
Protective Effects of Amarogentin against Carbon Tetrachloride-Induced Liver Fibrosis in Mice

Abstract
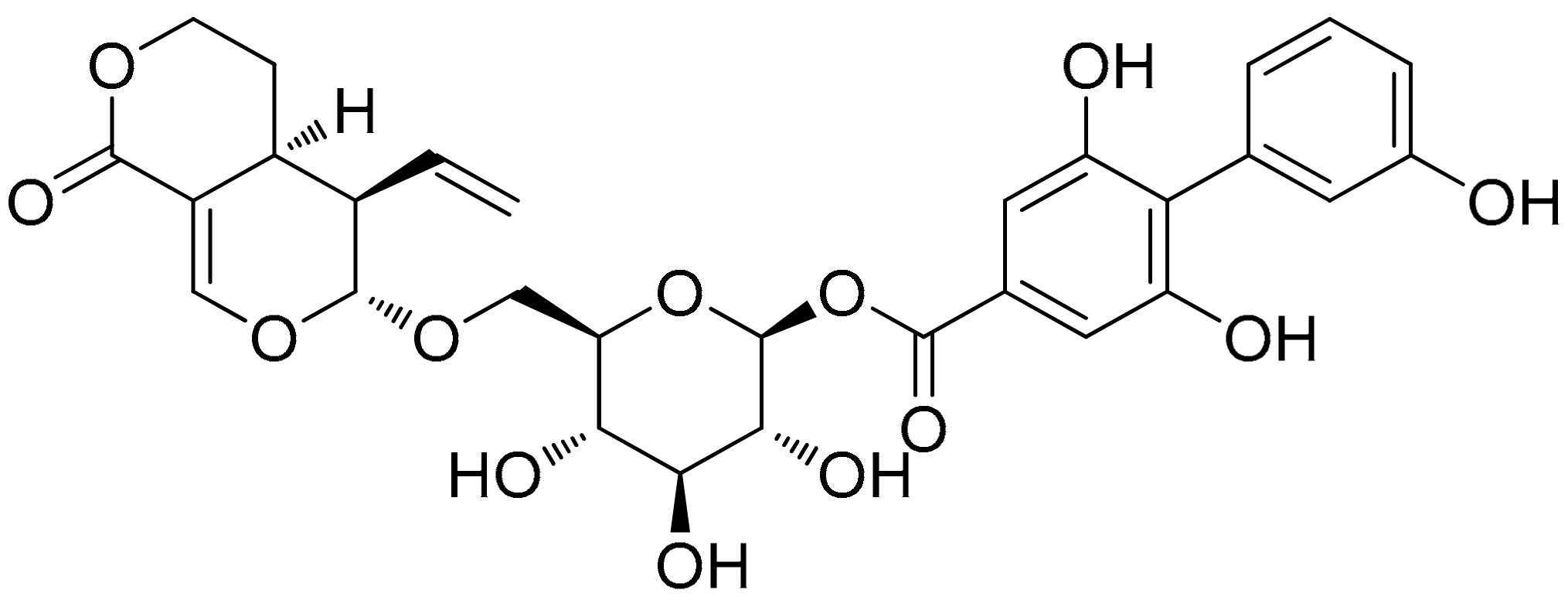
:1. Introduction
2. Results
2.1. Effect of AG on HSC Proliferation and Apoptosis
2.2. Effects of AG on the Relative Liver Weights and Liver Function in CCl4-Treated Mice
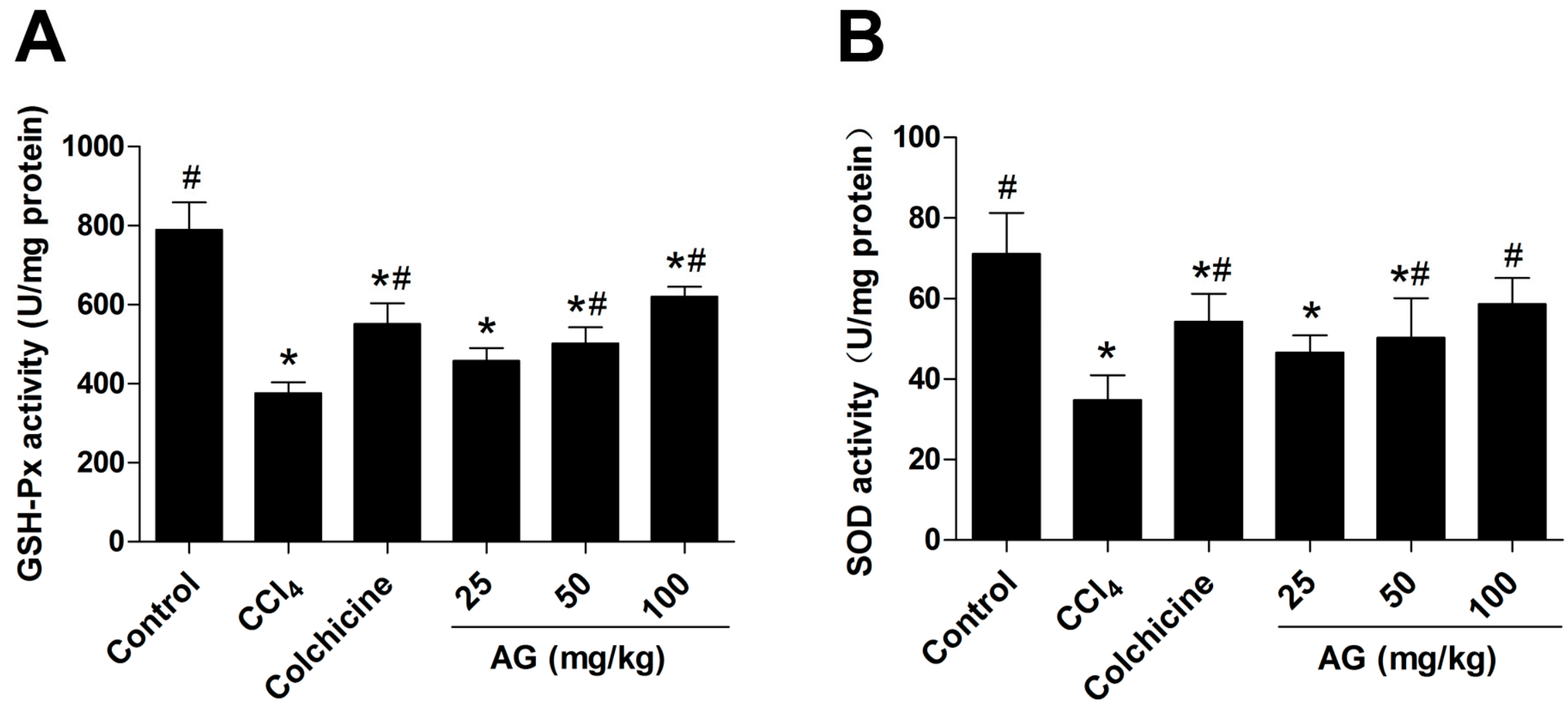
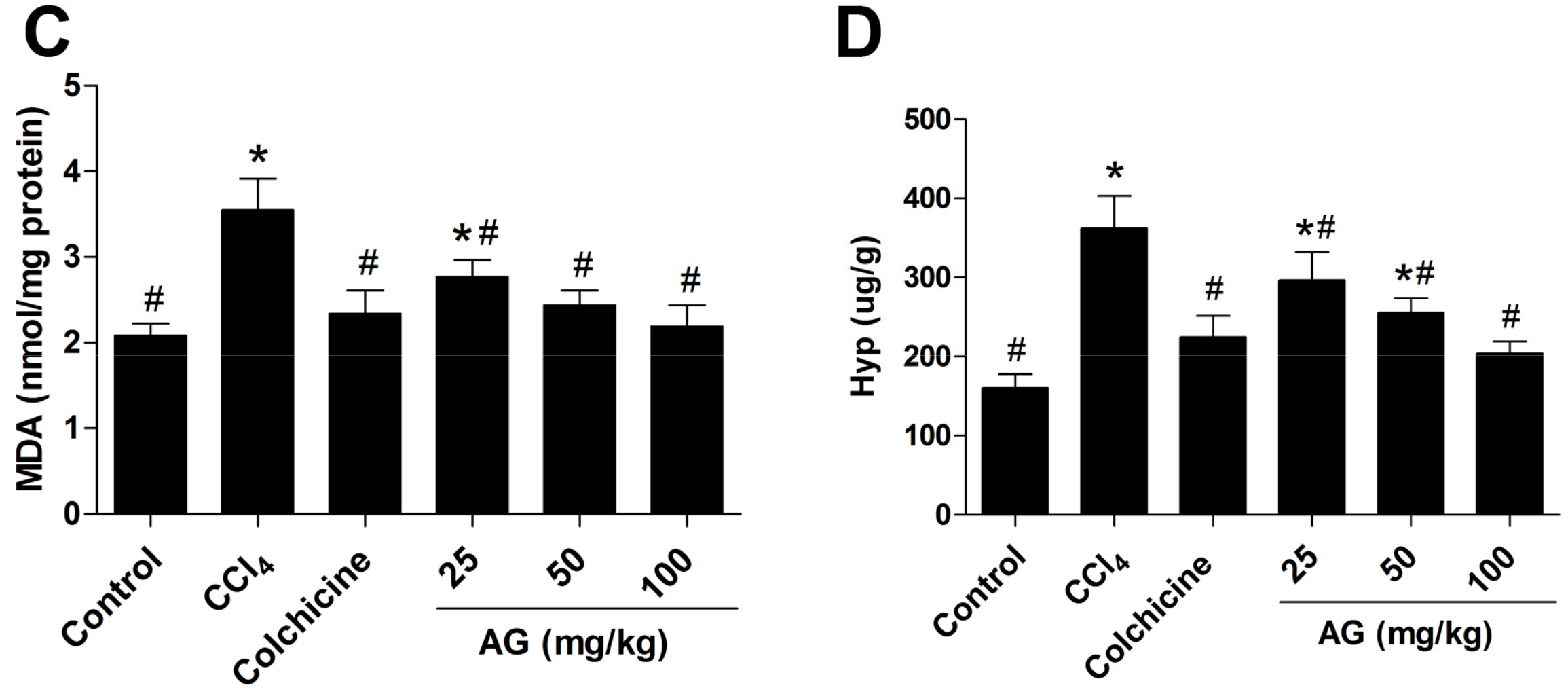
2.3. Effects of AG on Liver Lipid Peroxidation and Antioxidant Capacity
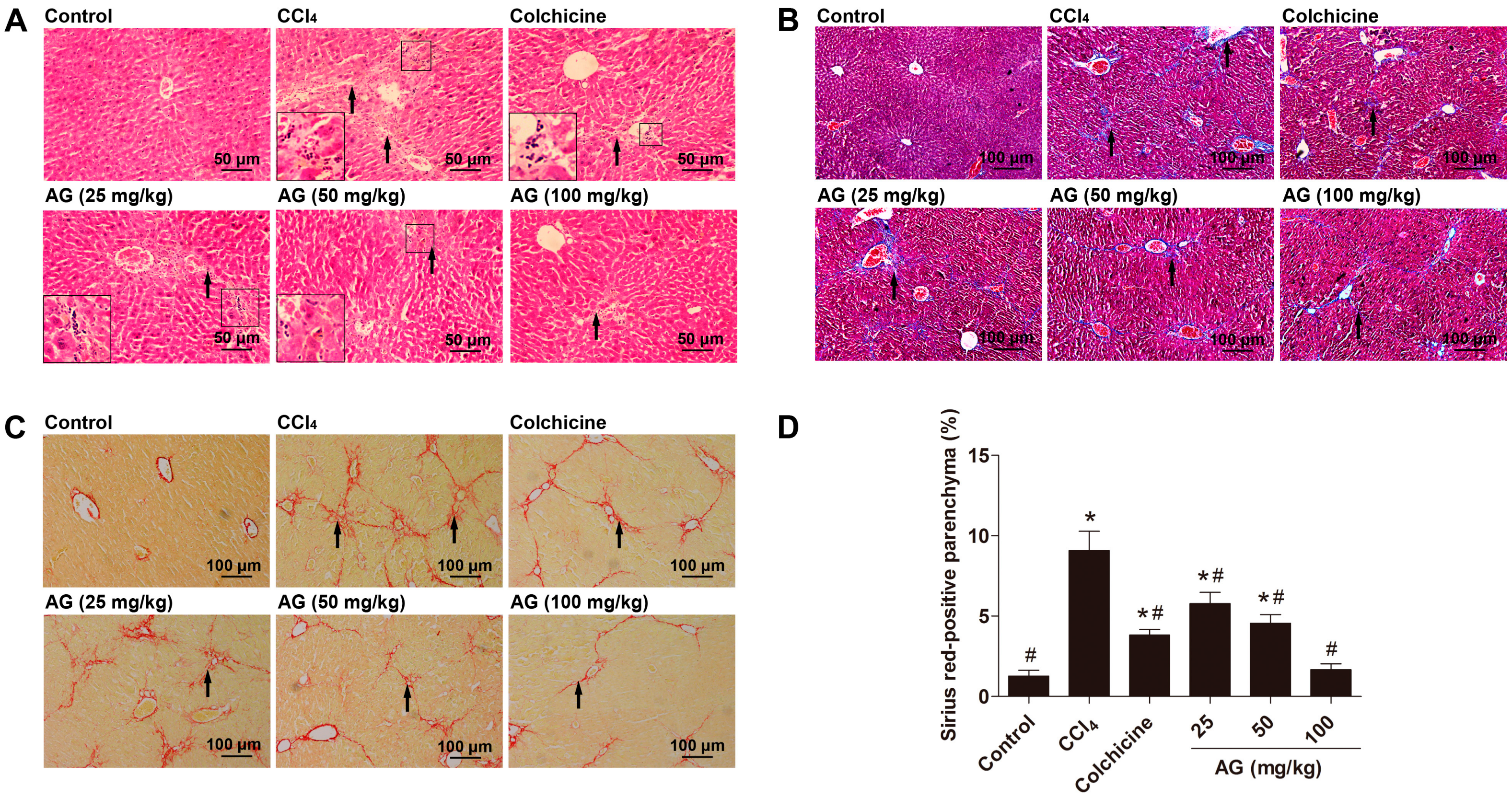
2.4. Effects of AG on Histological Changes
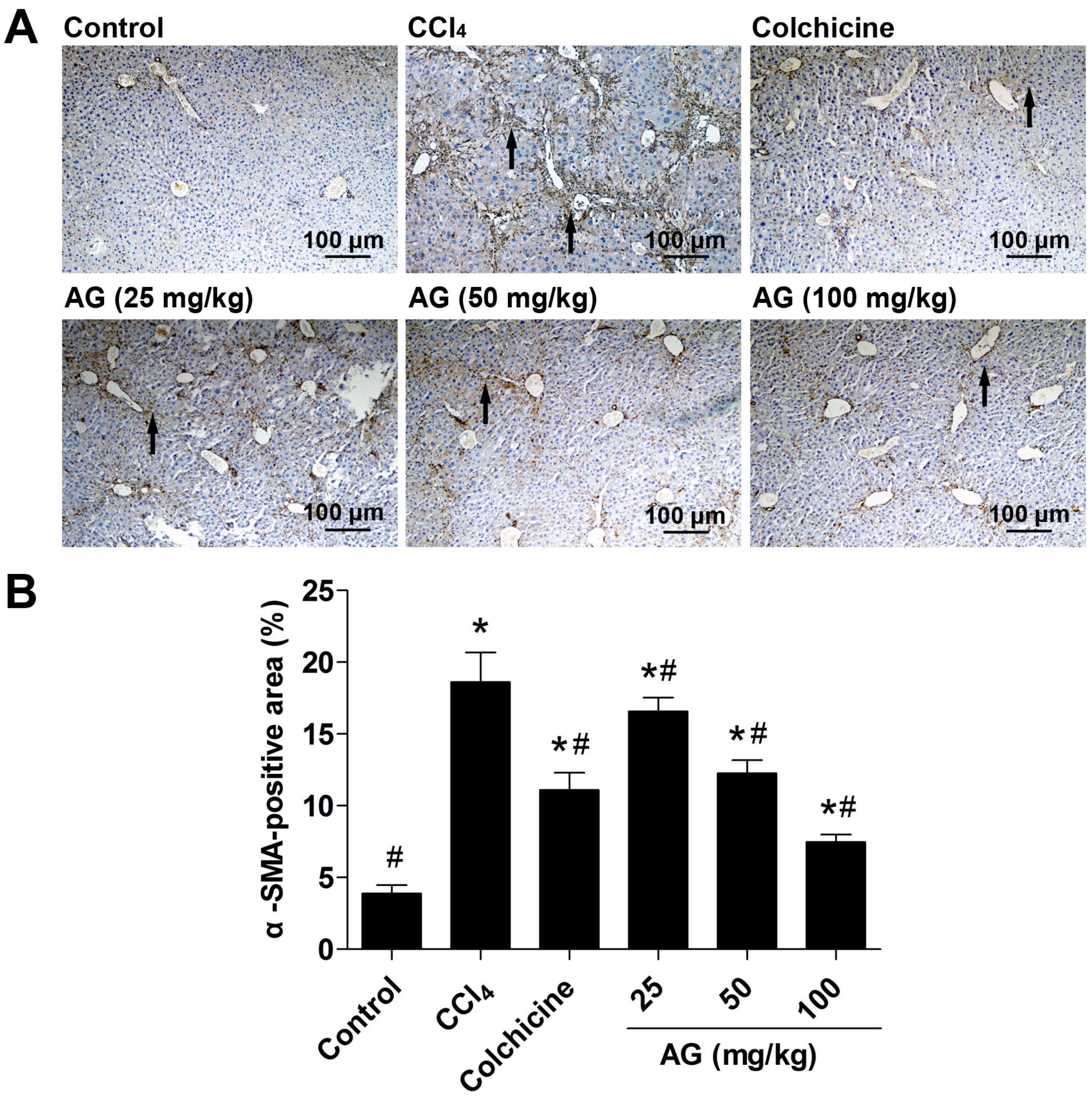
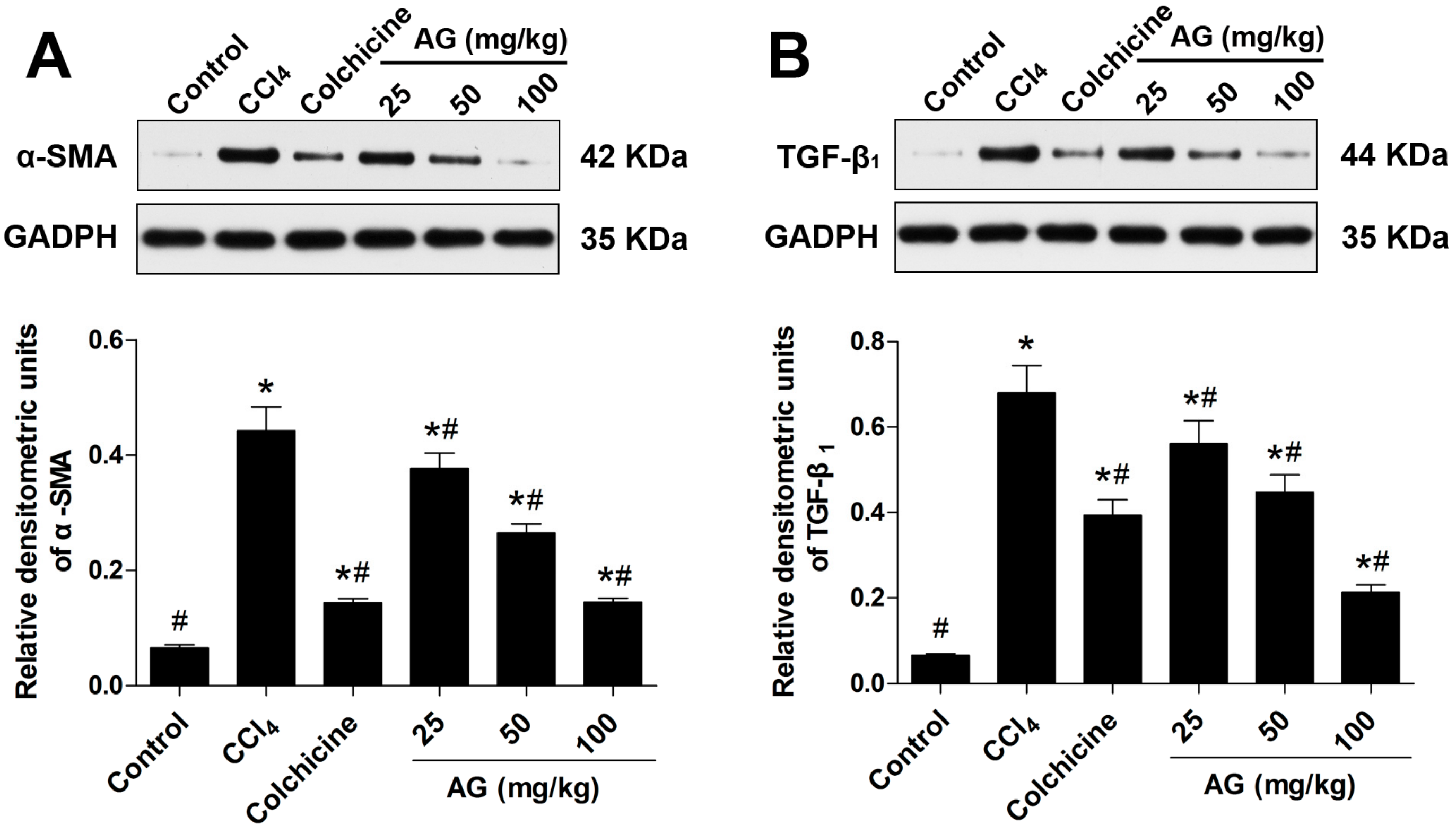
2.5. Effects of AG on the Expression of α-SMA and TGF-β1
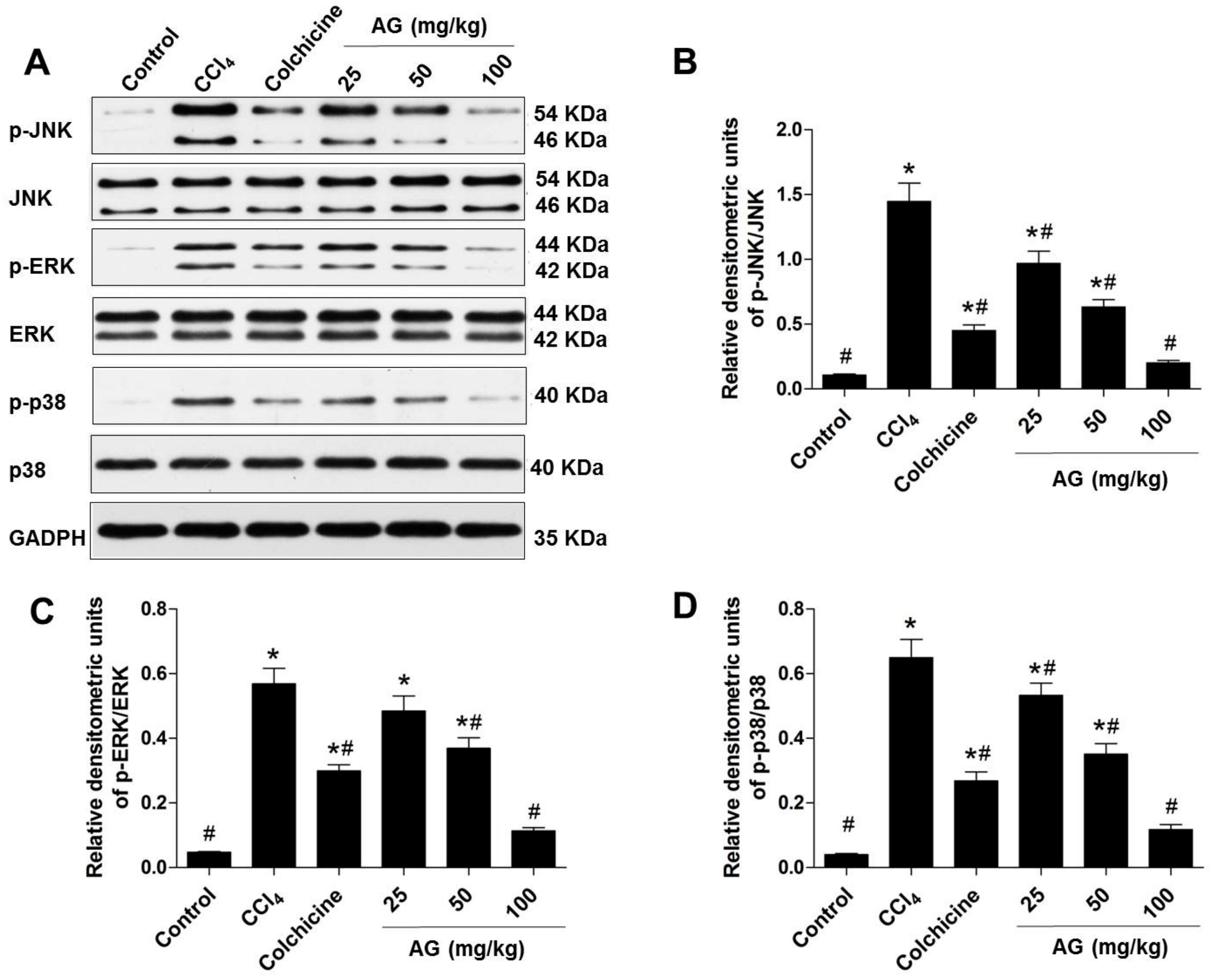
2.6. Effects of AG on MAPK Phosphorylation
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. HSC Culture and CCK-8 Assay
4.3. Animals and Treatment
4.4. Acute Toxicological Test
4.5. Biochemical Analysis
4.6. Histopathology Examinations
4.7. Immunohistochemical Staining for α-SMA
4.8. Western Blot Analysis
4.9. Statistical Analysis
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Friedman, S.L. Liver fibrosis—From bench to bedside. J. Hepatol. 2003, 38, 38–53. [Google Scholar] [CrossRef]
- Iwaisako, K.; Brenner, D.A.; Kisseleva, T. What’s new in liver fibrosis? The origin of myofibroblasts in liver fibrosis. J. Gastroenterol. Hepatol. 2012, 27 (Suppl. 2), 65–68. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Duan, F.; Peng, P.; Sun, L.; Ruan, Y.; Gu, J. Pyrroloquinoline-quinone suppresses liver fibrogenesis in mice. PLoS ONE 2015, 10, e0121939. