sartippour, M.; Harris, D.M.; Rettig, M.; Suchard, M.A.; et al. Pomegranate ellagitannin-derived metabolites inhibit prostate cancer growth and localize to the mouse prostate gland. J. Agric. Food Chem. 2007, 55, 7732–7737. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-H.; Larn, T.-H.; Lau, K.-K.; Yiu, K.-H.; Siu, C.-W.; Li, S.-W.; Chan, H.-T.; Tam, H.-T.; Lau, C.-P.; Tse, H.-F. Dietary intake of phytoestrogen is associated with increased circulating endothelial progenitor cells in patients with cardiovascular disease. Eur. J. Prev. Cardiol. 2011, 18, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Yeo, Y.; Ko, K.-P.; Ma, S.-H.; Yang, J.J.; Shin, A.; Park, S.K.; Chang, S.-H.; Shin, H.-R.; Kang, D.; Yoo, K.-Y. Isoflavones from phytoestrogens and colorectal cancer risk: A nested case-control study within the Korean multicenter cancer cohort. Cancer Res. 2013, 73, 4823. [Google Scholar] [CrossRef]
- Poluzzi, E.; Piccinni, C.; Raschi, E.; Rampa, A.; Recanatini, M.; de Ponti, F. Phytoestrogens in postmenopause: The state of the art from a chemical, pharmacological and regulatory perspective. Curr. Med. Chem. 2014, 21, 417–436. [Google Scholar] [PubMed]
- Chen, M.-N.; Lin, C.-C.; Liu, C.-F. Efficacy of phytoestrogens for menopausal symptoms: A meta-analysis and systematic review. Climacteric 2015, 18, 260–269. [Google Scholar] [PubMed]
- Edel, A.L.; Pierce, G.N.; Aliani, M. Age-dependency in the metabolism of flaxseed lignans by healthy adults. J. Funct. Foods 2015, 17, 948–957. [Google Scholar] [CrossRef]
- Pierce, G.N.; Aliani, M. The effect of flaxseed dose on circulating concentrations of alpha-linolenic acid and secoisolariciresinol diglucoside derived enterolignans in young healthy adults. Eur. J. Nutr. 2016, 55, 651–663. [Google Scholar]
- Setchell, K.D.R.; Brown, N.M.; Lydeking-Olsen, E. The clinical importance of the metabolite equol—A clue to the effectiveness of soy and its isoflavones. J. Nutr. 2002, 132, 3577–3584. [Google Scholar] [PubMed]
- Setchell, K.D.R.; Brown, N.M.; Zimmer-Nechemias, L.; Brashear, W.T.; Wolfe, B.E.; Kirschner, A.S.; Heubi, J.E. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. Am. J. Clin. Nutr. 2002, 76, 447–453. [Google Scholar] [PubMed]
- Guadamuro, L.; Delgado, S.; Redruello, B.; Flórez, A.B.; Suárez, A.; Martínez-Camblor, P.; Mayo, B. Equol status and changes in fecal microbiota in menopausal women receiving long-term treatment for menopause symptoms with a soy-isoflavone concentrate. Front. Microbiol. 2015, 6, 777. [Google Scholar] [PubMed]
- Zoetendal, E.G.; Antoon, D.L.; Akkermans, A.D.L.; de Vos, W.M. Temperature gradient gel electrophoresis analysis of 16S rRNA from human fecal hamples reveals stable and host-specific communities of active bacteria. J. Appl. Environ. Microbiol. 1998, 64, 3854–3859. [Google Scholar]
- Kemperman, R.A.; Bolca, S.; Loger, L.C.; Vaughan, E.E. Novel approaches for analyzing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Harris, K.S.; Wang, H.J.; Murphy, P.A.; Hendrich, S. Bioavailability of soybean isoflavones depends upon gut microflora in women. J. Nutr. 1995, 125, 2307–2315. [Google Scholar] [PubMed]
- Day, A.J.; DuPont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.; Morgan, M.R.; Williamson, G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver beta-glucosidase activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef]
