Molecular Cloning, Characterization, and Functional Analysis of Acetyl-CoA C-Acetyltransferase and Mevalonate Kinase Genes Involved in Terpene Trilactone Biosynthesis from Ginkgo biloba

Abstract
:1. Introduction
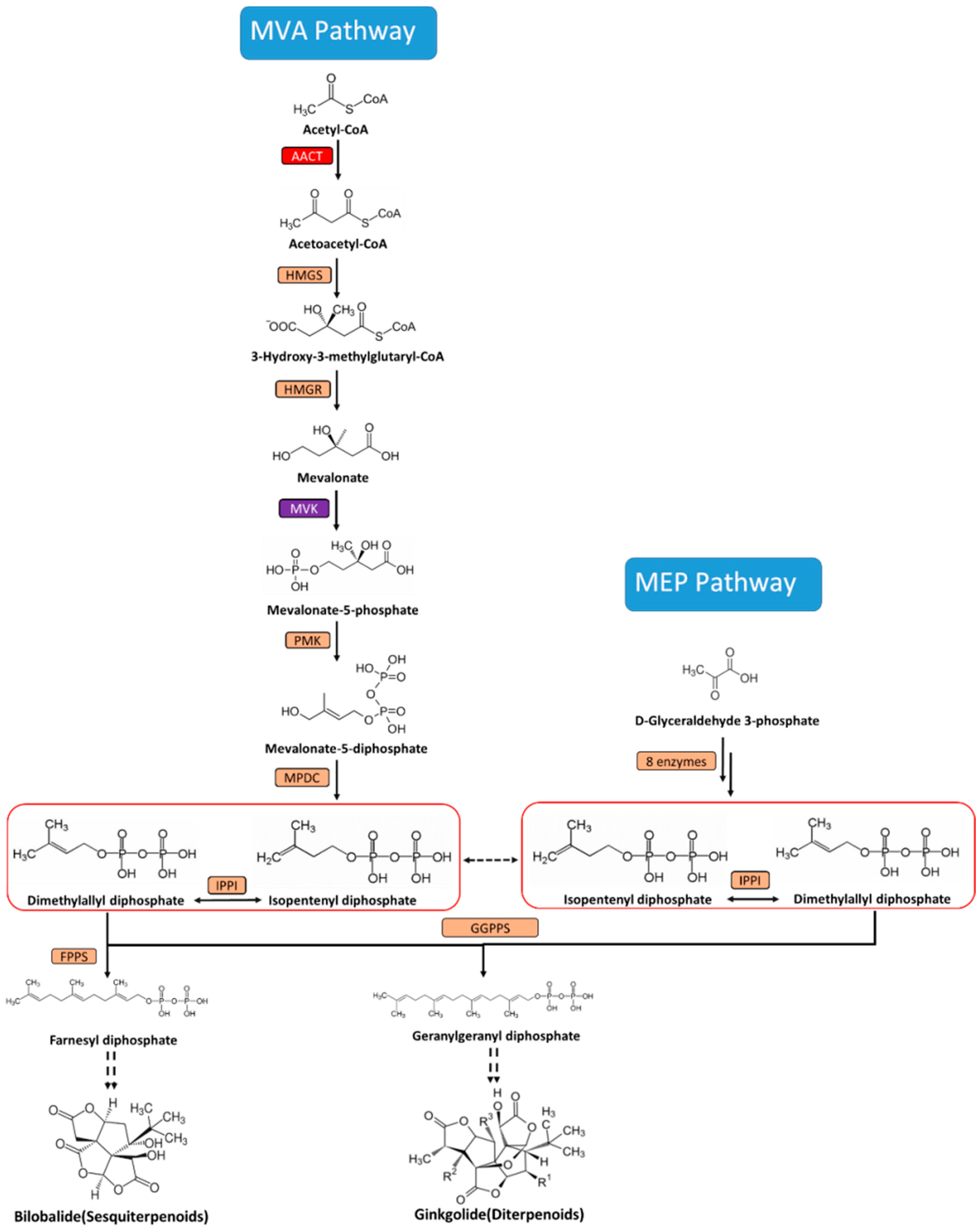
2. Results and Discussion
2.1. Isolation and Characterization of the cDNA of GbAACT and GbMVK
2.2. Bioinformatics Analysis of the Deduced GbAACT and GbMVK Protein
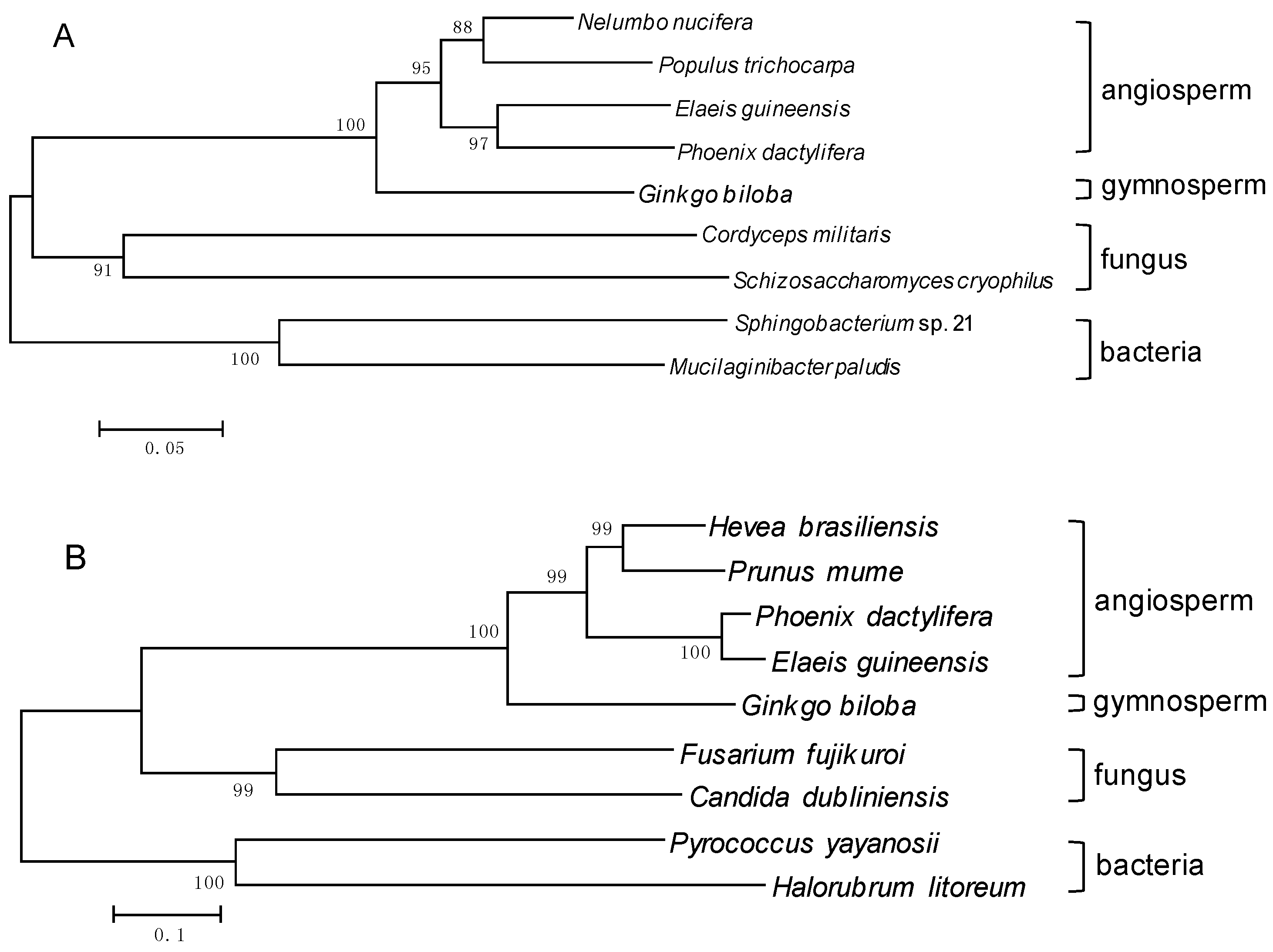
2.3. Molecular Evolution Analysis
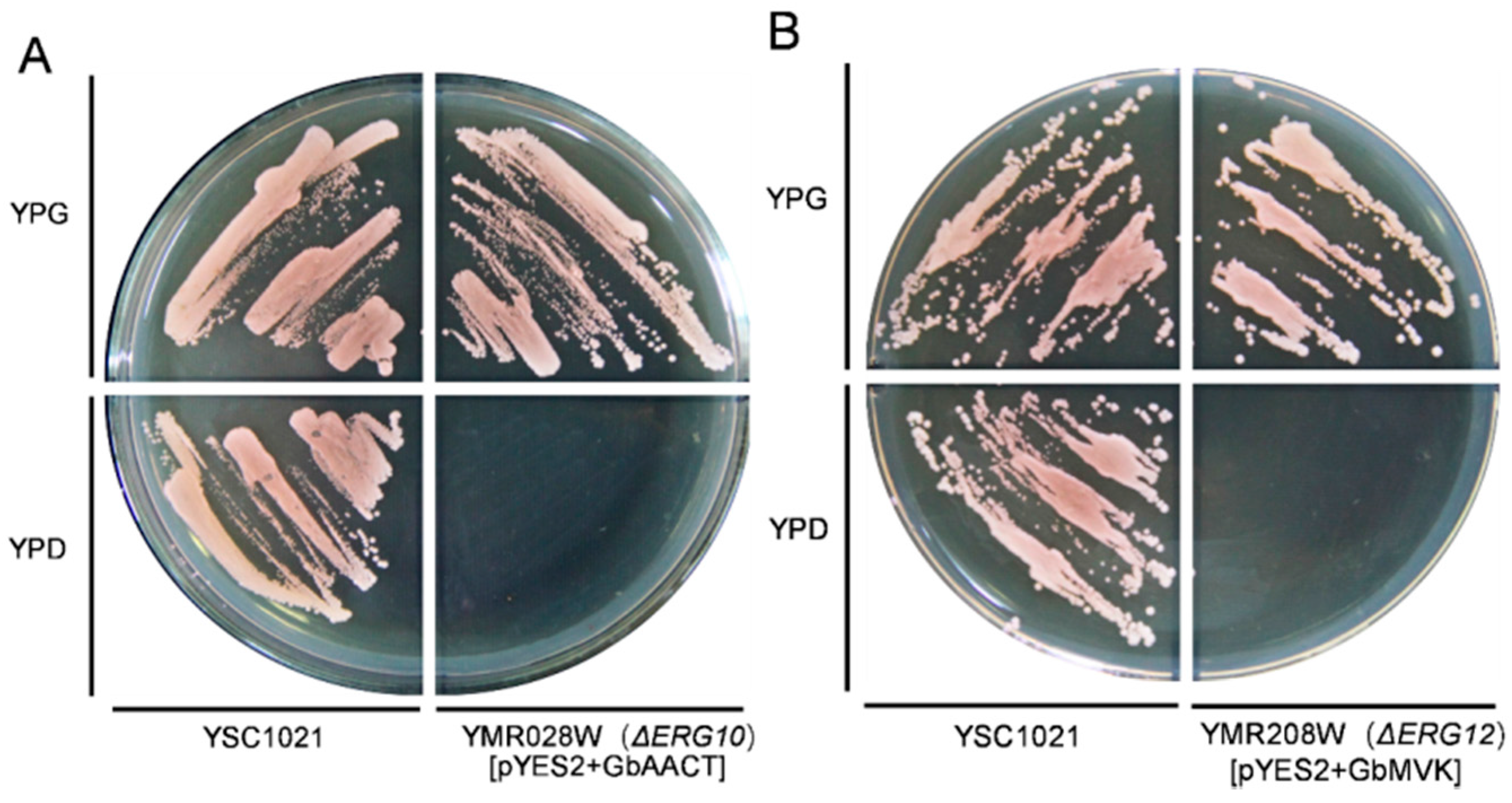
2.4. Functional Complementation of GbAACT and GbMVK in Saccharomyces Cerevisiae
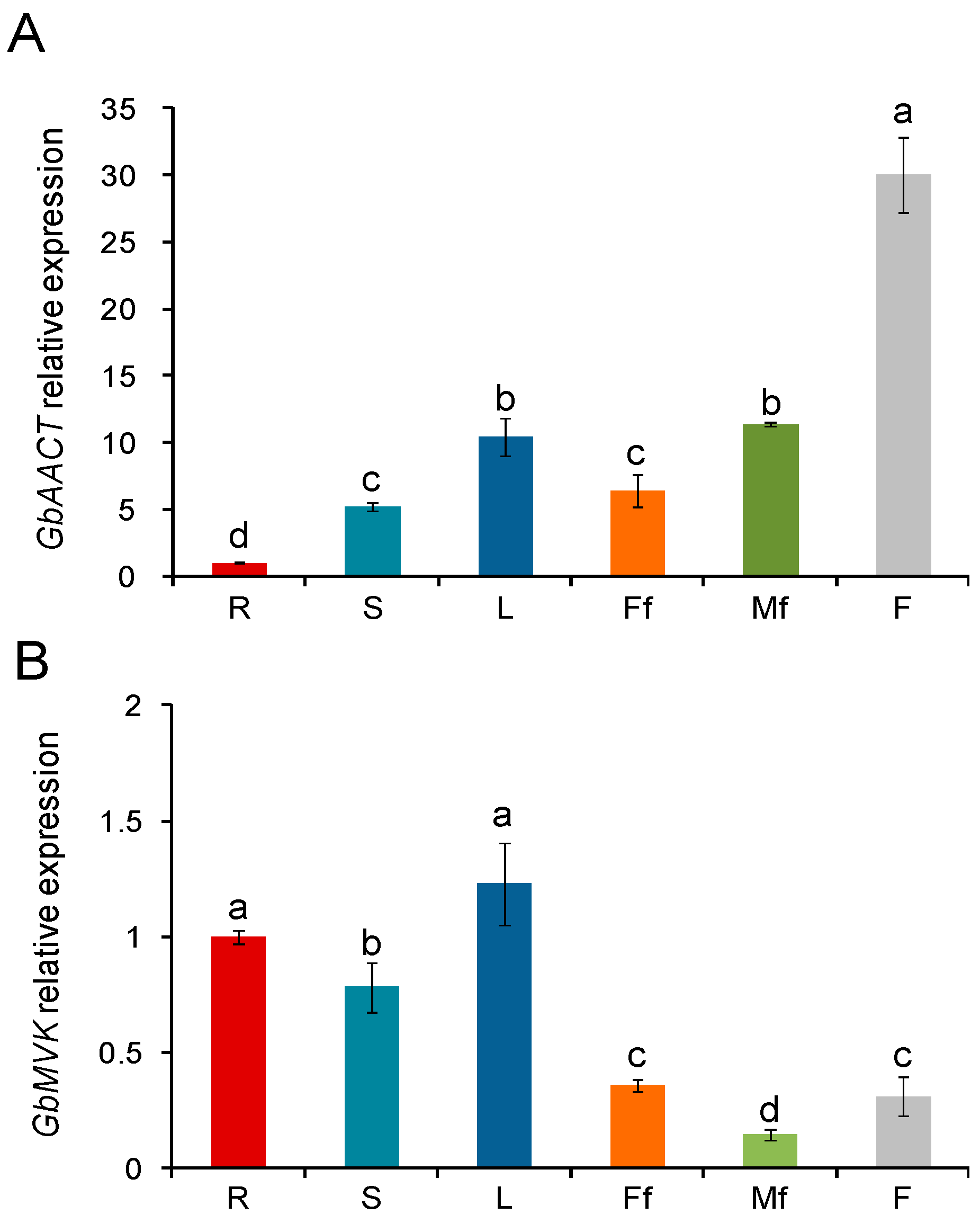
2.5. Transcript Level of the Gene Expression Pattern of GbAACT and GbMVK in Different Tissues of G. biloba
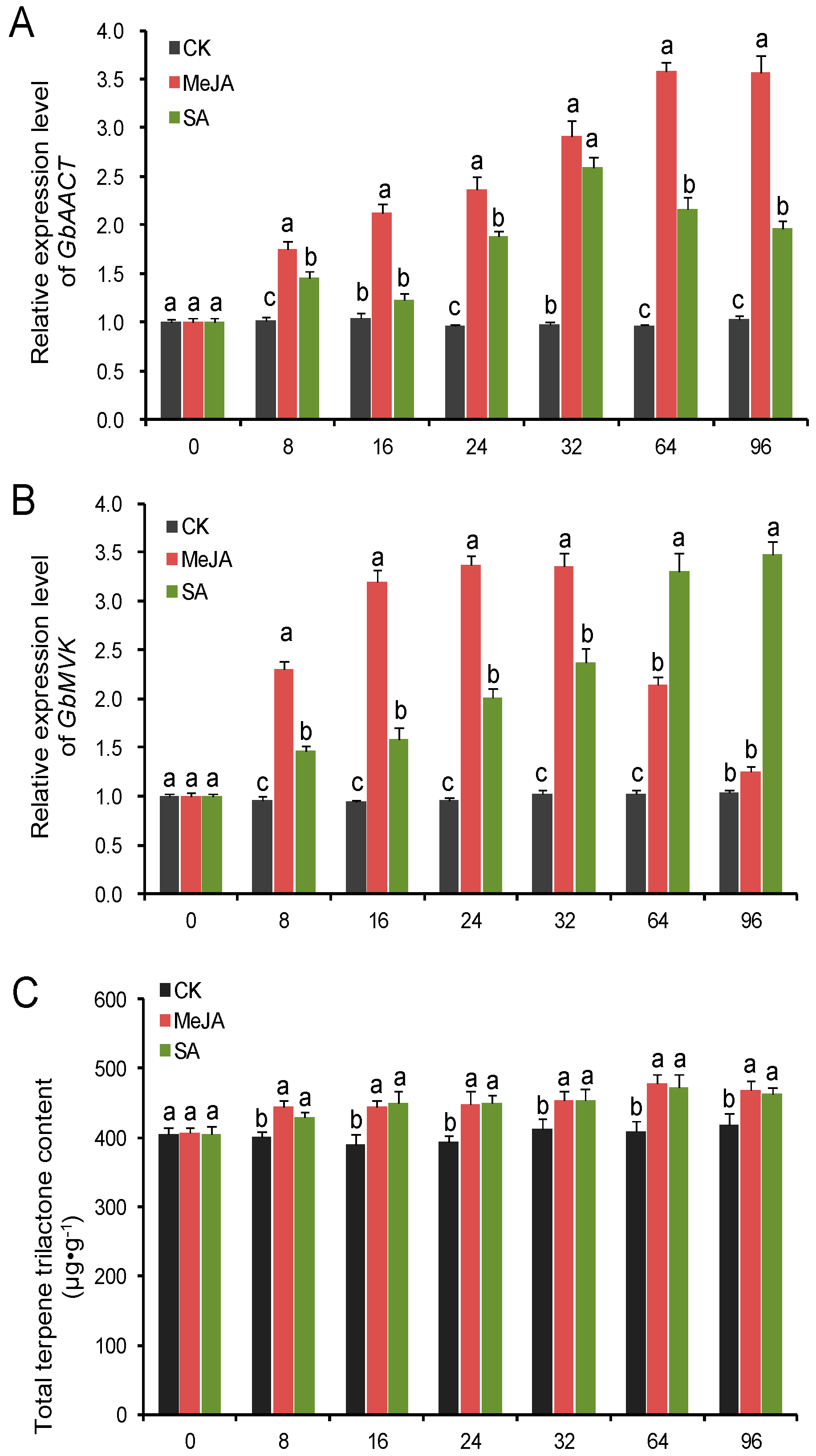
2.6. Transcript Level of GbAACT and GbMVK and TTL Content Changes in Ginkgo biloba under the Induction of MeJA and SA Elicitors
3. Experimental Section
3.1. Plant Materials
3.2. Cloning of the Full-Length cDNA of GbAACT and GbMVK
3.3. Bioinformatics Analysis and Molecular Evolution Analyses
3.4. Construction of Expression Plasmids for Yeast Complementation
3.5. GbAACT and GbMVK Tissue-Specific Analysis
3.6. Determination of TTL Contents in Ginkgo under Induction of MeJA and SA Elicitor
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carrier, D.J.; van Beek, T.A.; van der Heijden, R.; Verpoorte, R. Distribution of ginkgolides and terpenoid biosynthetic activity in Ginkgo biloba. Phytochemistry 1998, 48, 89–92. [Google Scholar] [CrossRef]
- Hosford, D.; Domingo, M.; Chabrier, P.; Braquet, P. Ginkgolides and platelet-activating factor binding sites. Methods Enzymol. 1990, 187, 433–446. [Google Scholar] [PubMed]
- Stromgaard, K.; Nakanishi, K. Chemistry and biology of terpene trilactones from Ginkgo biloba. Angew. Chem. 2004, 43, 1640–1658. [Google Scholar] [CrossRef] [PubMed]
