Astragalus Polysaccharide Protect against Cadmium-Induced Cytotoxicity through the MDA5/NF-κB Pathway in Chicken Peripheral Blood Lymphocytes

 ,
,
Abstract
:1. Introduction
2. Results
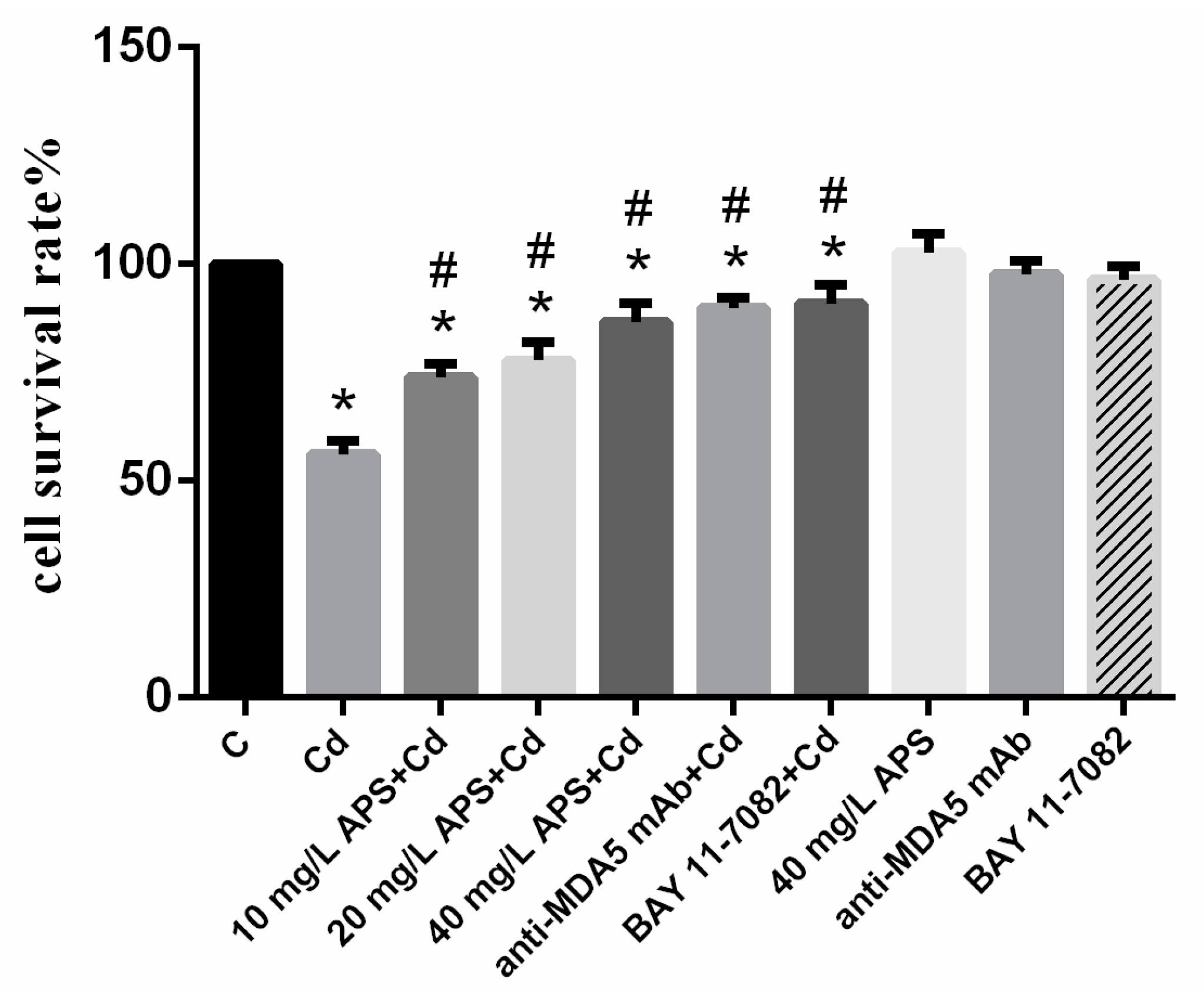
2.1. Cell Viability
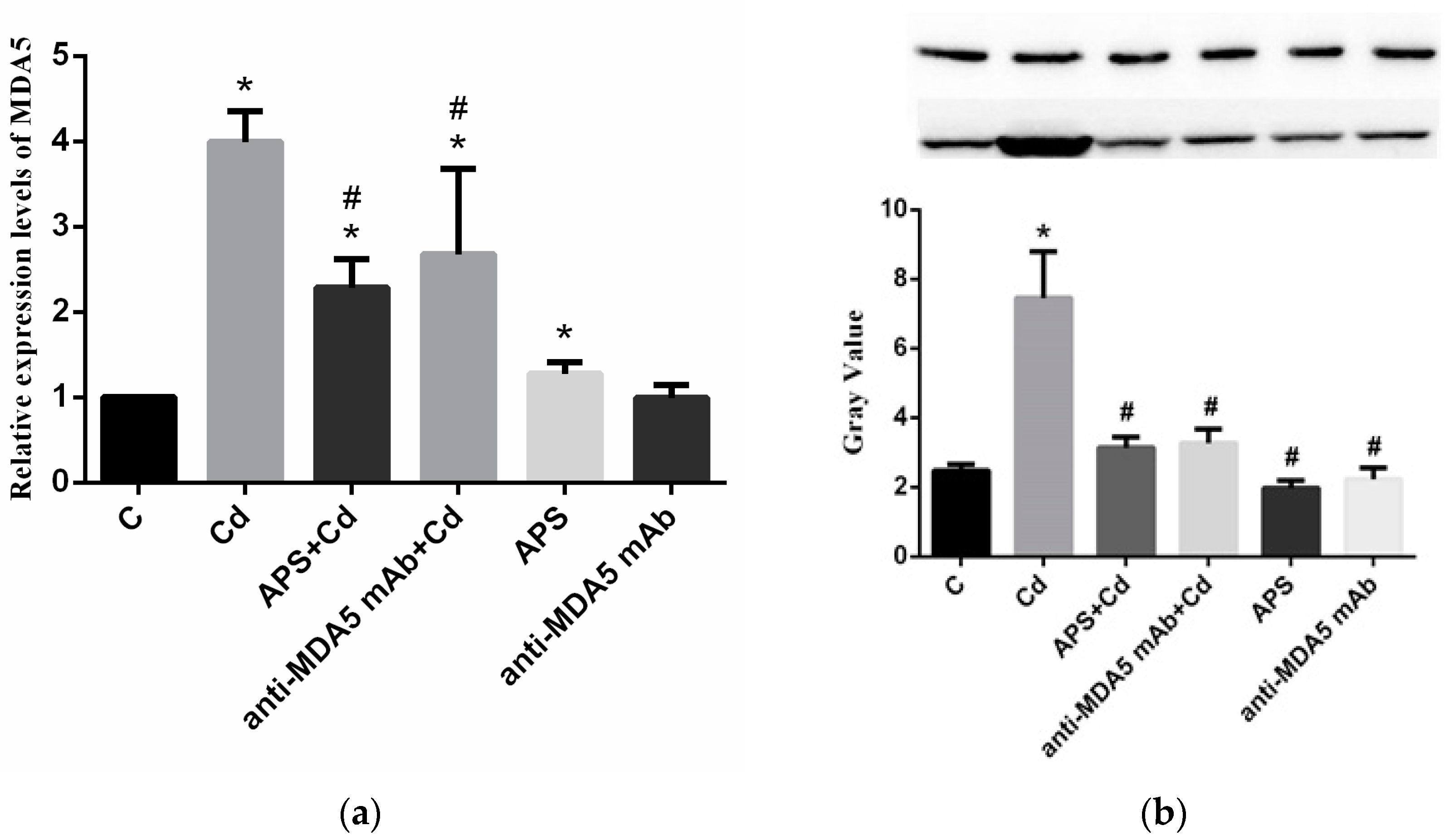
2.2. Expression of MDA5
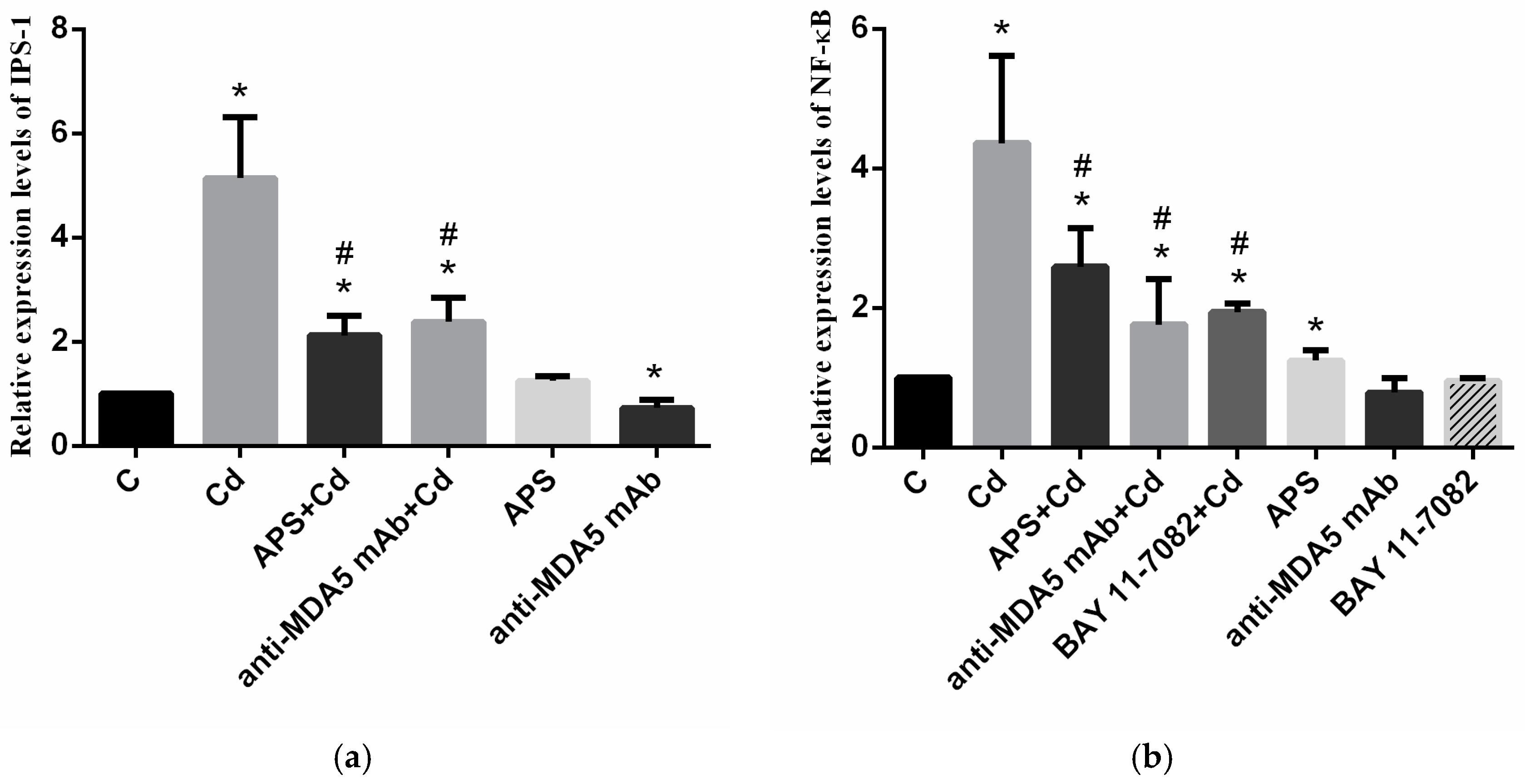
2.3. mRNA Levels of MDA5’s Downstream Signaling Molecule
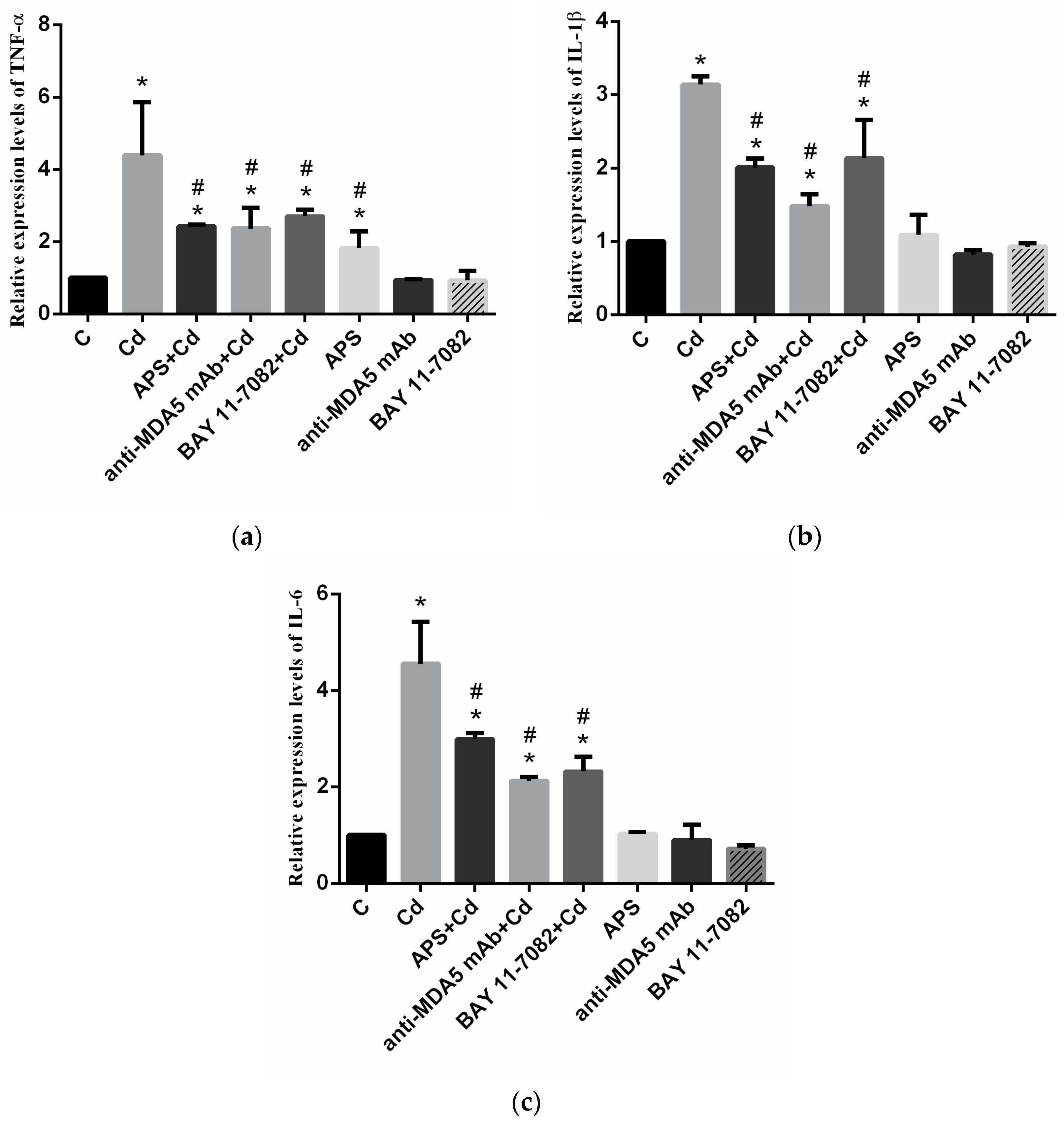
2.4. mRNA Levels of Cytokines
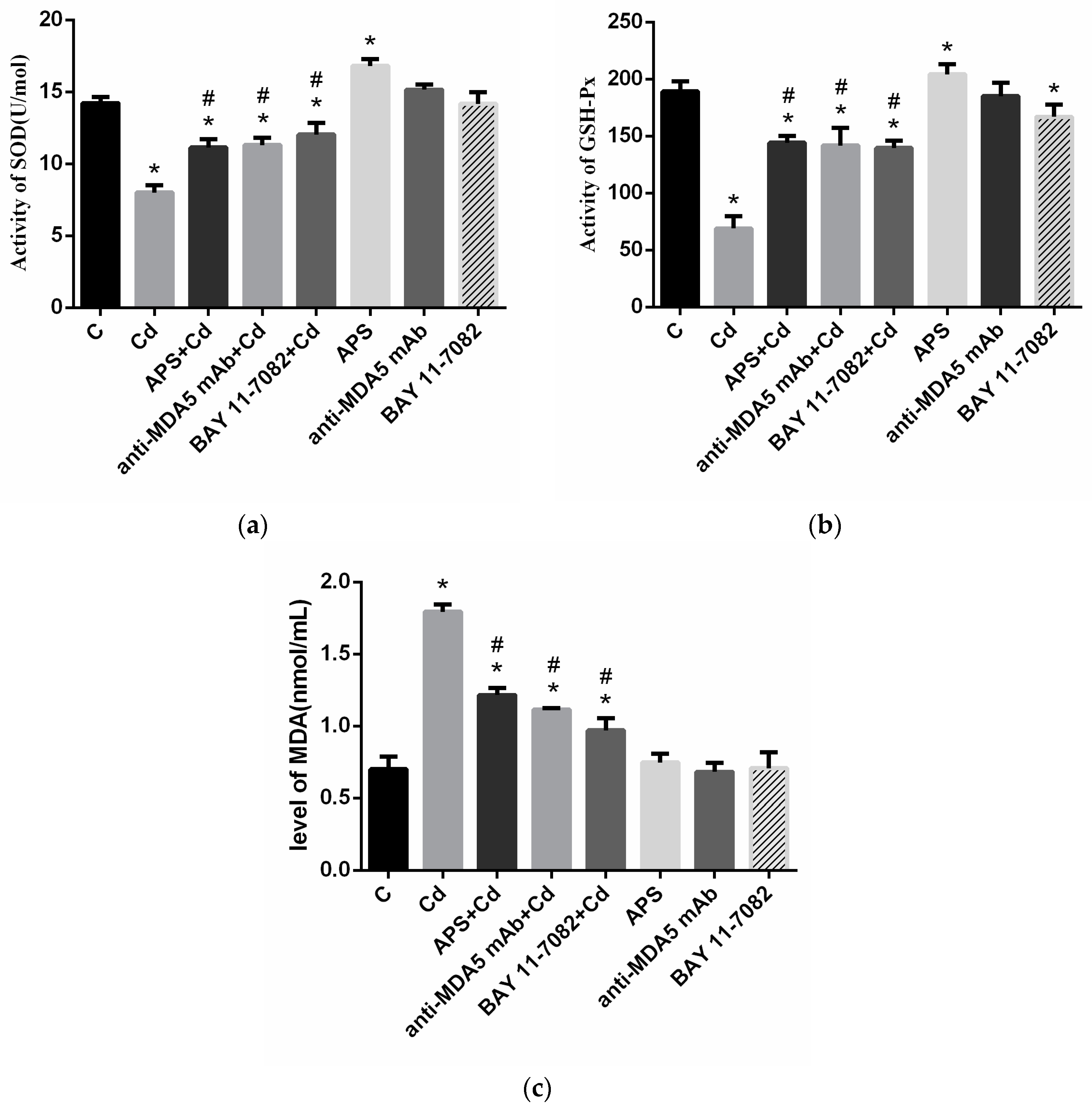
2.5. Antioxidant Activity
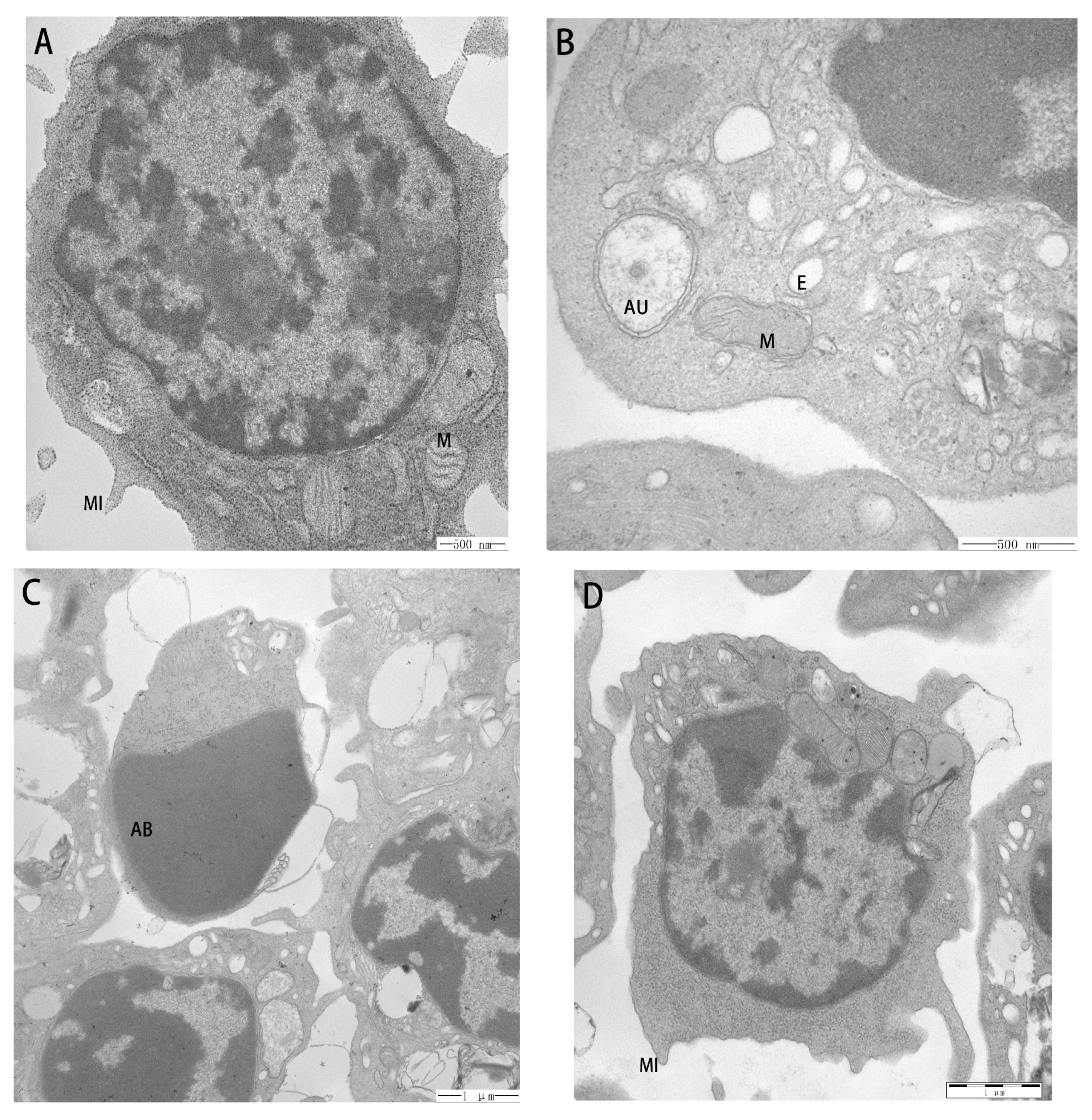
2.6. Ultrastructural Pathological Changes
3. Discussion
4. Materials and Methods
4.1. Preparation of Chicken PBLs and Treatment
4.2. MTT Assay
4.3. Quantitative Real-Time PCR
4.4. Western Blotting Analysis
4.5. Measurement of Oxidative Stress
4.6. Morphological Study
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mera, R.; Torres, E.; Abalde, J. Influence of sulphate on the reduction of cadmium toxicity in the microalga Chlamydomonas moewusii. Ecotoxicol. Environ. Saf. 2016, 128, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Van Maele-Fabry, G.; Lombaert, N.; Lison, D. Dietary exposure to cadmium and risk of breast cancer in postmenopausal women: A systematic review and meta-analysis. Environ. Int. 2016, 86, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lag, M.; Refsnes, M.; Lilleaas, E.M.; Holme, J.A.; Becher, R.; Schwarze, P.E. Role of mitogen activated protein kinases and protein kinase C in cadmium-induced apoptosis of primary epithelial lung cells. Toxicology 2005, 211, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Huang, Y.; Zhang, K.; Huang, Y.; Yan, Y.; Wang, F.; Wu, J.; Wang, X.; Xu, Z.; Chen, Y.; et al. Cadmium-induced immune abnormality is a key pathogenic event in human and rat models of preeclampsia. Environ. Pollut. 