- Gaya, P.; Peirotén, A.; Medina, M.; Landete, J.M. Isoflavone metabolism by a collection of lactic acid bacteria and bifidobacteria with biotechnological interest. Int. J. Food Sci. Nutr. 2016, 67, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [PubMed]
- Crespy, V.; Aprikian, O.; Morand, C.; Besson, C.; Manach, C.; Demigné, C.; Rémésy, C. Bioavailability of phloretin and phloridzin in rats. J. Nutr. 2001, 131, 3227–3230. [Google Scholar] [PubMed]
- Donova, J.L.; Crespy, V.; Manach, C.; Morand, C.; Besson, C.; Scalbert, A.; Rémésy, C. Catechin is metabolized by both the small intestine and liver of rats. J. Nutr. 2001, 131, 1753–1757. [Google Scholar]
- Mcbain, A.J.; Macfarlane, G.T. Ecological and physiological studies on large intestinal bacteria in relation to production of hydrolytic and reductive enzymes involved in formation of genotoxic metabolites. J. Med. Microbiol. 1998, 47, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Faughnan, M.; Hoey, L.; Wähälä, K.; Williamson, G.; Cassidy, A. Bioavailability of phyto-estrogens. Br. J. Nutr. 2003, 89, S45–S58. [Google Scholar] [PubMed]
- Setchell, K.D.R.; Cassidy, A. Dietary isoflavones: Biological effects and relevance to human health. J. Nutr. 1999, 129, 758S–767S. [Google Scholar] [PubMed]
- Decroos, K.; Vanhemmens, S.; Cattoir, S.; Boon, N.; Verstraete, W. Isolation and characterisation of an equol-producing mixed microbial culture from a human fecal sample and its activity under gastrointestinal conditions. Arch. Microbiol. 2005, 183, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Zubik, L.; Meydani, M. Bioavailability of soybean isoflavones from aglycone and glucoside forms in American women. Am. J. Clin. Nutr. 2003, 77, 1459–1465. [Google Scholar] [PubMed]
- Hur, H.G.; Lay, J.O., Jr.; Beger, R.D.; Freeman, J.P.; Rafii, F. Isolation of human intestinal bacteria metabolizing the natural isoflavone glycosides daidzin and genistin. Arch. Microbiol. 2000, 174, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, M.; Obata, A. β-Glucosidases from soybean hydrolyze daidzin and genistin. J. Food Sci. 1993, 58, 144–147. [Google Scholar] [CrossRef]
- Delmonte, P.; Perry, J.; Rader, J.I. Determination of isoflavones in dietary supplements containing soy, Red Clover and kudzu: Extraction followed by basic or acid hydrolysis. J. Chrom. A 2006, 1107, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Barnes, S.; Coward, L.; Kirk, M.; Sfakianos, J. HPLC-mass spectrometry analysis of isoflavones. Proc. Soc. Exp. Biol. Med. 1998, 217, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Song, K.B.; Atkinson, C.; Frankenfeld, C.L.; Jokela, T.; Wähälä, K.; Thomas, W.K.; Lampe, J.W. Prevalence of daidzein-metabolizing phenotypes differs between Caucasian and Korean American women and girls. J. Nutr. 2006, 136, 1347–1351. [Google Scholar] [PubMed]
- Nylund, L.; Kaukinen, K.; Lindfors, K. The microbiota as a component of the celiac disease andnon-celiac gluten sensitivity. Clin. Nutr. Exper. 2016, 6, 17–24. [Google Scholar] [CrossRef]
- Adlercreutz, H.; Musey, P.I.; Fotsis, T.; Bannwart, C.; Wahala, K.; Makela, T.; Brunow, G.; Hase, T. Identification of lignans and phytoestrogens in urine of chimpanzees. Clin. Chim. Acta 1986, 158, 147–154. [Google Scholar] [CrossRef]
- Lamartiniere, C.A.; Wang, J.; Smith-Johnson, M.; Eltoum, I.E. Daidzein: Bioavailability, potential for reproductive toxicity, and breast cancer chemoprevention in female rats. Toxicol. Sci. 2002, 65, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Bowey, E.; Adlercreutz, H.; Rowland, I. Metabolism of isoflavones and lignans by the gut microflora: A study in germ-free and human flora associated rats. Food Chem. Toxicol. 2003, 41, 631–636. [Google Scholar] [CrossRef]
