Triterpenoids from Ocimum labiatum Activates Latent HIV-1 Expression In Vitro: Potential for Use in Adjuvant Therapy

 ,
,
Abstract
:1. Introduction
2. Results
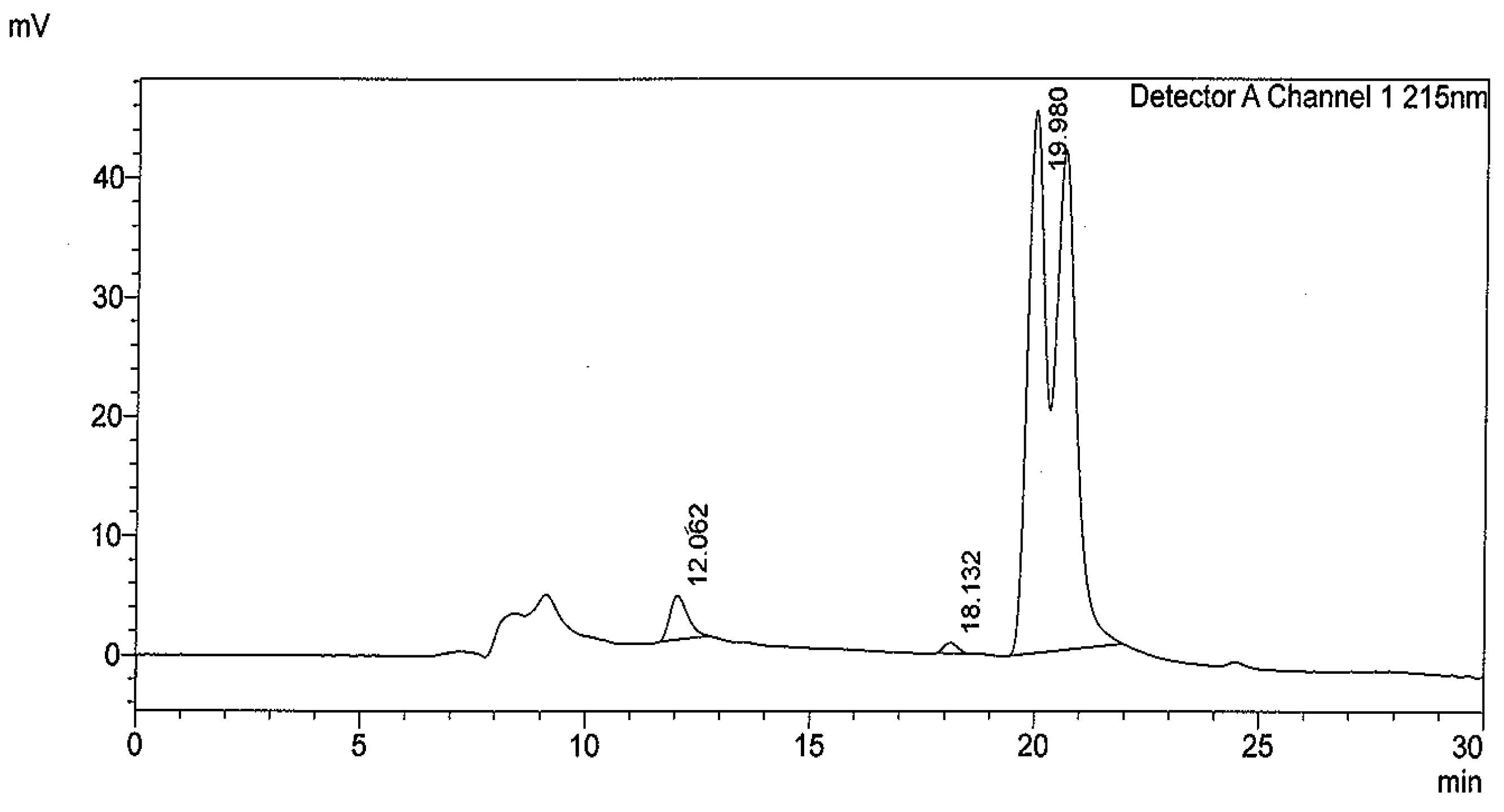
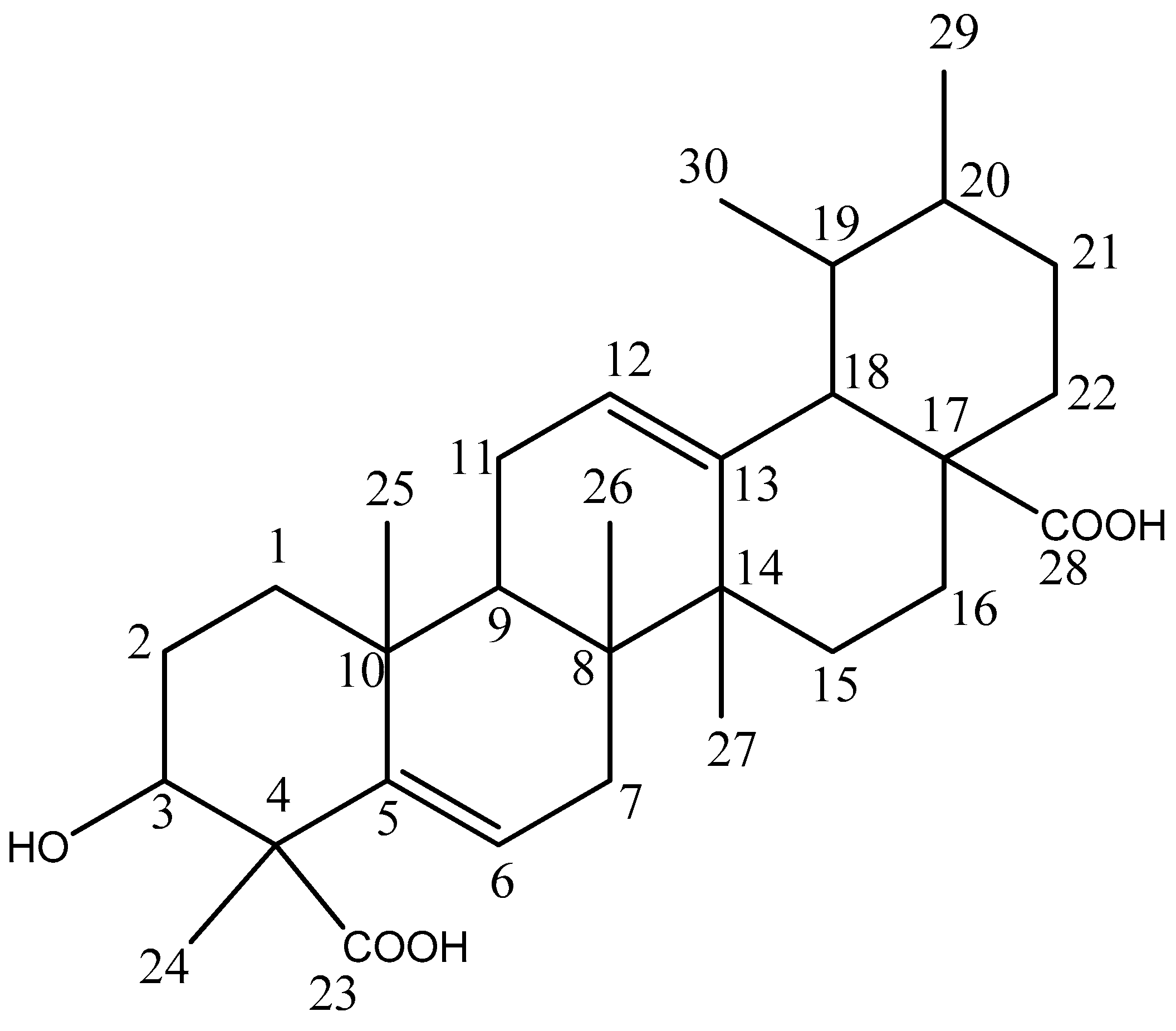
2.1. Compounds Isolation
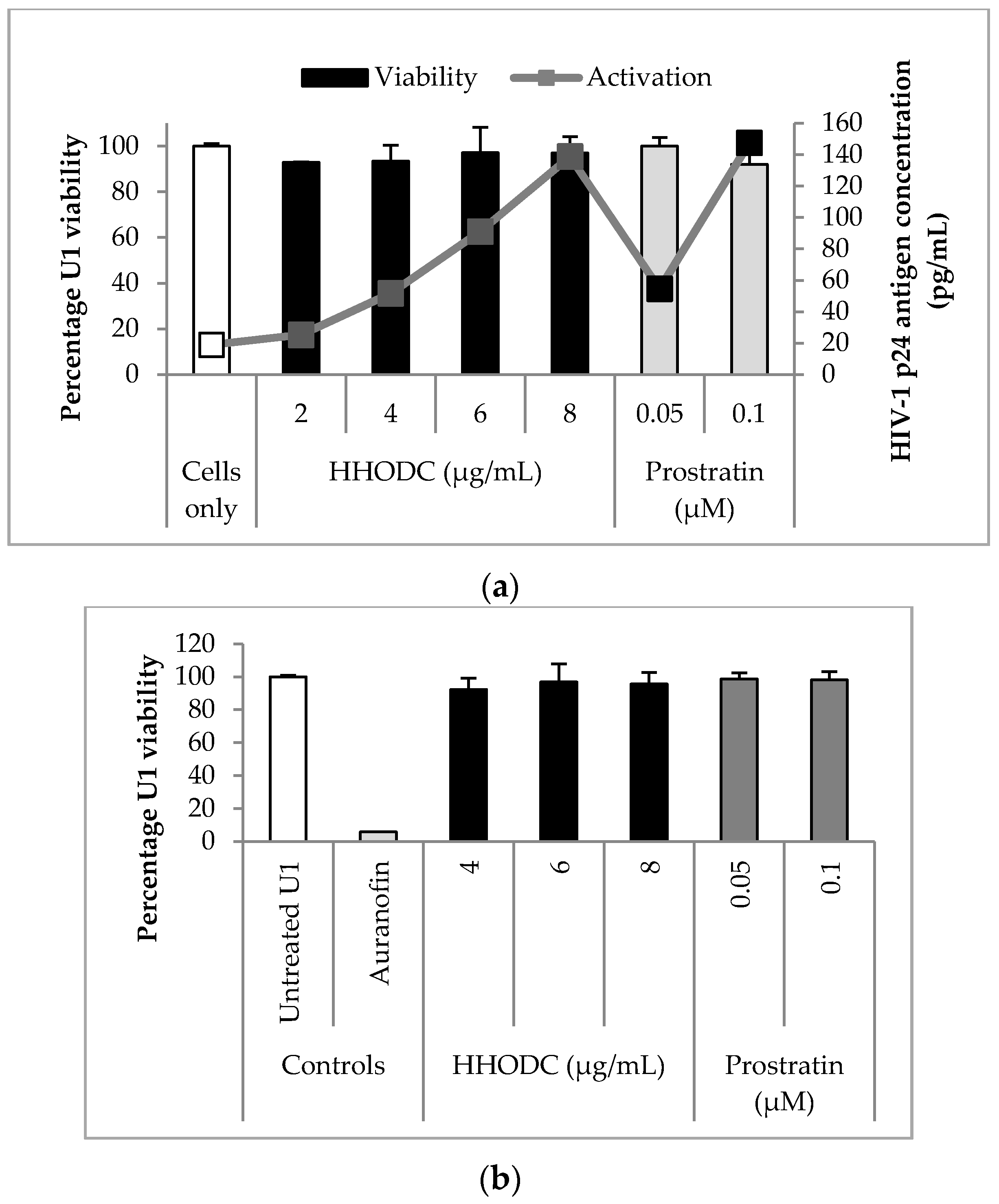
2.2. Viability Studies
2.3. Effect of HHODC on Viral Expression
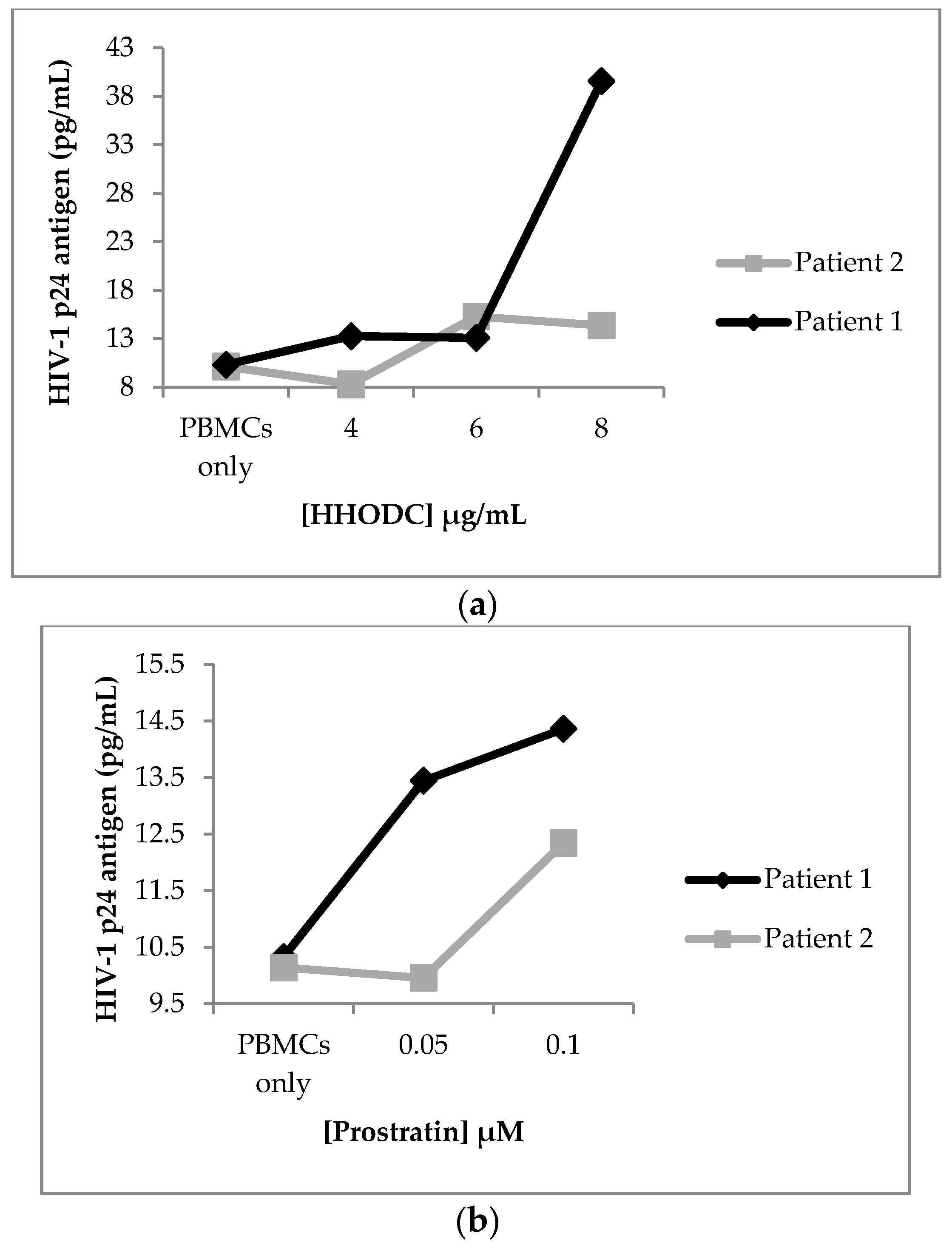
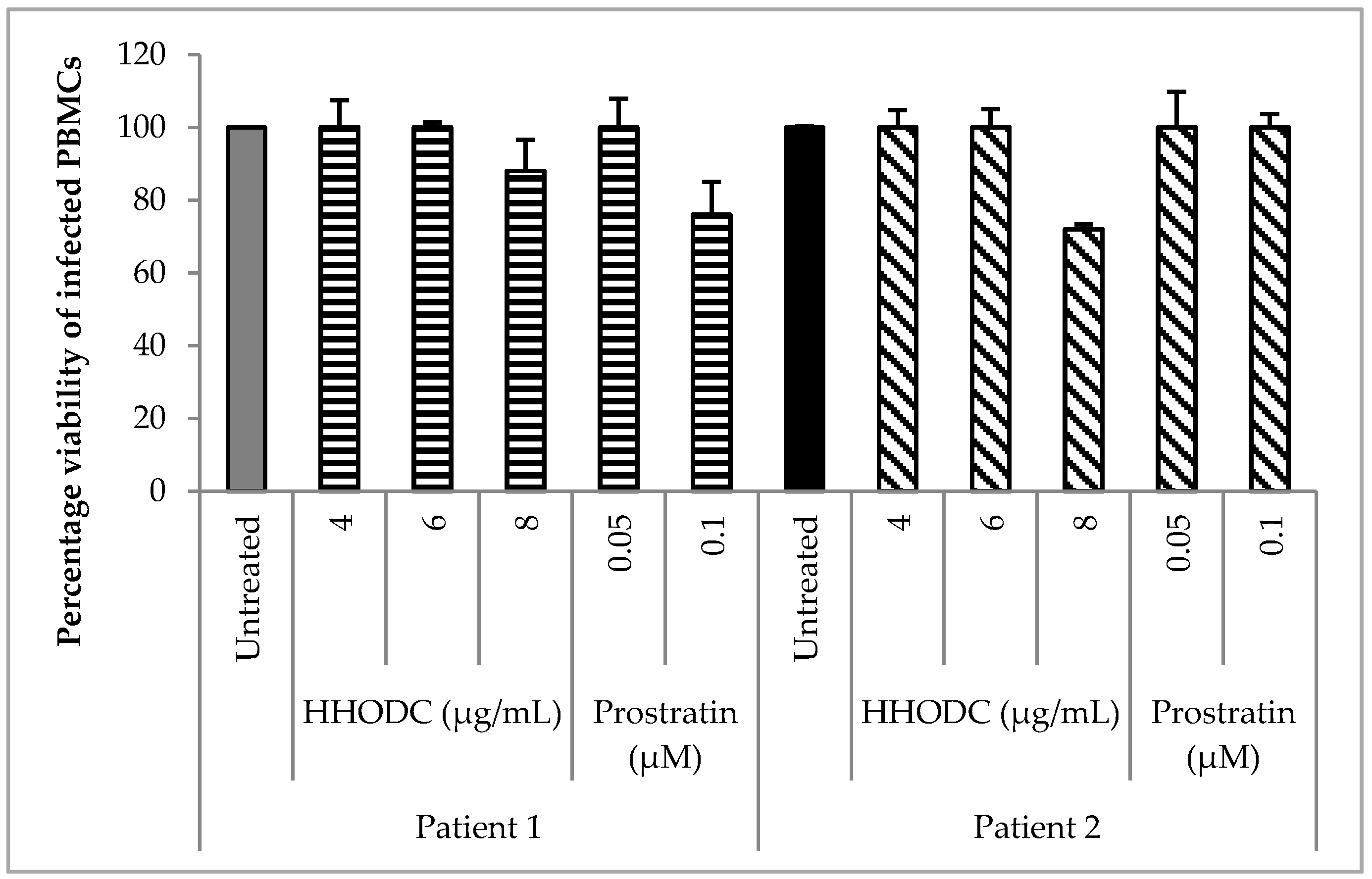
2.4. Effect of HHODC on Natural Infection
2.5. Viral Reactivation Mechanism of HHODC
2.6. Effect of HHODC on the Endogenous Production of Pro-Inflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. General
4.2. Plant Material
4.3. Isolation and Identification of Compounds
4.4. Cytotoxicity of Isolated Compounds
4.5. Stimulation of Latent HIV-1 Production
4.5.1. HIV-1 Expression from Latently Infected Promonocytic U1 Cells
4.5.2. HIV-1 Expression from PBMCs of Infected Individuals on cART
4.6. Viral Reactivating Mechanism
4.6.1. Effect of HHODC on Histone Deacetylase Activity
4.6.2. Effect of HHODC on Protein Kinase C Activity
4.6.3. Effect of HHODC on the Production of Pro-inflammatory Cytokines
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hodel, F.; Patxot, M.; Snäkä, T.; Ciuffi, A. HIV-1 latent reservoir: Size matters. Future Virol. 2016, 11, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Kulkosky, J.; Bray, S. HAART-persistent HIV-1 latent reservoirs: Their origin, mechanisms of stability and potential strategies for eradication. Curr. HIV Res. 2006, 4, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.A.; Sholtis, K.; Kirchherr, J.; Kuruc, J.D.; Gay, C.L.; Nordstrom, J.L.; Bollard, C.M.; Archin, N.M.; Margolis, D.M. Vorinostat Renders the Replication-Competent Latent Reservoir of Human Immunodeficiency Virus (HIV) Vulnerable to Clearance by CD8 T Cells. EBioMedicine 2017. [Google Scholar] [CrossRef] [PubMed]
- Marcello, A. Latency: The hidden HIV-1 challenge. Retrovirology 2006, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Palmer, S.; Josefsson, L.; Coffin, J.M. HIV reservoirs and the possibility of a cure for HIV infection. J. Intern. Med. 2011, 270, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, M.; Bukrinsky, M. Human immunodeficiency virus (HIV) latency: The major hurdle in HIV eradication. Mol. Med. 2012, 18, 1096–1108. [Google Scholar] [CrossRef] [PubMed]