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L. Cytokines and fibrogenesis. Semin. Liver Dis. 1999, 19, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Xiao, C.; Bandsma, R.H.; Naples, M.; Adeli, K.; Lewis, G.F. C-reactive protein impairs hepatic insulin sensitivity and insulin signaling in rats: Role of mitogen-activated protein kinases. Hepatology 2011, 53, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.W.D.; Boll, M.; Stamptl, A. Hepatotoxicity and mechanism of action of haloalkanes carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 32, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Shah, N.A. Hepatoprotective activity of Sonchus asper against carbon tetrachloride-induced injuries in male rats: A randomized controlled trial. BMC Complement. Altern. Med. 2012, 12, 90. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, G.; Shen, M.; Lin, K.; Lan, M.; Wu, L.; Chou, D.; Lin, C.; Su, C.; Sheu, J. Antioxidative and hepatoprotective effects of antrodia camphorata extract. J. Agric. Food Chem. 2003, 51, 3302–3308. [Google Scholar] [CrossRef] [PubMed]
- Barta, A.; Janega, P.; Babal, P.; Murar, E.; Cebova, M.; Pechanova, O. The effect of curcumin on liver fibrosis in the rat model of microsurgical cholestasis. Food Funct. 2015, 6, 2187–2193. [Google Scholar] [CrossRef] [PubMed]
- Weerawatanakorn, M.; Lee, Y.L.; Tsai, C.Y.; Lai, C.S.; Wan, X.; Ho, C.T.; Li, S.; Pan, M.H. Protective effect of theaflavin-enriched black tea extracts against dimethylnitrosamine-induced liver fibrosis in rats. Food Funct. 2015, 6, 1832–1840. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.B.; Sun, X.G.; Liu, Z.F.; Liu, Y.W.; Tang, J.; Liu, Q.; Ji, B.M.; Song, Y.H.; Zhou, Y.C.; Yang, M.H.; et al. Effects of dahuangzhechong pills on cytokines and mitogen activated protein kinase activation in rats with hepatic fibrosis. J. Ethnopharmacol. 