2016, 218, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhang, Z.; He, J.; Li, J.; Zhang, J.; Xing, H.; Xu, S. Ovarian toxicity induced by dietary cadmium in hen. Biol. Trace Elem. Res. 2012, 148, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Z.; Han, C.; Zhang, Z.; Xu, S. Effects of dietary manganese on Cu, Fe, Zn, Ca, Se, IL-1beta, and IL-2 changes of immune organs in cocks. Biol. Trace Elem. Res. 2012, 148, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Pathak, N.; Khandelwal, S. Influence of cadmium on murine thymocytes: Potentiation of apoptosis and oxidative stress. Toxicol. Lett. 2006, 165, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Pathak, N.; Khandelwal, S. Oxidative stress and apoptotic changes in murine splenocytes exposed to cadmium. Toxicology 2006, 220, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Fagerberg, B.; Borne, Y.; Barregard, L.; Sallsten, G.; Forsgard, N.; Hedblad, B.; Persson, M.; Engstrom, G. Cadmium exposure is associated with soluble urokinase plasminogen activator receptor, a circulating marker of inflammation and future cardiovascular disease. Environ. Res. 2017, 152, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Meylan, E.; Tschopp, J. Intracellular pattern-recognition receptors. Adv. Drug Deliv. Rev. 2008, 60, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Belgnaoui, S.M.; Paz, S.; Hiscott, J. Orchestrating the interferon antiviral response through the mitochondrial antiviral signaling (MAVS) adapter. Curr. Opin. Immunol. 2011, 23, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Lv, A.; Ge, M.; Hu, X.; Liu, W.; Li, G.; Zhang, R. Effects of Agaricus blazei murill polysaccharide on cadmium poisoning on the MDA5 signaling pathway and antioxidant function of chicken peripheral blood lymphocytes. Biol. Trace Elem. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Li, X.T.; Zhang, Y.K.; Kuang, H.X.; Jin, F.X.; Liu, D.W.; Gao, M.B.; Liu, Z.; Xin, X.J. Mitochondrial protection and anti-aging activity of astragalus polysaccharides and their potential mechanism. Int. J. Mol. Sci. 2012, 13, 1747–1761. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Chen, X.; Zhang, Y.; Xu, J.; Zhang, L.; Li, Z.; Liu, W.; Ouyang, J.; Han, S.; He, X. Astragalus polysaccharide induces anti-inflammatory effects dependent on ampk activity in palmitate-treated Raw264.7 cells. Int. J. Mol. Med. 2013, 31, 1463–1470. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Qin, J.; Liu, M.; Luo, J.; Ding, F.; Wang, M.; Zheng, L. Astragalus polysaccharide attenuates lipopolysaccharide-induced inflammatory responses in microglial cells: Regulation of protein kinase B and nuclear factor-κB signaling. Inflamm. Res. 2015, 64, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Zhao, L.H.; Zhi, C.; Pan, J.P. Regulation on maturation and function of dendritic cells by astragalus mongholicus polysaccharides. Int. Immunopharmacol. 2006, 6, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Shu, J.; Xu, L.; Lu, C.; Lu, A. Inhibitory effect of astragalus polysaccharides on lipopolysaccharide-induced TNF-a and IL-1beta production in THP-1 cells. Molecules 2012, 17, 3155–3164. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, F.; Yang, Y.; Li, D.; Lv, J.; Ou, Y.; Sun, F.; Chen, J.; Shi, Y.; Xia, P. Astragalus polysaccharide ameliorates ionizing radiation-induced oxidative stress in mice. Int. J. Biol. Macromol. 2014, 68, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, Z.; Wang, Z.; Yu, S.; Long, T.; Zhou, X.; Bao, Y. Astragalus polysaccharides exerts immunomodulatory effects via tlr4-mediated myd88-dependent signaling pathway In Vitro and In Vivo. Sci. Rep. 2017, 7, 44822. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, Y.H.; Cheng, X.Y.; Zhang, Z.W.; Xu, S.W. The protection of selenium against cadmium-induced cytotoxicity via the heat shock protein pathway in chicken splenic lymphocytes. Molecules 2012, 17, 14565–14572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yu, Q.; Shi, G.; Liu, R.; Zhang, W.; Zhao, X.; Li, G.; Ge, M. Chtlr4 pathway activation by astragalus polysaccharide in bursa of fabricius. BMC Vet. Res. 2017, 13, 119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Liu, W.; Chen, X.; Zhu, Y.; Zhang, Z.; Yao, H.; Xu, S. Four endoplasmic reticulum resident