- Archin, N.M.; Sung, J.M.; Garrido, C.; Soriana-Sarabia, N.; Margolis, D.M. Eradicating HIV-1 infection: Seeking to clear a persistent pathogen. Nat. Rev. Microbiol. 2014, 12, 750–764. [Google Scholar] [CrossRef] [PubMed]
- Gallastegui, E.; Marshall, B.; Vidal, D.; Sanchez-Duffhues, G.; Collado, J.A.; Alvarez-Fernández, C.; Luque, N.; Terme, J.-M.; Gatell, J.M.; Sánchez-Palomino, S.; et al. Combination of biological screening in a cellular model of viral latency and virtual screening identifies novel compounds that reactivate HIV-1. J. Virol. 2012, 86, 3795–3808. [Google Scholar] [CrossRef] [PubMed]
- Darcis, G.; Van Driessche, B.; Van Lint, C. HIV Latency: Should We Shock or Lock? Trends Immunol. 2017, 38, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Frick, M.; Gaudino, A.; Harrington, M.; Horn, T.; Jefferys, R.; Johnson, J.; Lessem, E.; Low, M.; McKenna, L. 2017 Pipeline Report. HIV, TB & HCV: Drugs, Diagnostics, Vaccines, Preventive Technologies, Cure Research, and Immune-Based and Gene Therapies in Development; Treatment Action Group (TAG): New York, NY, USA, 2017. [Google Scholar]
- Kulkosky, J.; Culnan, D.M.; Roman, J.; Dornadula, G.; Schnell, M.; Boyd, M.R.; Pomerantz, R.J. Prostratin: Activation of latent HIV-1 expression suggests a potential inductive adjuvant therapy for HAART. Blood 2001, 98, 3006–3015. [Google Scholar] [CrossRef] [PubMed]
- Rullas, J.; Bermejo, M.; García-Pérez, J.; Beltán, M.; González, N.; Hezareh, M.; Brown, S.J.; Alcamí, J. Prostratin induces HIV activation and downregulates HIV receptors in peripheral blood lymphocytes. Antivir. Ther. 2004, 9, 545–554. [Google Scholar] [PubMed]
- Pettit, G.R.; Herald, C.L.; Doubek, D.L.; Herald, D.L. Isolation and structure of bryostatin 1. J. Am. Chem. Soc. 1982, 104, 6846–6848. [Google Scholar] [CrossRef]
- Marsden, M.D.; Zack, J.A. HIV/AIDS eradication. Bioorg. Med. Chem. Lett. 2013, 23, 4003–4010. [Google Scholar] [CrossRef] [PubMed]
- DeChristopher, B.A.; Loy, B.A.; Marsden, M.D.; Schrier, A.J.; Zack, J.A.; Wender, P.A. Designed, synthetically accessible bryostatin analogues potently induce activation of latent HIV reservoirs in vitro. Nat. Chem. 2012, 4, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Wender, P.A.; Nakagawa, Y.; Near, K.E.; Staveness, D. Computer-guided design, synthesis, and protein kinase C affinity of a new salicylate-based class of Bryostatin analogs. Org. Lett. 2014, 16, 5136–5139. [Google Scholar] [CrossRef] [PubMed]