- Uchiyama, S.; Ueno, T.; Suzuki, T. Identification of a newly isolated equol-producing lactic acid bacterium from the human feces. J. Intest. Microbiol. 2007, 21, 217–220. [Google Scholar]
- Niwa, T.; Yokoyama, S.; Matsugasaki, N.; Inomata, E.; Taira, A.; Osawa, T. Stereochemical determination of O-desmethylangolensin produced from daidzein. Food Chem. 2015, 171, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Clerici, C.; Lephart, E.D.; Cole, S.J.; Heenan, C.; Castellani, D.; Wolfe, B.E.; Nechemias-Zimmer, L.; Brown, N.M.; Lund, T.D.; et al. S-Equol, a potent ligand for estrogen receptor, is the exclusive enantiomeric form of the soy isoflavone metabolite produced by human intestinal bacterial flora. Am. J. Clin. Nutr. 2005, 81, 1072–1079. [Google Scholar] [PubMed]
- Mahmoud, A.M.; Yang, W.; Bosland, M.C. Soy isoflavones and prostate cancer: A review of molecular mechanisms. J. Steroid Biochem. Mol. Biol. 2014, 140, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Rüfer, C.E.; Kulling, S.E. Antioxidant activity of isoflavones and their major metabolites using different in vitro assays. J. Agric. Food Chem. 2006, 54, 2926–2931. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Nair, M.G.; Strasburg, M. Antioxidant activities of isoflavones and their biological metabolites in a liposomal system. Arch. Biochem. Biophys. 1998, 356, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Matthies, A.; Clavel, T.; Gütschow, M.; Engst, W.; Haller, D.; Blaut, M.; Braune, A. Conversion of daidzein and genistein by an anaerobic bacterium newly isolated from the mouse intestine. Appl. Environ. Microbiol. 2008, 74, 4847–4852. [Google Scholar] [CrossRef] [PubMed]
- Schöefer, L.; Mohan, R.; Braune, A.; Birringer, M.; Blaut, M. Anaerobic C-ring cleavage of genistein and daidzein by Eubacterium ramulus. FEMS Microbiol. Lett. 2002, 208, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Mertens-Talcott, S.; Jilma-Stohlawetz, P.; Rios, J.; Hingorani, L.; Derendorf, H. Absorption, metabolism and antioxidant effects of pomegranate polyphenols after ingestion of a standardized extract in healthy human volunteers. J. Agric. Food Chem. 2006, 54, 8956–8961. [Google Scholar] [CrossRef] [PubMed]
- Borges, G.; Roowi, S.; Rouanet, J.-M.; Duthie, G.G.; Lean, M.E.J.; Crozier, A. The bioavailability of raspberry anthocyanins and ellagitannins in rats. Mol. Nutr. Food Res. 2007, 51, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Espin, J.C.; Tomás-Barberan, F.A. Interaction between phenolics and gut microbiota role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Espín, J.C.; Parra, S.; Martínez, P.; Tomás-Barberán, F.A. The potent in vitro antioxidant ellagitannins from pomegranate juice are metabolized into bioavailable but poor antioxidant hydroxy-6H-dibenzopyran-6-one derivatives by the colonic microflora in healthy humans. Eur. J. Nutr. 2004, 43, 205–220. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, D.; Beltrán, D.; Espín, J.C.; Selma, M.V.; Tomás-Barberán, F.A. Time course production of urolithins from ellagic acid by human gut microbiota. J. Agric. Food Chem. 2013, 61, 8797–8806. [Google Scholar] [CrossRef] [PubMed]