selenoproteins may be related to the protection of selenium against cadmium toxicity in chicken lymphocytes. Biol. Trace Elem. Res. 2014, 161, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Claudio, S.R.; Gollucke, A.P.; Yamamura, H.; Morais, D.R.; Bataglion, G.A.; Eberlin, M.N.; Peres, R.C.; Oshima, C.T.; Ribeiro, D.A. Purple carrot extract protects against cadmium intoxication in multiple organs of rats: Genotoxicity, oxidative stress and tissue morphology analyses. J. Trace Elem. Med. Biol. 2016, 33, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Saidi, I.; Chtourou, Y.; Djebali, W. Selenium alleviates cadmium toxicity by preventing oxidative stress in sunflower (Helianthus annuus) seedlings. J. Plant Physiol. 2014, 171, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; He, J.; Wang, B.; Zhang, F.; Chen, G.; Yin, S.; Shen, H. Suppressive effect of Astragalus membranaceus bunge on chemical hepatocarcinogenesis in rats. Cancer Chemother. Pharmacol. 2003, 51, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.M.; Xu, W.; Dai, H.; Tu, P.; Li, Z.; Gao, X.M. A study on the immune receptors for polysaccharides from the roots of Astragalus membranaceus, a chinese medicinal herb. Biochem. Biophys. Res. Commun. 2004, 320, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.F.; Lu, L.; Zhu, D.J.; Wang, M.; Yin, Y.; Chen, D.X.; Wei, L.B. Effects of astragalus polysaccharides on dysfunction of mitochondrial dynamics induced by oxidative stress. Oxid. Med. Cell. Longev. 2016, 2016, 9573291. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.O.; Wang, X.; Hitron, J.A.; Zhang, Z.; Cheng, S.; Budhraja, A.; Ding, S.; Lee, J.C.; Shi, X. Cadmium induces autophagy through ros-dependent activation of the LKB1-AMPK signaling in skin epidermal cells. Toxicol. Appl. Pharmacol. 2011, 255, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Lu, L.; Wang, D.; Shi, Y.; Wang, M.; Huang, Y.; Chen, D.; Deng, C.; Chen, J.; Lv, P.; et al. Astragalus polysaccharide inhibits autophagy and apoptosis from peroxide-induced injury in c2c12 myoblasts. Cell Biochem. Biophys. 