- Darcis, G.; Kula, A.; Bouchat, S.; Fujinaga, K.; Corazza, F.; Ait-Ammar, A.; Delacourt, N.; Melard, A.; Kabeya, K.; Vanhulle, C.; et al. An In-Depth Comparison of Latency-Reversing Agent Combinations in Various In Vitro and Ex Vivo HIV-1 Latency Models Identified Bryostatin-1+JQ1 and Ingenol-B+JQ1 to Potently Reactivate Viral Gene Expression. PLoS Pathog. 2015, 11, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.J.; Brits, G.J.; Potgieter, C.J.; Staden, J. Van Plectranthus: A plant for the future? S. Afr. J. Bot. 2011, 77, 947–959. [Google Scholar] [CrossRef]
- Van Wyk, B.-E.; Van Oudtshoorn, B.; Gericke, N. Medicinal Plants of South Africa; Briza Publications: Pretoria, South Africa, 2009. [Google Scholar]
- Macías, F.A.; Lacret, R.; Varela, R.M.; Nogueiras, C.; Molinillo, J.M.G. Bioactive apocarotenoids from Tectona grandis. Phytochemistry 2008, 69, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.A.; Meyer, J.J.M.; Jimeno, L. Bioactive diterpenes from Orthosiphon labiatus and Salvia africana-lutea. J. Nat. Prod. 2007, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Ma, C.; Chen, D.; Hattori, M. Anti-HIV-1 protease triterpenoids from Stauntonia obovatifoliola Hayata subsp. Intermedia. Phytochemistry 2008, 69, 1875–1879. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.F.; Ogundele, A.; Forrest, A.; Wilton, J.; Salzwedel, K.; Doto, J.; Allaway, G.P.; Martin, D.E. Phase I and II study of the safety, virologic effect, and pharmacokinetics/pharmacodynamics of single-dose 3-O-(3’,3’-dimethylsuccinyl) betulinic acid (bevirimat) against human immunodeficiency virus Infection. Antimicrob. Agents Chemother. 2007, 51, 3574–3581. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, C.A.; Joshi, P.; Sloan, B.; Bare, J.C.; Smith, P.C.; Allaway, G.P.; Wild, C.T.; Martin, D.E. Potent activity of the HIV-1 maturation inhibitor bevirimat in SCID-hu Thy/Liv mice. PLoS ONE 2007, 2, e1251. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Goila-Gaur, R.; Salzwedel, K.; Kilgore, N.R.; Reddick, M.; Matallana, C.; Castillo, A.; Zoumplis, D.; Martin, D.E.; Orenstein, J.M.; et al. PA-457: A potent HIV inhibitor that disrupts core condensation by targeting a late step in Gag processing. Proc. Natl. Acad. Sci. USA 2003, 100, 13555–13560. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.; Ding, J.; Yu, S.; Wu, T.; Zhang, Q.; Liang, F. Progress and challenges in the use of latent HIV-1 reactivating agents. Acta Pharmacol. Sin. 2015, 36, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Kushiro, T.; Shibuya, M.; Ebizuka, Y. Beta-amyrin synthase-cloning of oxidosqualene cyclase that catalyzes the formation of the most popular triterpene among higher plants. Eur. J. Bochem. 1998, 256, 238–244. [Google Scholar] [CrossRef]
- Zhou, C.-X.; Zou, L.; Gan, L.-S.; Cao, Y.-L. Kleinhospitines A-D, new cycloartane triterpenoid alkaloids from Kleinhovia hospita. Org. Lett. 2013, 15, 2734–2737. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, D.; Wong, S. Carboxyfluorescein succinimidyl ester-based proliferative assays for assessment of T cell function in the diagnostic laboratory. Immunol. Cell Biol. 1999, 77, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Gianella, S.; Taylor, J.; Brown, T.R.; Kaytes, A.; Achim, C.L.; Moore, D.J.; Little, S.J.; Ellis, R.J.; Smith, D.M. Can research at the end of life be a useful tool to advance HIV cure? AIDS 2017, 31, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Pinkevych, M.; Cromer, D.; Tolstrup, M.; Grimm, A.J.; Cooper, D.A.; Lewin, S.R.; Søgaard, O.S.; Rasmussen, T.A.; Kent, S.J.; Kelleher, A.D.; et al. HIV Reactivation from Latency after Treatment Interruption Occurs on Average Every 5–8 Days—Implications for HIV Remission. PLoS Pathog. 2015, 11, e1005000. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.A.; Chaves, M.H.; Almeida, F.R.C.; Lima, R.C.P.; Silva, R.M.; Maia, J.L.; Brito, G.A.A.C.; Santos, F.A.; Rao, V.S. Protective effect of alpha- and beta-amyrin, a triterpene mixture from Protium heptaphyllum (Aubl.) March. trunk wood resin, against acetaminophen-induced liver injury in mice. J. Ethnopharmacol. 2005, 98, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Dandekar, S. Targeting NF-κB Signaling with Protein Kinase C Agonists As an Emerging Strategy for Combating HIV Latency. AIDS Res. Hum. Retrovirus 2015, 31, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Vigushin, D.M.; Ali, S.; Pace, P.E.; Mirsaidi, N.; Ito, K.; Adcock, I.; Coombes, R.C. Trichostatin A is a histone deacetylase inhibitor with potent antitumor activity against breast cancer in vivo. Clin. Cancer Res. 2001, 7, 971–976. [Google Scholar] [PubMed]