- González-Barrio, R.; Borges, G.; Mullen, W.; Crozier, A. Bioavailability of anthocyanins and ellagitannins following consumption of raspberries by healthy humans and subjects with an ileostomy. J. Agric. Food Chem. 2010, 58, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Romo-Vaquero, M.; García-Villalba, R.; González-Sarrias, A.; Beltran, D.; Tomás-Barberan, F.A.; Espin, J.C.; Selma, M.V. Interindividual variability in the human metabolism of ellagic acid: Contribution of Gordonibacter to urolithin production. J. Funct. Foods 2015, 17, 785–791. [Google Scholar] [CrossRef]
- Mazur, W.; Uehara, M.; Wähälä, K.; Adlercreutz, H. Phytooestrogen content of berries, and plasma concentrations and urinary excretion of enterolactone after a single strawberry-meal in human subjects. Br. J. Nutr. 2000, 83, 381–387. [Google Scholar] [PubMed]
- Clavel, T.; Lippman, R.; Gavini, F.; Dore, J.; Blaut, M. Clostridium saccharogumia sp. nov. and Lactonifactor longoviformis gen. nov., sp. nov., two novel human fecal bacteria involved in the conversion of the dietary phytoestrogen secoisolariciresinol diglucoside. Syst. Appl. Microbiol. 2007, 30, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Woting, A.; Clavel, T.; Loh, G.; Blaut, M. Bacterial transformation of dietary lignans in gnotobiotic rats. FEMS Microbiol. Ecol. 2010, 72, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.D.; Huang, K.S.; Wang, H.B.; Davin, L.B.; Lewis, N.G. Biosynthetic pathway to the cancer chemopreventive secoisolariciresinol diglucosidehydroxymethylglutaryl ester-linked lignan oligomers in flax seeds Linumusitatissimum). J. Nat. Prod. 2011, 64, 1388–1397. [Google Scholar] [CrossRef]
- Touré, A.; Xueming, X. Flaxseed lignans: Source, biosynthesis, metabolism, antioxidant activity, bio-active components, and health benefits. Compr. Rev. Food Sci. Food Saf. 2010, 9, 261–269. [Google Scholar] [CrossRef]
- Clavel, T.; Borrmann, D.; Braune, A.; Doré, J.; Blaut, M. Occurrence and activity of human intestinal bacteria involved in the conversion of dietary lignans. Anaerobe 2006, 12, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.H.; Akao, T.; Hamasaki, K.; Deyama, T.; Hattori, M. Biotransformation of pinoresinol diglucoside to mammalian lignans by human intestinal microflora, and isolation of Enterococcus fecalis strain PDG-1 responsible for the transformation of (+)-pinoresinol to (+)-lariciresinol. Chem. Pharm. Bull. 2003, 51, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Borriello, S.P.; Setchell, K.D.R.; Axelson, M.; Lawson, A.M. Production and metabolism of lignans by the human fecal flora. J. Appl. Bacteriol. 1985, 58, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Possemiers, S.; Bolca, S.; Eeckhaut, E.; Depypere, H.; Verstraete, W. Metabolism of isoflavones, lignans and prenylflavonoids by intestinal bacteria: Producer phenotyping and relation with intestinal community. FEMS Microbiol. Ecol. 2007, 61, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Knust, U.; Spiegelhalder, B.; Strowitzki, T.; Owen, R.W. Contribution of linseed intake to urine and serum enterolignan levels in German females: A randomized controlled intervention trial. Food Chem. Toxicol. 2006, 44, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Gaya, P.; Arqués, J.L.; Medina, M.; Landete, J.M. Analysis of bacterial metabolism of phytoestrogens by means of HPLC-PAD and HPLC-ESI/MS. Food Anal. Meth. 2015, 9, 537–547. [Google Scholar] [CrossRef]
- Berg, J.O.; Nord, C.E.; Wadstrom, T. Formation of glycosidases in batch and continuous cultures of Bacteroides fragilis. Appl. Environ. Microbiol. 1978, 35, 269–273. [Google Scholar] [PubMed]
- Sample Availability: not available.