2015, 73, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Gumuslu, S. Cold-stress-induced modulation of antioxidant defence: Role of stressed conditions in tissue injury followed by protein oxidation and lipid peroxidation. Int. J. Biometeorol. 2004, 48, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Jihen el, H.; Imed, M.; Fatima, H.; Abdelhamid, K. Protective effects of selenium (Se) and zinc (Zn) on cadmium (Cd) toxicity in the liver of the rat: Effects on the oxidative stress. Ecotoxicol. Environ. Saf. 2009, 72, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhang, R.; Xie, Y.; Wang, H.; Ge, M. The protective effects of polysaccharides from Agaricus blazei murill against cadmium-induced oxidant stress and inflammatory damage in chicken livers. Biol. Trace Elem. Res. 2017, 178, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, V.K.; Bhatanagar, A.; Chaudhary, M. Protective role of ceftriaxone plus sulbactam with VRP1034 on oxidative stress, hematological and enzymatic parameters in cadmium toxicity induced rat model. Interdiscip. Toxicol. 2012, 5, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fang, J.; Leonard, S.S.; Rao, K.M. Cadmium inhibits the electron transfer chain and induces reactive oxygen species. Free Radic. Biol. Med. 2004, 36, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Jiang, C.Y.; Li, S.; Xu, S.W. Cadmium induced hepatotoxicity in chickens (Gallus domesticus) and ameliorative effect by selenium. Ecotoxicol. Environ. Saf. 2013, 96, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xu, F.P.; Yang, Z.J.; Li, M.; Min, Y.H.; Li, S. Cadmium-induced injury and the ameliorative effects of selenium on chicken splenic lymphocytes: Mechanisms of oxidative stress and apoptosis. Biol. Trace Elem. Res. 2014, 160, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Yan, G.; Li, Y.; Han, Z.; Zhang, L.; Chen, S.; Feng, C.; Huang, Q.; Ding, F.; Yu, Y.; et al. Astragalus polysaccharide attenuated iron overload-induced dysfunction of mesenchymal stem cells via suppressing mitochondrial ros. Cell. Physiol. Biochem. 