2010, 132, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Luk, J.M.; Wang, X.; Liu, P.; Wong, K.F.; Chan, K.L.; Tong, Y.; Hui, C.K.; Lau, G.K.; Fan, S.T. Traditional chinese herbal medicines for treatment of liver fibrosis and cancer: From laboratory discovery to clinical evaluation. Liver Int. 2007, 27, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Inouye, H.; Nakamura, Y. Ṻber die monoterpenglucoside und verwandte naturstoffe—XIV: Die struktur der beiden stark bitter schmeckenden glucoside amarogentin und amaroswerin qus Swertia japonica. Tetrahedron 1971, 27, 1951–1966. [Google Scholar] [CrossRef]
- Wang, C.-Z.; Maier, U.H.; Eisenreich, W.; Adam, P.; Obersteiner, I.; Keil, M.; Bacher, A.; Zenk, M.H. Unexpected biosynthetic precursors of amarogentin- a retrobiosynthetic 13C NMR study. Eur. J.Org. Chem. 2001, 2001, 1459–1465. [Google Scholar] [CrossRef]
- Du, G.; Li, G. Sweertia mileensis in the treatment of acute virus hepatitis (an analytic report of 422 cases). Yunnan J. Tradit. Chin. Med. Mater. Med. 1981, 35–37. [Google Scholar]
- Karan, M.; Vasisht, K.; Handa, S.S. Antihepatotoxic activity of Swertia chirata on carbon tetrachloride induced hepatotoxicity in rats. Phytother. Res. 1999, 13, 24–30. [Google Scholar] [CrossRef]
- Pal, D.; Sur, S.; Mandal, S.; Das, A.; Roy, A.; Das, S.; Panda, C.K. Prevention of liver carcinogenesis by amarogentin through modulation of G1/S cell cycle check point and induction of apoptosis. Carcinogenesis 2012, 33, 2424–2431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, Y.; Qi, Y.; Han, X.; Yin, L.; Xu, L.; Liu, K.; Peng, J. Potent effects of dioscin against thioacetamide-induced liver fibrosis through attenuating oxidative stress in turn inhibiting inflammation, TGF-β/Smad and MAPK signaling pathways. J. Funct. Foods 2015, 16, 436–447. [Google Scholar] [CrossRef]