{kind=link}
{kind=link}
{kind=link}
| Comp. | Control | H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | H10 | H11 | H12 | H13 | H14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Equol | nd | nd | nd | nd | nd | nd | 1.0 | nd | nd | nd | nd | nd | nd | nd | nd |
| Daidzein | 3158.2 | 1040.5 | 993.1 | 2348.1 | 2505.4 | 3076.4 | 1739.5 | 1873.5 | 1200.1 | 1081.6 | 2584.4 | 1564.1 | 1408.0 | 306.6 | 710.1 |
| Daidzin | 4437.9 | 39.2 | 445.4 | 95.3 | 31.3 | 39.7 | 50.3 | 127.6 | 68.1 | 42.3 | 43.6 | 190.0 | 355.8 | 244.2 | 220.4 |
| Genistin | 3032.2 | nd | 210.1 | nd | nd | nd | 20.6 | nd | 36.8 | nd | 45.8 | 110.8 | 81.4 | 44.6 | 33.3 |
| Genistein | 1877.7 | 2009.6 | 1522.4 | 4103.0 | 2249.0 | 4158.0 | 3241.0 | 1713.4 | 1615.7 | 1816.4 | 4144.6 | 2024.1 | 2561.2 | 917.3 | 1176.7 |
| Glycitein | 829.5 | 80.9 | 327.1 | 547.9 | 498.8 | 846.1 | 526.7 | 119.7 | 197.4 | 130.7 | 824.2 | 244.8 | 268.4 | 39.2 | 175.5 |
| Glycitin | 101.9 | 49.8 | 509.5 | 75.7 | 95.4 | 112.5 | 158.3 | 185.1 | 200.8 | 67.8 | 286.4 | 90.5 | 234.7 | 161.5 | 83.3 |
| DHD | 154.8 | 163.4 | 455.5 | 172.1 | 167.4 | 201.5 | 1812.8 | 134.0 | 273.6 | 190.9 | 295.2 | 136.8 | 146.6 | 152.1 | 358.3 |
| DHG | 23.6 | 25.4 | 71.2 | 49.0 | 63.8 | 52.7 | 21.7 | 29.1 | 128.0 | 35.7 | 103.2 | 408.0 | 21.3 | 315.7 | 78.2 |
| O-DMA | nd | nd | 1264.1 | 89.8 | 1373.1 | 1520.9 | 66.1 | 1164.6 | 109.6 | 108.9 | 135.3 | 854.9 | nd | 74.6 | 88.6 |
| Comp. | Control | H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | H10 | H11 | H12 | H13 | H14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Urolithin A | 0 | nd | nd | nd | nd | 0.105 | nd | nd | 1.739 | nd | nd | nd | nd | nd | nd |
| Urolithin B | 0 | nd | 33.472 | nd | 0.108 | 3.079 | 0.251 | 0.080 | 1.720 | nd | 20.180 | nd | nd | nd | nd |
| Urolithin C | 0 | nd | nd | nd | 14.072 | nd | nd | nd | nd | nd | 24.231 | nd | 68.440 | 14.792 | nd |
| Urolithin D | 0 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| Urolithin E | 0 | nd | 0.509 | 1.633 | 2.030 | 4.250 | 3.339 | 0.088 | 0.706 | 0.335 | nd | 1.277 | 0.2586 | 4.293 | 0.208 |
| Urolithin M-5 | 0 | nd | 15.064 | 0.214 | nd | 6.628 | 10.068 | 4.192 | 0.283 | 2.785 | nd | 3.243 | 5.69 | 3.988 | nd |
| Comp. | Control | H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | H10 | H11 | H12 | H13 | H14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enterodiol | nd | nd | 4.84 | nd | 14.74 | nd | 5.77 | nd | 0.56 | nd | 14.77 | nd | nd | nd | nd |
| DHend | nd | ↑↑ | ↑ | ↑ | ↑↑ | ↑ | ↑ | ↑ | ↑↑ | ↑↑ | ↑↑↑ | = | ↑↑ | ↑ | ↑ |
| Enterolactone | nd | 7.06 | 29.01 | 8.61 | 26.89 | 24.03 | 11.88 | 26.55 | 67.04 | 18.14 | 34.90 | 15.95 | 9.25 | 16.89 | 60.95 |
| SECO | 70.88 | 685.83 | 334.74 | 733.73 | 776.09 | 684.45 | 842.82 | 792.43 | 900.69 | 846.70 | 764.19 | 474.85 | 647.90 | 848.64 | 379.05 |
| Matairesinol | 0.92 | 23.15 | 25.56 | 14.72 | 29.06 | 26.68 | 17.72 | 35.44 | 30.07 | 26.09 | 15.90 | 14.47 | 23.12 | 37.29 | 37.15 |
