An Efficient PCR-RFLP Method for the Rapid Identification of Korean Pyropia Species


Abstract
:1. Introduction
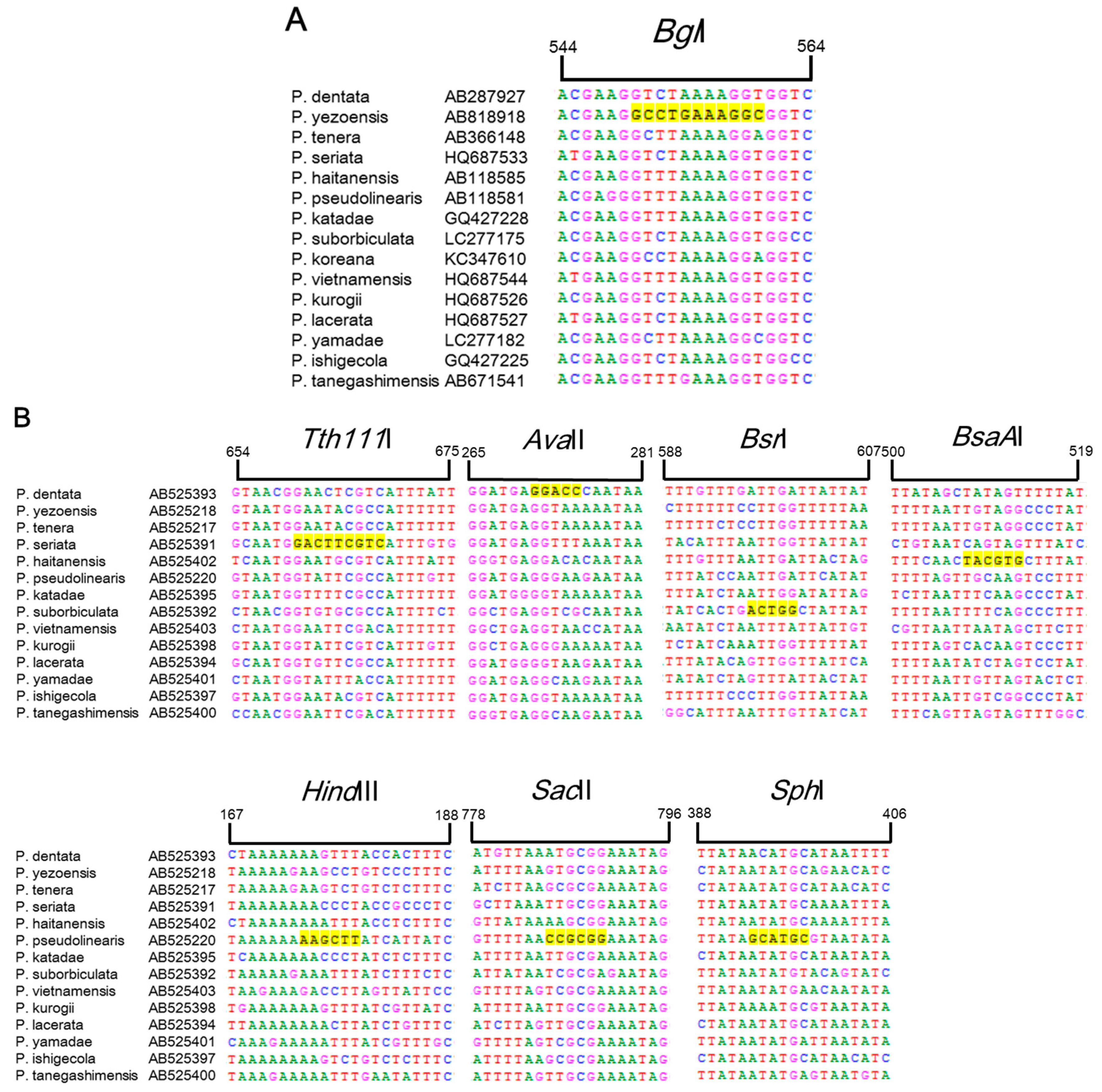
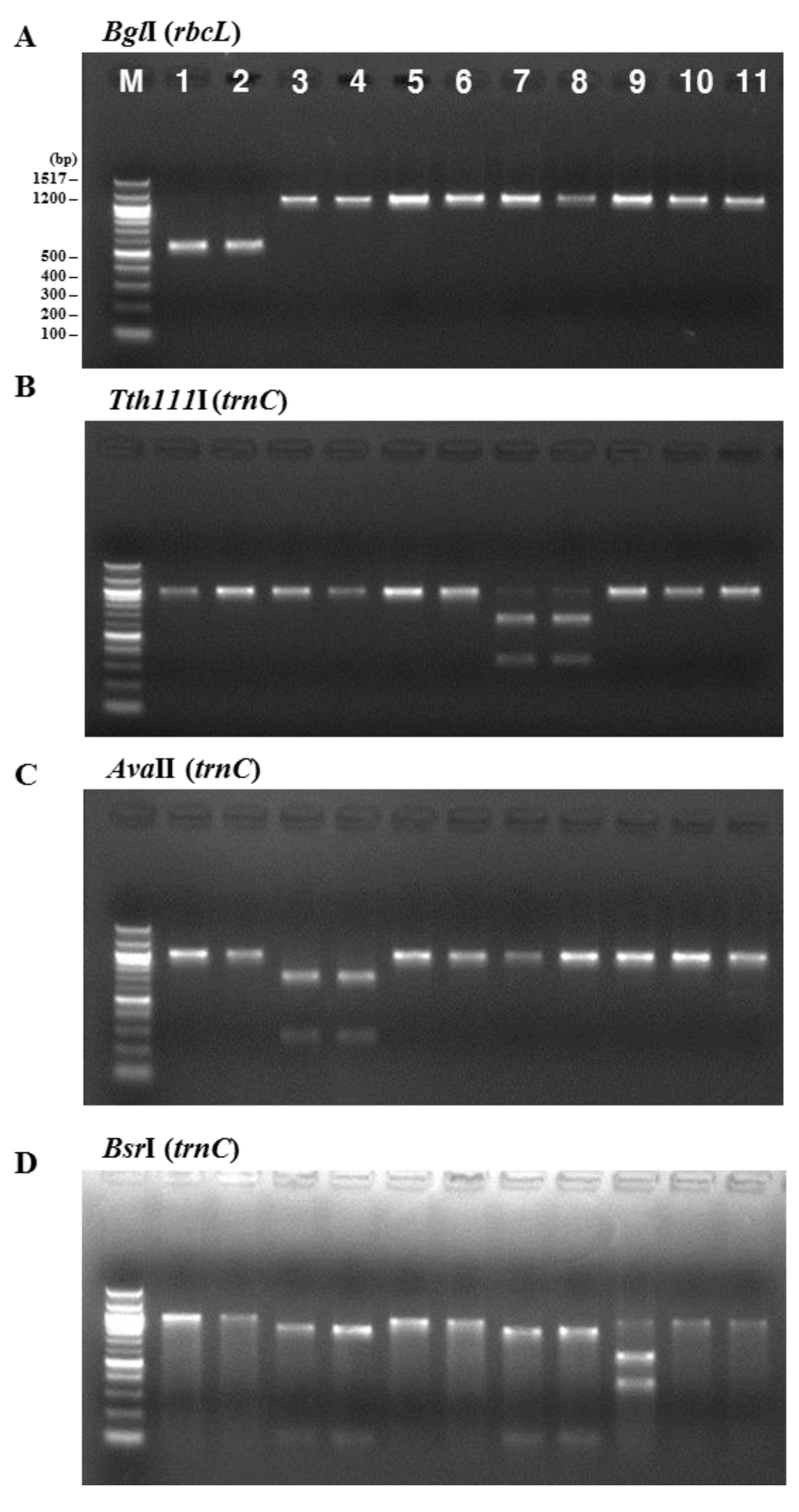
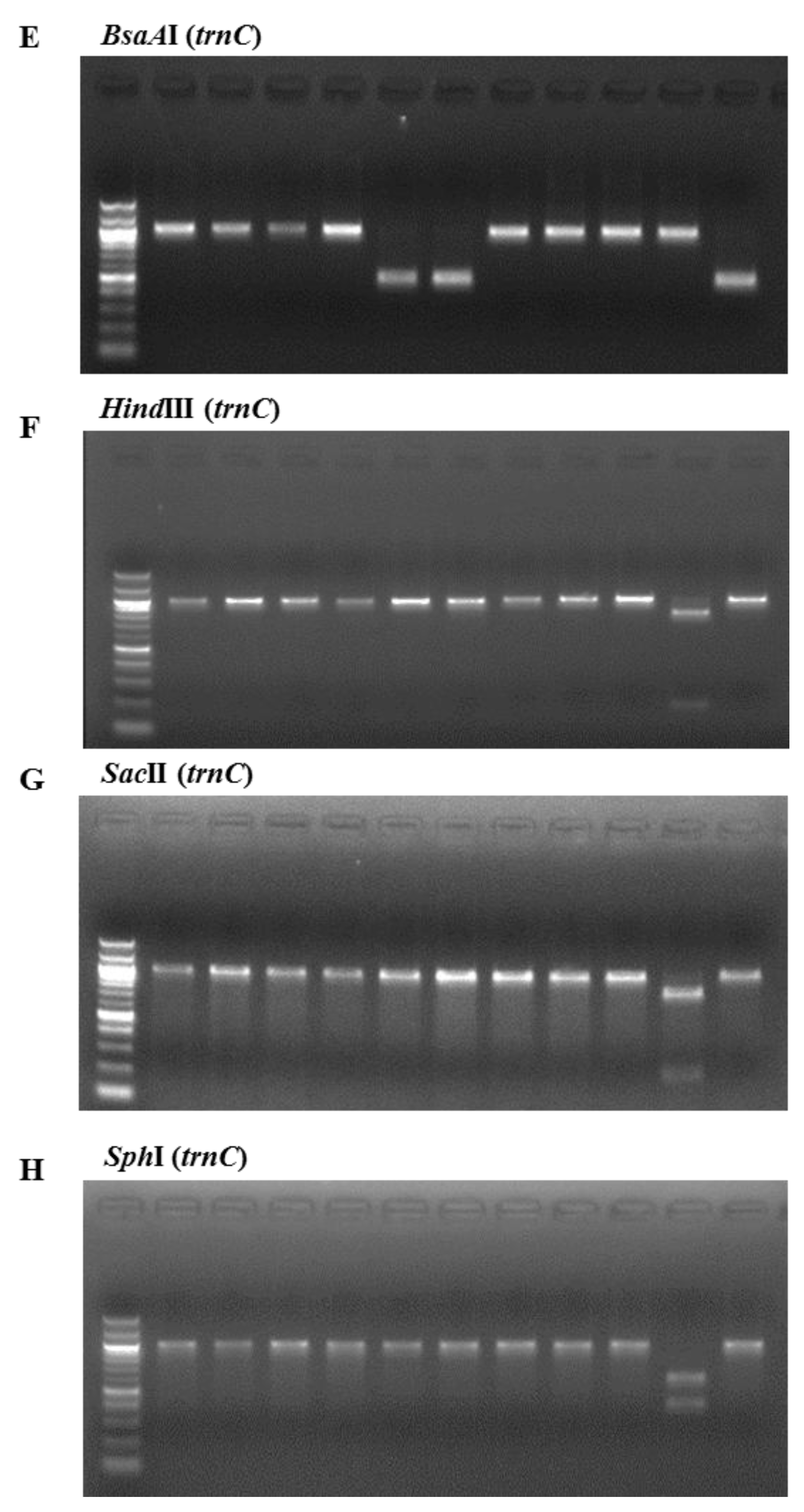
2. Results and Discussion
3. Materials and Methods
3.1. Sequence Analysis
3.2. Sample Collection and DNA Isolation
3.3. PCR Amplification and Restriction Digestion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vergés, A.; Sánchez, N.; Peteiro, C.; Polo, L.; Brodie, J. Pyropia suborbiculata (Bangiales, Rhodophyta): First records from the northeastern Atlantic and Mediterranean of this North Pacific species. Phycologia 2013, 52, 121–129. [Google Scholar] [CrossRef]
- Hallmann, A. Algae biotechnology–green cell-factories on the rise. Curr. Biotechnol. 2015, 4, 389–415. [Google Scholar] [CrossRef]
- K-Fish Information Portal. Statistical Database for Seasoned Laver Export. Available online: http://www.kfishinfo.net (accessed on 25 November 2017).