- Braquent, P. BN 52021 and related compounds: A new series of highly specific paf-acether receptor antagonists. Prostaglandins 1985, 30, 687. [Google Scholar] [CrossRef]
- Jaracz, S.; Nakanishi, K.; Jensen, A.A.; Strømgaard, K. Ginkgolides and glycine receptors: A structure-activity relationship study. Chemistry 2004, 10, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Sabater-Jara, A.B.; Souliman-Youssef, S.; Novo-Uzal, E.; Almagro, L.; Belchí-Navarro, S.; Pedreño, M.A. Biotechnological approaches to enhance the biosynthesis of ginkgolides and bilobalide in Ginkgo biloba. Phytochem. Rev. 2013, 12, 191–205. [Google Scholar] [CrossRef]
- Lange, B.M.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Rohmer, M.; Schwender, J. Two independent biochemical pathways for isopentenyl diphosphate and isoprenoid biosynthesis in higher plants. Physiol. Plant. 1997, 101, 643–652. [Google Scholar] [CrossRef]
- Schwarz, M.A.D. Ginkgolide biosynthesis. In Comprehensive Natural Products Chemistry; Cane, D., Ed.; Pergamon Press: Oxford, UK, 1999; pp. 367–400. [Google Scholar]
- Pang, Y.; Shen, G.A.; Bergès, T.; Cardier, H.; Wu, W.; Sun, X.; Tang, K. Molecular cloning, characterization and heterologous expression in Saccharomyces cerevisiae of a mevalonate diphosphate decarboxylase cDNA from Ginkgo biloba. Physiol. Plant. 2006, 127, 19–27. [Google Scholar] [CrossRef]
- Liao, Y.; Xu, F.; Huang, X.; Zhang, W.; Cheng, H.; Wang, X.; Cheng, S.; Shen, Y. Characterization and transcriptional profiling of Ginkgo biloba mevalonate diphosphate decarboxylase gene (GbMVD) promoter towards light and exogenous hormone treatments. Plant Mol. Biol. Rep. 2016, 34, 566–581. [Google Scholar] [CrossRef]