- Tsukamoto, H. Adipogenic phenotype of hepatic stellate cells. Alcohol. Clin. Exp. Res. 2005, 29, 132S–133S. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, C.; Luedde, T.; Sauerbruch, T.; Scholten, D.; Streetz, K.; Tacke, F.; Tolba, R.; Trautwein, C.; Trebicka, J.; Weiskirchen, R. Experimental liver fibrosis research update on animal models, legal issues and translational aspects. Fibrogenesis Tissue Repair 2013, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Floreani, A.; Lobello, S.; Brunetto, M.; Aneloni, V.; Chiaramonte, M. Colchicine in chronic hepatitis B: A pilot study. Aliment Pharmacol. Ther. 1998, 12, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Leveille, C.R.; Arias, I.M. Pathophysiology and pharmacologic modulation of hepatic fibrosis. J. Vet. Intern. Med. 1993, 7, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Nikolaidis, N.; Kountouras, J.; Giouleme, O.; Tzarou, V.; Chatzizisi, O.; Patsiaoura, K.; Papageorgiou, A.; Leontsini, M.; Eugenidis, N.; Zamboulis, C. Colchicine treatment of liver fibrosis. Hepatogastroenterology 2006, 53, 281–285. [Google Scholar] [PubMed]
- Chen, J.; Zhao, Y.; Tao, X.-y.; Zhang, M.; Sun, A.-D. Protective effect of blueberry anthocyanins in a CCl4-induced liver cell model. LWT Food Sci. Technol. 2015, 60, 1105–1112. [Google Scholar] [CrossRef]
- Abhilash, P.A.; Harikrishnan, R.; Indira, M. Ascorbic acid supplementation down-regulates the alcohol induced oxidative stress, hepatic stellate cell activation, cytotoxicity and mrna levels of selected fibrotic genes in guinea pigs. Free Radic. Res. 2012, 46, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Kawaratani, H.; Tsujimoto, T.; Kitazawa, T.; Yoshiji, H.; Uemura, M.; Fukui, H. Therapeutic effects of cytokine modulator Y-40138 in the rat alcoholic liver disease model. J. Gastroenterol. Hepatol. 