- McManamy, M.; Hakre, S.; Verdin, E.; Margolis, D. Therapy for latent human immunodeficiency virus type 1 infection: The role of histone deacetylase inhibitors. Antivir. Chem. Chemother. 2015, 23, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Archin, N.M.; Liberty, A.L.; Kashuba, A.D.; Choudhary, S.K.; Kuruc, J.D.; Crooks, A.M.; Parker, D.C.; Anderson, E.M.; Kearney, M.F.; Strain, M.C.; et al. Administration of vorinostat disrupts HIV-1 latency in patients on antiretroviral therapy. Nature 2012, 487, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.G.; Chiang, V.; Fyne, E.; Balakrishnan, M.; Barnes, T.; Graupe, M.; Hesselgesser, J.; Irrinki, A.; Murry, J.P.; Stepan, G.; et al. Histone Deacetylase Inhibitor Romidepsin Induces HIV Expression in CD4 T Cells from Patients on Suppressive Antiretroviral Therapy at Concentrations Achieved by Clinical Dosing. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaki, E.; Hiraga, N.; Imamura, M.; Uchida, T.; Kan, H.; Tsuge, M.; Abe-Chayama, H.; Hayes, C.N.; Makokha, G.N.; Serikawa, M.; et al. Interferon alpha treatment stimulates interferon gamma expression in type I NKT cells and enhances their antiviral effect against hepatitis C virus. PLoS ONE 2017, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wei, Y.; Sun, Y.; Shi, W.; Yang, J.; Zhu, L.; Li, M. Interferon-gamma inhibits melanogenesis and induces apoptosis in melanocytes: A pivotal role of CD8+ cytotoxic T lymphocytes in vitiligo. Acta Derm. Venereol. 2015, 95, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.M.; Lee, E.J.; Bramante, J.M.; Barker, E.; Wilson, C.C. The natural killer cell interferon-gamma response to bacteria is diminished in untreated HIV-1 infection and defects persist despite viral suppression. J. Acquir. Immune Defic. Syndr. 2014, 1, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, P.; Meyer, D. In vitro reactivation of latent HIV-1 by cytostatic bis(thiosemicarbazonate) gold(III) complexes. BMC Infect. Dis. 2014, 14, 680. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Steffens, F.; Reinecke, C.; Meyer, D. The Th1/Th2/Th17 cytokine profile of HIV-infected individuals: A multivariate cytokinomics approach. Cytokine 2013, 61, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Worsley, C.M.; Suchard, M.S.; Stevens, W.S.; Van Rie, A.; Murdoch, D.M. Multi-analyte profiling of ten cytokines in South African HIV-infected patients with Immune Reconstitution Inflammatory Syndrome (IRIS). AIDS Res. Ther. 2010, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Nixon, D.E.; Landay, A.L. Biomarkers of immune dysfunction in HIV. Curr. Opin. HIV AIDS 2010, 5, 498–503. [Google Scholar] [CrossRef] [PubMed]
- French, M.A.; Cozzi-Lepri, A.; Arduino, R.C.; Johnson, M.; Achhra, A.C.; Landay, A. Plasma levels of cytokines and chemokines and the risk of mortality in HIV-infected individuals. Aids 2015, 29, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Kinter, A.L.; Fauci, A.S. Interleukin 1 induces expression of the human immunodeficiency virus alone and in synergy with interleukin 6 in chronically infected U1 cells: Inhibition of inductive effects by the interleukin 1 receptor antagonist. Proc. Natl. Acad. Sci. USA 1994, 91, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.; Poli, G.; Kinter, A.L.; Justement, J.S.; Stanley, S.K.; Maury, W.J.; Bressler, P.; Orenstein, J.M.; Fauci, A.S. Interferon ɣ induces the expression of human immunodeficiency virus in persistently infected promonocytic cells (U1) and redirects the production of virions to intracytoplasmic vacuoles in phorbol myristate acetate-differentiated U1 cells. J. Exp. Med. 1992, 176, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Kalebic, T.; Kinter, A.; Poli, G.; Anderson, M.E.; Meister, A.; Fauci, A.S. Suppression of human immunodeficiency virus expression in chronically infected monocytic cells by glutathione, glutathione ester, and N-acetylcysteine. Proc. Natl. Acad. Sci. USA 1991, 88, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Folks, T.M.; Justement, J.; Kinter, A.; Dinarello, C.A.; Fauci, A.S. Cytokine-induced expression of HIV-1 in a chronically infected promonocyte cell line. Science 1987, 238, 800–802. [Google Scholar] [CrossRef] [PubMed]