- Shen, G.; Pang, Y.; Wu, W.; Liao, Z.; Zhao, L.; Sun, X.; Tang, K. Cloning and characterization of a root-specific expressing gene encoding 3-hydroxy-3-methylglutaryl coenzyme A reductase from Ginkgo biloba. Mol. Biol. Rep. 2006, 33, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.F.; Liao, Z.H.; Guo, B.H.; Sun, X.F.; Tang, K.X. Molecular cloning and expression profile analysis of Ginkgo biloba DXS gene encoding 1-deoxy-d-xylulose 5-phosphate synthase, the first committed enzyme of the 2-C-methyl-d-erythritol 4-phosphate pathway. Planta Med. 2006, 72, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kuzuyama, T.; Chang, Y.J.; Song, K.S.; Kim, S.U. Identification of class 2 1-deoxy-d-xylulose 5-phosphate synthase and 1-deoxy-d-xylulose 5-phosphate reductoisomerase genes from Ginkgo biloba and their transcription in embryo culture with respect to ginkgolide biosynthesis. Planta Med. 2006, 72, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, Y.B.; Kuzuyama, T.; Kim, S.U. Two copies of 4-(cytidine 5′-diphospho)-2-C-methyl-d-erythritol kinase (CMK) gene in Ginkgo biloba: Molecular cloning and functional characterization. Planta 2008, 228, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Lin, J.; Liu, X.; Deng, Z.; Li, Y.; Sun, X.; Tang, K. Molecular cloning, characterization and functional analysis of a 2C-methyl-d-erythritol 2,4-cyclodiphosphate synthase gene from Ginkgo biloba. J. Biochem. Mol. Biol. 2006, 39, 502–510. [Google Scholar] [PubMed]
- Kim, S.M.; Kuzuyama, T.; Chang, Y.J.; Kim, S.U. Cloning and characterization of 2-C-methyl-d-erythritol 2,4-cyclodiphosphate synthase (MECS) gene from Ginkgo biloba. Plant Cell Rep. 2006, 25, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, S.U. Characterization of 1-hydroxy-2-methyl-2-(E)-butenyl-4-diphosphate synthase (HDS) gene from Ginkgo biloba. Mol. Biol. Rep. 2010, 37, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kuzuyama, T.; Kobayashi, A.; Sando, T.; Chang, Y.J.; Kim, S.U. 1-Hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate reductase (IDS) is encoded by multicopy genes in gymnosperms Ginkgo biloba and Pinus taeda. Planta 2008, 227, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, K.I.; Chang, Y.J.; Kim, S.U. Developmental pattern of Ginkgo biloba levopimaradiene synthase (GbLPS) as probed by promoter analysis in Arabidopsis thaliana. Plant Cell Rep. 2012, 31, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Khadijah Hanim, A.R.; Samian, M.R. Cloning and promoter identification of acetoacetyl-CoA thiolase gene from oil palm, Elaeis guineensis Jacq. Int. J. Environ. Sci. Dev. 2014, 5, 362–366. [Google Scholar]
- Fang, X.; Shi, L.; Ren, A.; Jiang, A.L.; Wu, F.L.; Zhao, M.W. The cloning, characterization and functional analysis of a gene encoding an acetyl-CoA acetyltransferase involved in triterpene biosynthesis in Ganoderma lucidum. Mycoscience 2013, 54, 100–105. [Google Scholar] [CrossRef]
- Vishwakarma, R.K.; Singh, S.; Sonawane, P.D.; Srivastava, S.; Kumari, U.; Kumar, R.J.S.; Khan, B.M. Molecular cloning, biochemical characterization, and differential expression of an acetyl-CoA C-acetyltransferase gene (AACT) of Brahmi (Bacopa monniera). Plant Mol. Biol. Rep. 2013, 31, 547–557. [Google Scholar] [CrossRef]
- Wuyuntana; Wang, L.; Ye, S.J. Identification and bioinformatics analysis of EuMK gene in Eucommia ulmoides. Nonwood For. Res. 2014, 32, 6–12. [Google Scholar]
- Deng, X.M.; Shao-Hua, W.U.; Dai, X.M.; Tian, W.M. Expression analysis of MVA and MEP metabolic pathways genes in latex and suspension cells of Hevea brasiliensis. Guihaia 2016, 36, 449–455. [Google Scholar]
- Liao, Y.L.; Xu, F.; Huang, X.H.; Zhang, W.W.; Cheng, H.; Li, L.L.; Cheng, S.Y.; Shen, Y.B. Promoter analysis and transcriptional profiling of Ginkgo biloba 3-hydroxy-3-methylglutaryl coenzyme a reductase (GbHMGR) gene in abiotic stress responses. Not. Bot. Horti Agrobot. 2015, 43, 25–34. [Google Scholar] [CrossRef]