2011, 26, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Fazaeli, M.; Yousefi, S.; Emam-Djomeh, Z. Investigation on the effects of microwave and conventional heating methods on the phytochemicals of pomegranate (Punica granatum L.) and black mulberry juices. Food Res. Int. 2013, 50, 568–573. [Google Scholar] [CrossRef]
- Singh, P.P.; Ambika; Chauhan, S.M. Activity-guided isolation of antioxidant xanthones from swertia chirayita (Roxb.) H. Karsten (Gentianaceae). Nat. Prod. Res. 2012, 26, 1682–1686. [Google Scholar] [CrossRef] [PubMed]
- Nastasijevic, B.; Lazarevic-Pasti, T.; Dimitrijevic-Brankovic, S.; Pasti, I.; Vujacic, A.; Joksic, G.; Vasic, V. Inhibition of myeloperoxidase and antioxidative activity of Gentiana lutea extracts. J. Pharm. Biomed. Anal. 2012, 66, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Moal, F.; Veal, N.; Vuillemin, E.; Barriere, E.; Wang, J.; Fizanne, L.; Oberti, F.; Douay, O.; Gallois, Y.; Bonnefont-Rousselot, D.; et al. Hemodynamic and antifibrotic effects of a selective liver nitric oxide donor V-PYRRO/NO in bile duct ligated rats. World J. Gastroenterol. 2006, 12, 6639–6645. [Google Scholar] [CrossRef] [PubMed]
- Medda, S.; Mukhopadhyay, S.; Basu, M.K. Evaluation of the in-vivo activity and toxicity of amarogentin, an antileishmanial agent, in both liposomal and niosomal forms. J. Antimicrob. Chemother. 1999, 44, 791–794. [Google Scholar] [CrossRef]
- Friedman, S.L. Hepatic stellate cells: Protean, multifunctional, and enigmatic cells of the liver. Physiol. Rev. 2008, 88, 125–172. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Gea, V.; Friedman, S.L. Pathogenesis of liver fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef] [PubMed]