- Ock, Y.S. The research on the development steps and facing problems of Korean Japanese laver industry. J. Fish. Bus. Adm. 2011, 42, 113–130. (In Korean) [Google Scholar]
- Hwang, M.-S.; In-Gyu, L. Character analysis and numerical taxonomy of Porphyra (Bangiales, Rhodophyta) from Korea. Algae 2002, 17, 217–233. [Google Scholar] [CrossRef]
- Xie, C.; Li, B.; Xu, Y.; Ji, D.; Chen, C. Characterization of the global transcriptome for Pyropia haitanensis (Bangiales, Rhodophyta) and development of cSSR markers. BMC Genom. 2013, 14, 107. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Rentsch, J.; Hübner, P. PCR−RFLP analysis of mitochondrial DNA: A reliable method for species identification. J. Agric. Food Chem. 1999, 47, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Burgener, M.; Hübner, P.; Lüthy, J. PCR-RFLP analysis of mitochondrial DNA: Differentiation of fish species. LWT-Food Sci. Technol. 2000, 33, 144–150. [Google Scholar] [CrossRef]
- Abe, M.; Kobayashi, M.; Fujiyoshi, E.; Tamaki, M.; Kikuchi, N.; Murase, N. Use of PCR-RFLP for the discrimination of Japanese Porphyra and Pyropia species (Bangiales, Rhodophyta). J. Appl. Phycol. 2013, 25, 225–232. [Google Scholar] [CrossRef]
- Kyosuke, N.; Kobiyama, A.; Aruga, Y. Confirmation of cultivated Porphyra tenera (Bangiales, Rhodophyta) by polymerase chain reaction restriction fragment length polymorphism analyses of the plastid and nuclear DNA. Phycol. Res. 2005, 53, 296–302. [Google Scholar] [CrossRef]
- Niwa, K.; Iida, S.; Kato, A.; Kawai, H.; Kikuchi, N.; Kobiyama, A.; Aruga, Y. Genetic diversity and introgression in two cultivated species (Porphyra yezoensis and Porphyra tenera) and closely related wild species of Porphyra (Bangiales, Rhodophyta). J. Phycol. 2009, 45, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Li, Y.; Song, W.; Wang, Z.; Fu, M.; Li, R.; Zhang, X.; Zhu, M. Discrimination of the common macroalgae (Ulva and Blidingia) in coastal waters of Yellow Sea, northern China, based on restriction fragment-length polymorphism (RFLP) analysis. Harmful Algae 2013, 27, 130–137. [Google Scholar] [CrossRef]
- Lee, S.J.; Hwang, M.S.; Park, M.A.; Baek, J.M.; Ha, D.S.; Lee, J.E.; Lee, S.R. Molecular identification of the algal pathogen Pythium chondricola (Oomycetes) from Pyropia yezoensis (Rhodophyta) using ITS and cox1 markers. Algae 2015, 30, 217. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Tiina, S.; George, M. Predicting plastid marker variation: Can complete plastid genomes from closely related species help? PLoS ONE 2013, 8, e82266. [Google Scholar] [CrossRef]
- Touhata, K.; Namikoshi, A.; Suzuki, T.; Iguchi, J.; Mizusawa, N.; Hara, T.; Imamura, S.; Yabu, T.; Yamashita, Y.; Yamashita, M. Origin identification of dried seaweed product “nori” by PCR–RFLP analysis of Pyropia yezoensis in the internal transcribed spacer ITS-1 region. Fish. Sci. 2013, 79, 865–875. [Google Scholar] [CrossRef]
- Li, W.; Cowley, A.; Uludag, M.; Gur, T.; McWilliam, H.; Squizzato, S.; Park, Y.M.; Buso, N.; Lopez, R. The EMBL-EBI bioinformatics web and programmatic tools framework. Nucleic Acids Res. 2015, 43, W580–W584. [Google Scholar] [CrossRef] [PubMed]
- Sudhir, K.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
Sample Availability: Genomic DNA samples are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Restriction Enzymes | ||||||||
|---|---|---|---|---|---|---|---|---|
| Species | BglI | Tth111I | AvaII | BsrI | BsaAI | HindIII | SacII | SphI |
| P. yezoensis | 556 | - | - | - | - | - | - | - |
| P. seriata | - | 664 333 | - | 119 844 | - | - | - | - |
| P. dentata | - | - | 271 732 | 119 844 | - | - | - | - |
| P. suborbiculata | - | - | - | 600 401 | - | - | - | - |
| P. haitanenesis | - | - | - | - | 510 | - | - | - |
| P. pseudolinearis | - | - | - | - | - | 174 825 | 788 211 | 397 602 |
| No. | Species | Source | Accession Number | |
|---|---|---|---|---|
| rbcL | trnC–trnP | |||
| 1 | P. dentata | National Fisheries University, Shimonoseki, Japan | AB118579 | AB525393 |
| 2 | P. yezoensis | National Research Institute of Fisheries Science, Yokohama, Japan | AB818918 | |