| Pinoresinol | 0.02 | nd | 10.33 | 4.76 | 9.74 | 113.79 | 5.29 | 14.00 | 12.65 | 43.70 | 7.42 | 41.72 | 40.09 | 2.26 | 8.14 |
| Arctigenin | nd | nd | 5.93 | 0.81 | 4.31 | 2.94 | 1.84 | 1.28 | 5.37 | nd | 2.74 | 0.61 | 0.616 | 0.89 | 0.084 |
| Lariciresinol | nd | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | ↑↑ |
| SDG | nq | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ | ↓↓↓ |
| Factor | Eigenvalue | % Variance | Cumulative |
|---|---|---|---|
| 1 | 4.66 | 27.43 | 27.43 |
| 2 | 3.93 | 23.14 | 50.56 |
| 3 | 1.91 | 11.21 | 61.78 |
| 4 | 1.81 | 10.62 | 72.39 |
| 5 | 1.56 | 9.16 | 81.56 |
| 6 | 1.07 | 6.28 | 87.84 |
| 7 | 0.72 | 4.25 | 92.08 |
| 8 | 0.53 | 3.11 | 95.19 |
| 9 | 0.40 | 2.35 | 97.54 |
| 10 | 0.22 | 1.28 | 98.82 |
| 11 | 0.16 | 0.96 | 99.78 |
| 12 | 0.03 | 0.17 | 99.95 |
| 13 | 8.25 × 103 | 0.05 | 100.00 |
| 14 | 4.86 × 10−11 | 0.00 | 100.00 |
| 15 | 3.17 × 10−11 | 0.00 | 100.00 |
| 16 | 1.18 × 10−12 | 0.00 | 100.00 |
| 17 | 0.00 | 0.00 | 100.00 |
| Metabolites | Factor | Communalities | |||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| Daidzein | −0.23 | 0.82 | 0.13 | 0.43 | −0.04 | 0.16 | 0.95 |
| Daidzin | 0.83 | −0.39 | −0.12 | 0.03 | −0.02 | −0.08 | 0.87 |
| Genistin | 0.95 | −0.07 | 0.09 | −0.04 | 0.02 | −0.24 | 0.98 |
| Genistein | −0.16 | 0.93 | −0.12 | 0.13 | 0.10 | 0.15 | 0.95 |
| Glycitein | −0.01 | 0.88 | 0.20 | 0.28 | 0.16 | 0.07 | 0.92 |
| Glycitin | 0.83 | 0.05 | 0.41 | −0.06 | 0.18 | 0.13 | 0.91 |
| DHD | 0.04 | 0.16 | −0.02 | −0.28 | 0.82 | 0.17 | 0.80 |
| DHG | 0.10 | −0.15 | −0.03 | 0.00 | −0.08 | −0.97 | 0.97 |
| O-DMA | 0.24 | 0.11 | 0.28 | 0.81 | −0.02 | 0.01 | 0.80 |
| Urolithin B | 0.80 | 0.26 | 0.40 | −0.06 | 0.02 | 0.10 | 0.88 |
| Urolithin E | −0.27 | 0.07 | −0.05 | 0.45 | 0.67 | −0.41 | 0.91 |
| Urolithin M-5 | 0.70 | −0.05 | −0.08 | 0.28 | 0.61 | 0.14 | 0.98 |
| Enterolactone | −0.05 | −0.25 | 0.79 | −0.08 | −0.25 | 0.01 | 0.76 |
| SECO | −0.73 | 0.09 | 0.13 | −0.16 | 0.33 | 0.04 | 0.69 |
| Matairesinol | −0.18 | −0.82 | 0.35 | 0.27 | −0.03 | 0.14 | 0.92 |
| Pinoresinol | −0.05 | 0.23 | −0.18 | 0.81 | −0.05 | −0.02 | 0.74 |
| Arctigenin | 0.34 | 0.18 | 0.82 | 0.18 | 0.21 | 0.02 | 0.89 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaya, P.; Medina, M.; Sánchez-Jiménez, A.; Landete, J.M. Phytoestrogen Metabolism by Adult Human Gut Microbiota. Molecules 2016, 21, 1034. https://doi.org/10.3390/molecules21081034
Gaya P, Medina M, Sánchez-Jiménez A, Landete JM. Phytoestrogen Metabolism by Adult Human Gut Microbiota. Molecules. 2016; 21(8):1034. https://doi.org/10.3390/molecules21081034
Chicago/Turabian StyleGaya, Pilar, Margarita Medina, Abel Sánchez-Jiménez, and José Mᵃ Landete. 2016. "Phytoestrogen Metabolism by Adult Human Gut Microbiota" Molecules 21, no. 8: 1034. https://doi.org/10.3390/molecules21081034
APA StyleGaya, P., Medina, M., Sánchez-Jiménez, A., & Landete, J. M. (2016). Phytoestrogen Metabolism by Adult Human Gut Microbiota. Molecules, 21(8), 1034. https://doi.org/10.3390/molecules21081034