- Song, S.Z.; Choi, Y.H.; Jin, G.Y.; Li, G.Z.; Yan, G.H. Protective effect of cornuside against carbon tetrachloride-induced acute hepatic injury. Biosci. Biotechnol. Biochem. 2011, 75, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Niu, M.; Zhai, X.; Zhou, Q.; Zhou, Y. β-catenin pathway is required for TGF-β1 inhibition of PPARγ expression in cultured hepatic stellate cells. Pharmacol. Res. 2012, 66, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Ni, M.M.; Xing, L.; Li, X.F.; Meng, X.M.; Huang, C.; Li, J. NLRC5 regulates TGF-β1-induced proliferation and activation of hepatic stellate cells during hepatic fibrosis. Int. J. Biochem. Cell Biol. 2016, 70, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Korsmeyer, S.J.; Yin, X.M.; Oltvai, Z.N.; Veis-Novack, D.J.; Linette, G.P. Reactive oxygen species and the regulation of cell death by the Bcl-2 gene family. Biochim. Biophys. Acta 1995, 1271, 63–66. [Google Scholar] [CrossRef]
- Reed, D.R.G.A. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar]
- Burt, A.D.C.L. Oakley lecture (1993). Cellular and molecular aspects of hepatic fibrosis. J. patHol. 1993, 170, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, W.; Baldassare, J.J.; Bora, P.S.; Chen, S.; Poulos, J.E.; O'Neill, R.; Britton, R.S.; Bacon, B.R. The ethanol metabolite, linolenic acid ethyl ester, stimulates mitogen-activated protein kinase and cyclin signaling in hepatic stellate cells. Life Sci. 2003, 73, 1083–1096. [Google Scholar] [CrossRef]
- Marra, F.; Arrighi, M.C.; Fazi, M.; Caligiuri, A.; Pinzani, M.; Romanelli, R.G.; Efsen, E.; Laffi, G.; Gentilini, P. Extracellular signal-regulated kinase activation differentially regulates platelet-derived growth factor’s actions in hepatic stellate cells, and is induced by in vivo liver injury in the rat. Hepatology 1999, 30, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Foo, N.