2016, 39, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Kudr, J.; Nguyen, H.V.; Gumulec, J.; Nejdl, L.; Blazkova, I.; Ruttkay-Nedecky, B.; Hynek, D.; Kynicky, J.; Adam, V.; Kizek, R. Simultaneous automatic electrochemical detection of zinc, cadmium, copper and lead ions in environmental samples using a thin-film mercury electrode and an artificial neural network. Sensors 2014, 15, 592–610. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Son, Y.O.; Pratheeshkumar, P.; Shi, X. Oxidative stress and metal carcinogenesis. Free Radic. Biol. Med. 2012, 53, 742–757. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Qin, J.; Hao, Y.; Liu, M.; Luo, J.; Luo, T.; Wei, L. Astragalus polysaccharide suppresses skeletal muscle myostatin expression in diabetes: Involvement of ros-erk and nf-kappab pathways. Oxid. Med. Cell. Longev. 2013, 2013, 782497. [Google Scholar] [CrossRef] [PubMed]
- Black, P.H. Stress and the inflammatory response: A review of neurogenic inflammation. Brain Behav. Immun. 2002, 16, 622–653. [Google Scholar] [CrossRef]
- Hyun, J.S.; Satsu, H.; Shimizu, M. Cadmium induces interleukin-8 production via nf-kappab activation in the human intestinal epithelial cell, Caco-2. Cytokine 2007, 37, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Liu, S.; Li, S. Effects of selenium and cadmium on changes in the gene expression of immune cytokines in chicken splenic lymphocytes. Biol. Trace Elem. Res. 2015, 165, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-Y.; Gao, Y.-H.; Wang, Z.-Y.; Xu, B.; Wu, A.-M.; Xing, Y.-W.; Liu, B.; Lou, L.-X.; Chen, L.-X. Astragalus polysaccharide suppresses the expression of adhesion molecules through the regulation of the p38 mapk signaling pathway in human cardiac microvascular endothelial cells after ischemia-reperfusion injury. Evid. Based Complement. Altern. Med. 2013, 2013, 280493. [Google Scholar]
- Li, J.L.; Li, S.; Tang, Z.X.; Xu, S.W. Oxidative stress-mediated cytotoxicity of cadmium in chicken splenic lymphocytes. Toxicol. Lett. 2010, 196, S122. [Google Scholar] [CrossRef]