- Garrelds, I.M.; Van Hal, P.T.; Haakmat, R.C.; Hoogsteden, H.C.; Saxena, P.R.; Zijlstra, F.J. Time dependent production of cytokines and eicosanoids by human monocytic leukaemia U937 cells; effects of glucocorticosteroids. Mediat. Inflamm. 1999, 8, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Kapewangolo, P.; Hussein, A.A.; Meyer, D. Inhibition of HIV-1 enzymes, antioxidant and anti-inflammatory activities of Plectranthus barbatus. J. Ethnopharmacol. 2013, 149, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Martelanc, M.; Vovk, I.; Simonovska, B. Separation and identification of some common isomeric plant triterpenoids by thin-layer chromatography and high-performance liquid chromatography. J. Chromatogr. A 2009, 1216, 6662–6670. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Kapewangolo, P.; Omolo, J.J.; Bruwer, R.; Fonteh, P.; Meyer, D. Antioxidant and anti-inflammatory activity of Ocimum labiatum extract and isolated labdane diterpenoid. J. Inflamm. 2015, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Siliciano, R.F. Targeting HIV latency: Pharmacologic strategies toward eradication. Drug Discov. Today 2013, 18, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.A.; Søgaard, O.S.; Brinkmann, C.; Wightman, F.; Lewin, S.R.; Melchjorsen, J.; Dinarello, C.; Østergaard, L.; Tolstrup, M. Comparison of HDAC inhibitors in clinical development. Effect on HIV production in latently infected cells and T-cell activation. Hum. Vacc. Immunother. 2013, 9, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Wegener, D.; Wirsching, F.; Riester, D.; Schwienhorst, A. A fluorogenic histone deacetylase assay well suited for high-throughput activity screening. Chem. Biol. 2003, 10, 61–68. [Google Scholar] [CrossRef]
- Poli, G.; Bressler, P.; Kinter, A.; Duh, E.; Timmer, W.C.; Rabson, A.; Justement, J.S.; Stanley, S.; Fauci, A.S. Interleukin 6 induces human immunodeficiency virus expression in infected monocytic cells alone and in synergy with tumor necrosis factor alpha by transcriptional and post-transcriptional mechanisms. J. Exp. Med. 1990, 172, 151–158. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds, HHODC and amyrin, are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine Concentrations (pg/mL) | ||||
|---|---|---|---|---|
| PBMCs | U1 Cells | |||
| Analyte | Untreated | Treated with HHODC | Untreated | HHODC |