-P.; Lin, S.-H.; Lee, Y.-H.; Wu, M.-J.; Wang, Y.-J. Α-lipoic acid inhibits liver fibrosis through the attenuation of ros-triggered signaling in hepatic stellate cells activated by PDGF and TGF-β. Toxicology 2011, 282, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Varela-Rey, M.; Montiel-Duarte, C.; Oses-Prieto, J.A.; Lopez-Zabalza, M.J.; Jaffrezou, J.P.; Rojkind, M.; Iraburu, M.J. p38 MAPK mediates the regulation of α1(I) procollagen mRNA levels by TGF-α and TGF-β in a cell line of rat hepatic stellate cells(I). FEBS Lett. 2002, 528, 133–138. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, J.C.; Zhang, D.; Zhang, J.A.; Ma, J.J.; Jiang, H.Q. New insights into the antifibrotic effects of sorafenib on hepatic stellate cells and liver fibrosis. J. Hepatol. 2010, 53, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.M.; Jiang, M.D.; Zeng, W.Z.; Xu, H.; Wang, Y.X.; Ma, H.D.; Xie, F.W.; Zhang, Y.; Qin, J.P.; Wu, X.L. Effects of extracellular signal-regulated kinase on rat cultured hepatic stellate cells stimulated by acetaldehyde. J. Dig. Dis. 2007, 8, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhang, Y.; Zhao, H.; Wang, S.W. Study on the pharmacokinetics of amarogentin in rats. Northwest Pharm. J. 2015, 30, 605–609. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Control | CCl4 | Colchicine | AG (25 mg/kg) | AG (50 mg/kg) | AG (100 mg/kg) |
|---|---|---|---|---|---|---|
| Initial body weight (g) | 19.26 ± 1.89 | 20.75 ± 1.33 | 19.75 ± 1.02 | 20.83 ± 1.34 | 20.88 ± 1.86 | 20.96 ± 1.54 |
| Final body weight (g) | 21.74 ± 1.68 | 22.45 ± 0.81 | 22.14 ± 1.16 | 23.19 ± 1.92 | 23.25 ± 1.20 | 23.38 ± 0.60 |
| Weight increase (g) | 2.48 ± 0.31 # | 1.70 ± 0.22 * | 2.39 ± 0.25 # | 2.36 ± 0.21 # | 2.37 ± 0.26 # | 2.42 ± 0.30 # |