- Zheng, J.L.; Yuan, S.S.; Wu, C.W.; Li, W.Y. Chronic waterborne zinc and cadmium exposures induced different responses towards oxidative stress in the liver of zebrafish. Aquat. Toxicol. 2016, 177, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.L.; Yuan, S.S.; Wu, C.W.; Lv, Z.M. Acute exposure to waterborne cadmium induced oxidative stress and immunotoxicity in the brain, ovary and liver of zebrafish (Danio rerio). Aquat. Toxicol. 2016, 180, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; An, K.; Xie, L.; Wu, W.; Zhang, R.; Wang, D.; Fang, Y.; Chen, H.; Xiao, S.; Fang, L. Transmissible gastroenteritis virus infection induces NF-kappab activation through RLR-mediated signaling. Virology 2017, 507, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.; Petznick, A.; Heryati, S.; Rifada, M.; Tong, L. Nuclear factor-kappab: Central regulator in ocular surface inflammation and diseases. Ocul. surf. 2012, 10, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Sun, Y.; Song, W.; Xu, T. Microrna-145 regulates the rlr signaling pathway in miiuy croaker after poly(I:C) stimulation via targeting MDA5. Dev. Comp. Immunol. 2017, 68, 79–86. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds APS and CdCl2 are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment |
|---|---|
| control group | no treatment |
| Cd group | incubating with CdCl2 (10−6 mol/L, Guangfu Technology Co,. Ltd., Tianjin, China) for 24 h |
| APS + Cd group | incubating with APS (40 mg/L) for 2 h first, then adding CdCl2 (10−6 mol/L) for 24 h |
| anti-MDA5 mAb + Cd group | incubating with anti-MDA5 mAb (5 mg/L) for 2 h first, then adding CdCl2 (10−6 mol/L) for 24 h |
| BAY + Cd group | incubating with BAY11-7082 (10−4 mol/L, Beyotime, Haimen, China) for 2 h first, then adding CdCl2 (10−6 mol/L) for 24 h |
| 10 mg/L APS group | incubating with 10 mg/L APS for 24 h |
| 20 mg/L APS group | incubating with 20 mg/L APS for 24 h |
| 40 mg/L APS group | incubating with 40 mg/L APS for 24 h |