- Dyer, J.H.; Maina, A.; Gomez, I.D.; Cadet, M.; Oeljeklaus, S.; Schiedel, A.C. Cloning, expression and purification of an acetoacetyl CoA thiolase from sunflower cotyledon. Int. J. Biol. Sci. 2009, 5, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Anderson, V.E.; Bahnson, B.J.; Wlassics, I.D.; Walsh, C.T. The reaction of acetyldithio-CoA, a readily enolized analog of acetyl-CoA with thiolase from Zoogloea ramigera. J. Biol. Chem. 1990, 265, 6255–6261. [Google Scholar] [PubMed]
- Williams, S.F.; Palmer, M.A.; Peoples, O.P.; Walsh, C.T.; Sinskey, A.J.; Masamune, S. Biosynthetic thiolase from Zoogloea ramigera. Mutagenesis of the putative active-site base Cys-378 to Ser-378 changes the partitioning of the acetyl S-enzyme intermediate. J. Biol. Chem. 1992, 267, 16041–16043. [Google Scholar] [PubMed]
- Yang, S.Y.; Yang, X.Y.; Healy-Louie, G.; Schulz, H.; Elzinga, M. Nucleotide sequence of the fadA gene. Primary structure of 3-ketoacyl-coenzyme A thiolase from Escherichia coli and the structural organization of the fadAB operon. J. Biol. Chem. 1990, 266, 10424–10429. [Google Scholar]
- Sando, T.; Takaoka, C.; Mukai, Y.; Yamashita, A.; Hattori, M.; Ogasawara, N.; Fukusaki, E.; Kobayashi, A. Cloning and characterization of mevalonate pathway genes in a natural rubber producing plant, Hevea brasiliensis. Biosci. Biotechnol. Biochem. 2008, 72, 2049–2060. [Google Scholar] [CrossRef] [PubMed]
- Servouse, M.; Karst, F. Regulation of early enzymes of ergosterol biosynthesis in Saccharomyces cerevisiae. Biochem. J. 1986, 240, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Trocha, P.J.; Sprinson, D.B. Location and regulation of early enzymes of sterol biosynthesis in yeast. Arch. Biochem. Biophys. 1976, 174, 45–51. [Google Scholar] [CrossRef]