| Liver weight (g) | 0.93 ± 0.12 | 1.15 ± 0.15 | 1.05 ± 0.09 | 1.12 ± 0.27 | 1.10 ± 0.11 | 1.05 ± 0.08 |
| Relative liver weight (%) | 4.26 ± 0.46 # | 5.13 ± 0.45 * | 4.74 ± 0.35 | 4.82 ± 0.57 | 4.73 ± 0.26 | 4.48 ± 0.36 # |
| ALT (U/L) | 19.75 ± 4.74 # | 58.53 ± 6.30 * | 26.02 ± 6.92 # | 38.29 ± 5.89 *# | 29.56 ± 6.56 # | 26.07 ± 4.60 # |
| AST (U/L) | 36.05 ± 8.24 # | 124.59 ± 6.48 * | 58.81 ± 8.81 *# | 89.82 ± 6.30 *# | 74.75 ± 8.26 *# | 55.86 ± 4.29 # |
| Alb (g/L) | 22.27 ± 3.92 # | 11.22 ± 1.65 * | 15.47 ± 2.69 * | 13.44 ± 1.22 * | 14.36 ± 2.77 * | 20.90 ± 4.01 # |
| cGMP (pmol/mg) | 1253.95 ± 206.13 # | 561.38 ± 92.87 * | 854.96 ± 142.36 *# | 689.69 ± 119.36 * | 842.81 ± 135.64 *# | 965.58 ± 180.36 *# |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhao, H.; Li, H.; Cao, W.; Wang, F.; Zhang, T.; Wang, S.-W. Protective Effects of Amarogentin against Carbon Tetrachloride-Induced Liver Fibrosis in Mice. Molecules 2017, 22, 754. https://doi.org/10.3390/molecules22050754
Zhang Y, Zhao H, Li H, Cao W, Wang F, Zhang T, Wang S-W. Protective Effects of Amarogentin against Carbon Tetrachloride-Induced Liver Fibrosis in Mice. Molecules. 2017; 22(5):754. https://doi.org/10.3390/molecules22050754
Chicago/Turabian StyleZhang, Ya, Hang Zhao, Hua Li, Wei Cao, Fang Wang, Tian Zhang, and Si-Wang Wang. 2017. "Protective Effects of Amarogentin against Carbon Tetrachloride-Induced Liver Fibrosis in Mice" Molecules 22, no. 5: 754. https://doi.org/10.3390/molecules22050754
APA StyleZhang, Y., Zhao, H., Li, H., Cao, W., Wang, F., Zhang, T., & Wang, S. -W. (2017). Protective Effects of Amarogentin against Carbon Tetrachloride-Induced Liver Fibrosis in Mice. Molecules, 22(5), 754. https://doi.org/10.3390/molecules22050754