| anti-MDA5 mAb group | incubating with anti-MDA5 mAb (5 mg/L) for 24 h |
| BAY group | incubating with BAY11-7082 for 24 h |
| Primer Name | Primer Sequence | Primer Length (bp) | |
|---|---|---|---|
| MDA5 | Forward | TCAGGAGGAGGACGACCACGAT | 22 |
| Reverse | TTCCCACGACTCTCAATAACAG | 22 | |
| IPS-1 | Forward | TGCAGGGAGGCCATACACCAGTG | 23 |
| Reverse | TCCACCTCCCAAGGTGACCCGTG | 23 | |
| NF-κB | Forward | TCTGAACAGCAAGTCATCCATAACG | 25 |
| Reverse | AAGGAAGTGAGGTTGAGGAGTCG | 23 | |
| IL-1β | Forward | TTCCGCTACACCCGCTCACAGT | 22 |
| Reverse | CCGCTCATCACACACGACAT | 20 | |
| IL-6 | Forward | ATGGTGATAAATCCCGATGAAG | 22 |
| Reverse | CCTCACGGTCTTCTCCATAAAC | 22 | |
| TNF-α | Forward | CAGATGGGAAGGGAATGAAC | 20 |
| Reverse | AGAGCATCAACGCAAAAGGG | 20 | |
| β-actin | Forward | ATTGCTGCGCTCGTTGTT | 18 |
| Reverse | CTTTTGCTCTGGGCTTCA | 18 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, W.; Ge, M.; Li, G.; Zhang, L.; Tang, Z.; Li, R.; Zhang, R. Astragalus Polysaccharide Protect against Cadmium-Induced Cytotoxicity through the MDA5/NF-κB Pathway in Chicken Peripheral Blood Lymphocytes. Molecules 2017, 22, 1610. https://doi.org/10.3390/molecules22101610
Xie W, Ge M, Li G, Zhang L, Tang Z, Li R, Zhang R. Astragalus Polysaccharide Protect against Cadmium-Induced Cytotoxicity through the MDA5/NF-κB Pathway in Chicken Peripheral Blood Lymphocytes. Molecules. 2017; 22(10):1610. https://doi.org/10.3390/molecules22101610
Chicago/Turabian StyleXie, Wanqiu, Ming Ge, Guangxing Li, Linan Zhang, Zequn Tang, Ruyue Li, and Ruili Zhang. 2017. "Astragalus Polysaccharide Protect against Cadmium-Induced Cytotoxicity through the MDA5/NF-κB Pathway in Chicken Peripheral Blood Lymphocytes" Molecules 22, no. 10: 1610. https://doi.org/10.3390/molecules22101610
APA StyleXie, W., Ge, M., Li, G., Zhang, L., Tang, Z., Li, R., & Zhang, R. (2017). Astragalus Polysaccharide Protect against Cadmium-Induced Cytotoxicity through the MDA5/NF-κB Pathway in Chicken Peripheral Blood Lymphocytes. Molecules, 22(10), 1610. https://doi.org/10.3390/molecules22101610