- Oulmouden, A.; Karst, F. Isolation of the ERG12 gene of Saccharomyces cerevisiae encoding mevalonate kinase. Gene 1990, 88, 253–257. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Su, X.H.; Dong, C.M.; Chen, S.Q.; Shao, Y.Y.; Zhang, F.B. Cloning and expression analysis of acetyl-CoA C-acetyltransferase gene in Isodon rubeseens. J. Chin. Med. Mater. 2016, 39, 37–41. [Google Scholar]
- Guo, X.; Luo, H.M.; Chen, S.L. Cloning and analysis of mevalonate kinase (PnMVK1) gene in Panax notoginseng. Acta Pharm. Sin. 2012, 47, 1092–1097. [Google Scholar]
- Cartayrade, A.; Neau, E.; Sohier, C.; Balz, J.P.; Carde, J.P.; Walter, J. Ginkgolide and bilobalide biosynthesis in Ginkgo biloba. 1. Sites of synthesis, translocation and accumulation of ginkgolides and bilobalide. Plant Physiol. Biochem. 1997, 35, 859–868. [Google Scholar]
- Erb, M.; Lenk, C.; Degenhardt, J.; Turlings, T.C. The underestimated role of roots in defense against leaf attackers. Trends Plant Sci. 2009, 14, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Robertseilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, A.; Leonreyes, A.; Ritsema, T.; Verhage, A.; Den Otter, F.C.; Van Loon, L.C.; Pieterse, C.M. Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation. Plant Physiol. 2008, 147, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Brader, G.; Palva, E.T. The WRKY70 transcription factor: A node of convergence for jasmonate-mediated and salicylate-mediated signals in plant defense. Plant Cell 2004, 16, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Küpper, F.C.; Gaquerel, E.; Cosse, A.; Adas, F.; Peters, A.F.; Müller, D.G.; Kloareg, B.; Salaün, J.P.; Potin, P. Free fatty acids and methyl jasmonate trigger defense reactions in Laminaria digitata. Plant Cell Physiol. 