| IL-2 | 21.0 ± 6.4 | 26.9 ± 6.9 | 0 | 2.01 ± 5.4 |
| IL-6 | 2801.7 ± 21.9 | 14744.9 ± 17.9 | 1.4 ± 6.3 | 1.6 ± 4.6 |
| TNF-α | 164.1 ± 26.5 | 528.7 ± 27.7 | 1.7 ± 4.4 | 2.2 ± 4.9 |
| IFN-γ | 251.0 ± 22.9 | 842.6 ± 21.0 | 2.5 ± 4.6 | 1.8 ± 4.7 |
| Position | 13C δ ppm | 1H δ mult |
|---|---|---|
| 1 | 36.7 (t) | 1.41, 2H, (m) |
| 2 | 27.3 (t) | 1.61, 2H, (m) |
| 3 | 78.3 (d) | 2.89, 1H (m) |
| 4 | 55.3 (s) | |
| 5 | 143.8 (s) | |
| 6 | 122.2 (d) | 5.26, 1H, (m) |
| 7 | 33.9 (t) | 2.02, 2H, (s) |
| 8 | 39.0 (s) | |
| 9 | 47.8 (d) | 1.54, 1H, (m) |
| 10 | 30.2 (s) | |
| 11 | 26.4 (t) | 2.15, 2H, (m) |
| 12 | 125.4 (d) | 5.25, 1H, (m) |
| 13 | 138.2 (s) | |
| 14 | 41.3 (s) | |
| 15 | 27.8 (t) | 1.35, 2H |
| 16 | 25.0 (t) | 1.64, 2H |
| 17 | 48.1 (s) | |
| 18 | 52.9 (d) | 2.25, 1H |
| 19 | 38.6 (d) | 1.71, 1H |
| 20 | 38.4 (d) | 1.67, 1H |
| 21 | 30.4 (t) | 1.58, 2H |
| 22 | 33.5 (t) | 1.98, 2H |
| 23 | 180.3 (s) | |
| 24 | 18.1 (q) | 1.18, 3H (s) |
| 25 | 20.1 (q) | 1.14, 3H (s) |
| 26 | 14.9 (q) | 0.93, 3H (s) |
| 27 | 23.9 (q) | 1.00, 3H (s) |
| 28 | 180.5 (s) | |
| 29 | 23.1 (q) | 0.80, 3H |
| 30 | 16.3 (q) | 0.87, 3H |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapewangolo, P.; Omolo, J.J.; Fonteh, P.; Kandawa-Schulz, M.; Meyer, D. Triterpenoids from Ocimum labiatum Activates Latent HIV-1 Expression In Vitro: Potential for Use in Adjuvant Therapy. Molecules 2017, 22, 1703. https://doi.org/10.3390/molecules22101703
Kapewangolo P, Omolo JJ, Fonteh P, Kandawa-Schulz M, Meyer D. Triterpenoids from Ocimum labiatum Activates Latent HIV-1 Expression In Vitro: Potential for Use in Adjuvant Therapy. Molecules. 2017; 22(10):1703. https://doi.org/10.3390/molecules22101703
Chicago/Turabian StyleKapewangolo, Petrina, Justin J. Omolo, Pascaline Fonteh, Martha Kandawa-Schulz, and Debra Meyer. 2017. "Triterpenoids from Ocimum labiatum Activates Latent HIV-1 Expression In Vitro: Potential for Use in Adjuvant Therapy" Molecules 22, no. 10: 1703. https://doi.org/10.3390/molecules22101703
APA StyleKapewangolo, P., Omolo, J. J., Fonteh, P., Kandawa-Schulz, M., & Meyer, D. (2017). Triterpenoids from Ocimum labiatum Activates Latent HIV-1 Expression In Vitro: Potential for Use in Adjuvant Therapy. Molecules, 22(10), 1703. https://doi.org/10.3390/molecules22101703