| National Fisheries University, Shimonoseki, Japan | AB525218 | |||
| 3 | P. tenera | Fisheries Technology Institute, Hyogo Prefectural Technology Center for Agriculture, Japan | AB366148 | |
| National Fisheries University, Shimonoseki, Japan | AB525217 | |||
| 4 | P. seriata | Department of Biochemistry, University of Otago, New Zealand | HQ687533 | |
| National Fisheries University, Shimonoseki, Japan | AB525391 | |||
| 5 | P. haitanensis | National Fisheries University, Shimonoseki, Japan | AB118585 | AB525402 |
| 6 | P. pseudolinearis | National Fisheries University, Shimonoseki, Japan | AB118581 | AB525220 |
| 7 | P. katadae | Department of Biology and Food Engineering, Changshu Institute of Technology, China | GQ427228 | |
| National Fisheries University, Shimonoseki, Japan | AB525395 | |||
| 8 | P. suborbiculata | National Fisheries Research Institute, Nagasaki, Japan | LC277175 | |
| National Fisheries University, Shimonoseki, Japan | AB525392 | |||
| 9 | P. koreana | Environmental Sciences, Botany, University of Girona, Girona, Spain | KC347610 | |
| 10 | P. vietnamesis | Department of Biochemistry, University of Otago, New Zealand | HQ687544 | |
| National Fisheries University, Shimonoseki, Japan | AB525403 | |||
| 11 | P. kurogii | Department of Biochemistry, University of Otago, New Zealand | HQ687526 | |
| National Fisheries University, Shimonoseki, Japan | AB525398 | |||
| 12 | P. lacerata | Department of Biochemistry, University of Otago, New Zealand | HQ687527 | |
| National Fisheries University, Shimonoseki, Japan | AB525394 | |||
| 13 | P. yamadae | National Fisheries Research Institute, Nagasaki, Japan | LC277182 | |
| National Fisheries University, Shimonoseki, Japan | AB525401 | |||
| 14 | P. ishigecola | Department of Biology and Food Engineering, Changshu Institute of Technology, China | GQ427225 | |
| National Fisheries University, Shimonoseki, Japan | AB525397 | |||
| 15 | P. tanegashimensis | National Fisheries Research Institute, Nagasaki, Japan | AB671541 | |
| National Fisheries University, Shimonoseki, Japan | AB525400 | |||
| No. | Scientific Name | Common Name | Collection Site | Location |
|---|---|---|---|---|
| 1 | P. yezoensis | Bangsamunuigim | Songji-myeon, Haenam-gun, Jeollanam-do | 34°21′05.92′′ N 126°27′40.76′′ E |
| 2 | Soan-myeon, Wando-gun, Jeollanam-do | 34°08′53.12′′ N 126°41′10.12′′ E | ||
| 3 | P. dentata | Itbadidolgim | Yuldo-dong, Mokpo-si, Jeollanam-do | 34°48′13.22′′ N 126°18′34.88′′ E |
| 4 | Palgeum-myeon, Sinan-gun, Jeollanam-do | 34°46′13.04′′ N 126°10′22.44′′ E | ||
| 5 | Uisin-myeon, Jindo-gun, Jeollanam-do | 34°19′07.66′′ N 126°17′31.87′′ E | ||
| 6 | Songji-myeon, Haenam-gun, Jeollanam-do | 34°23′35.54′′ N 126°28′23.45′′ E | ||
| 7 | P. seriata | Momunuidolgim | Anjwa-myeon, Sinan-gun, Jeollanam-do | 34°45′49.37′′ N 126°07′50.54′′ E |
| 8 | Songji-myeon, Haenam-gun, Jeollanam-do | 34°45′49.37′′ N 126°07′50.54′′ E | ||
| 9 | P. suborbiculata | Dunggeundolgim | Nam-myeon, Yeosu-si, Jeollanam-do | 34°25′32.73′′ N 127°47′31.33′′ E |
| 10 | P. pseudolinearis | Ginipdolgim | Ulleung-gun, Gyeongsangbuk-do | 37°27′31.55′′ N 130°54′14.98′′ E |
| 11 | P. haitanensis | Haitanensisgim | Dried laver product from China |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Choi, S.-J.; Choi, C. An Efficient PCR-RFLP Method for the Rapid Identification of Korean Pyropia Species. Molecules 2017, 22, 2182. https://doi.org/10.3390/molecules22122182
Kim Y, Choi S-J, Choi C. An Efficient PCR-RFLP Method for the Rapid Identification of Korean Pyropia Species. Molecules. 2017; 22(12):2182. https://doi.org/10.3390/molecules22122182
Chicago/Turabian StyleKim, Yonguk, Sung-Je Choi, and Chulyung Choi. 2017. "An Efficient PCR-RFLP Method for the Rapid Identification of Korean Pyropia Species" Molecules 22, no. 12: 2182. https://doi.org/10.3390/molecules22122182
APA StyleKim, Y., Choi, S. -J., & Choi, C. (2017). An Efficient PCR-RFLP Method for the Rapid Identification of Korean Pyropia Species. Molecules, 22(12), 2182. https://doi.org/10.3390/molecules22122182