2009, 50, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Beckers, G.J.; Spoel, S.H. Fine-tuning plant defence signalling: Salicylate versus jasmonate. Plant Biol. 2006, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.M.; Min, J.Y.; Kim, Y.D.; Kang, Y.M.; Park, D.J.; Jung, H.N.; Kim, S.W.; Choi, M.S. Effects of methyl jasmonate and salicylic acid on the production of bilobalide and ginkgolides in cell cultures of Ginkgo biloba. In Vitro Cell. Dev. Biol. Plant 2006, 42, 44–49. [Google Scholar] [CrossRef]
- Kang, M.K.; Nargis, S.; Kim, S.M.; Kim, S.U. Distinct expression patterns of two Ginkgo biloba 1-hydroxy-2-methyl-2-(E)-butenyl-4-diphosphate reductase/isopentenyl diphospahte synthase (HDR/IDS) promoters in Arabidopsis model. Plant Physiol. Biochem. 2013, 62, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.L.; Shen, Y.B.; Chang, J.; Zhang, W.W.; Cheng, S.Y.; Xu, F. Isolation, expression, and promoter analysis of GbWRKY2: A novel transcription factor gene from Ginkgo biloba. Int. J. Genom. 2015, 2015, 607185. [Google Scholar]
- Lu, G.; Moriyama, E.N. Vector NTI, a balanced all-in-one sequence analysis suite. Brief Bioinform. 2004, 5, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Jansson, S.; Meyer-Gauen, G.; Cerff, R.; Martin, W. Nucleotide distribution in gymnosperm nuclear sequences suggests a model for GC-content change in land-plant nuclear genomes. J. Mol. Evol. 1994, 39, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.L.; Xu, F.; Zhu, J.; Wang, Y.; Cheng, S.Y. Separation and dtermination of terpene trilactones by gas chromatography with wide bore capillary column. Acta Agric. Boreali-Occident. Sin. 2008, 17, 146–149. [Google Scholar]
- Sample Availability: Samples of Ginkgo biloba and cDNAs of GbAACT and GbMVK are available from the authors.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Usage | Primer Name | Primer Sequence 5′-3′ |
|---|---|---|
| ORF-PCR | GbAACT-F | ATGGGTTCTACATCAGGCTCAGATTTGTT |
| GbAACT-R | TTACATGAGCTCCAAGACAACTGCAG | |
| GbMVK-F | ATATTGGGAGTTCAGAATGGTGC | |
| GbMVK-R | ACATTTATGAGCTTGAGCATTGGAA | |
| qRT-PCR | GbAACT-qF | CATTGGGGGTCTGAGTGGTTC |
| GbAACT-qR | CATTGTTGCTTTCATTCCCGA | |
| GbMVK-qF | GGCATTAGTTGCTGGAGTTTCTGAG | |
| GbMVK-qR | GGTGCCTGAATAATCTCTGCCATCT | |
| GbGAPDHF | CTGGCGTAGAGTATGTGGTTGAAT | |
| GbGAPDHR | CACGCCAACAACGAACATG | |
| Yeast-PCR | GbAACT-yup | CGGGATCCATGGGTTCTACATCAGGCTCAG |
| GbAACT-yud | CGGAATTCTTACATGAGCTCCAAGACAACTG | |
| GbMVK-yup | CGGGATCCATATTGGGAGTTCAGAATGGTGC | |
| GbMVK-yud | CGGAATTCACATTTATGAGCTTGAGCATTGGAA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Yan, J.; Meng, X.; Xu, F.; Zhang, W.; Liao, Y.; Qu, J. Molecular Cloning, Characterization, and Functional Analysis of Acetyl-CoA C-Acetyltransferase and Mevalonate Kinase Genes Involved in Terpene Trilactone Biosynthesis from Ginkgo biloba. Molecules 2017, 22, 74. https://doi.org/10.3390/molecules22010074
Chen Q, Yan J, Meng X, Xu F, Zhang W, Liao Y, Qu J. Molecular Cloning, Characterization, and Functional Analysis of Acetyl-CoA C-Acetyltransferase and Mevalonate Kinase Genes Involved in Terpene Trilactone Biosynthesis from Ginkgo biloba. Molecules. 2017; 22(1):74. https://doi.org/10.3390/molecules22010074
Chicago/Turabian StyleChen, Qiangwen, Jiaping Yan, Xiangxiang Meng, Feng Xu, Weiwei Zhang, Yongling Liao, and Jinwang Qu. 2017. "Molecular Cloning, Characterization, and Functional Analysis of Acetyl-CoA C-Acetyltransferase and Mevalonate Kinase Genes Involved in Terpene Trilactone Biosynthesis from Ginkgo biloba" Molecules 22, no. 1: 74. https://doi.org/10.3390/molecules22010074
APA StyleChen, Q., Yan, J., Meng, X., Xu, F., Zhang, W., Liao, Y., & Qu, J. (2017). Molecular Cloning, Characterization, and Functional Analysis of Acetyl-CoA C-Acetyltransferase and Mevalonate Kinase Genes Involved in Terpene Trilactone Biosynthesis from Ginkgo biloba. Molecules, 22(1), 74. https://doi.org/10.3390/molecules22010074