
The Effects of Selected Sesquiterpenes from Myrica rubra Essential Oil on the Efficacy of Doxorubicin in Sensitive and Resistant Cancer Cell Lines

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
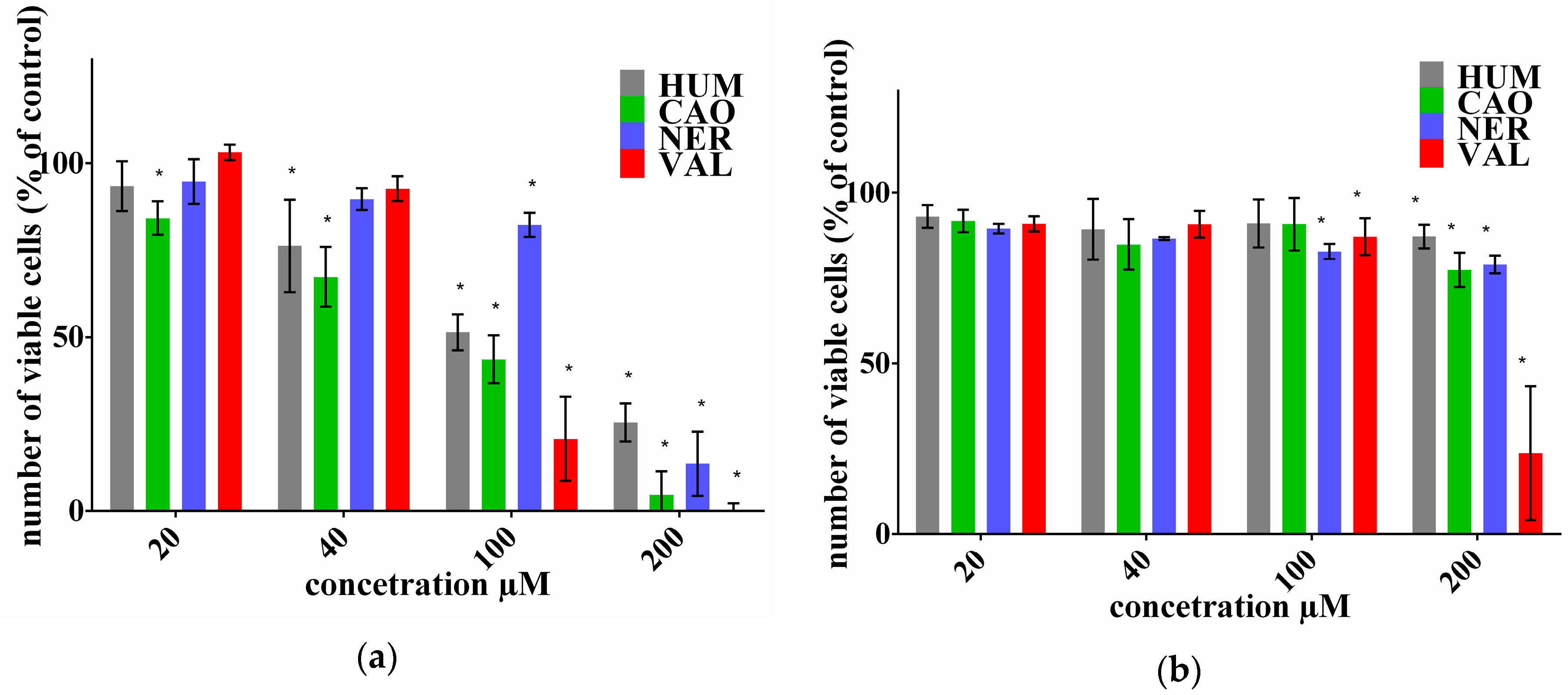
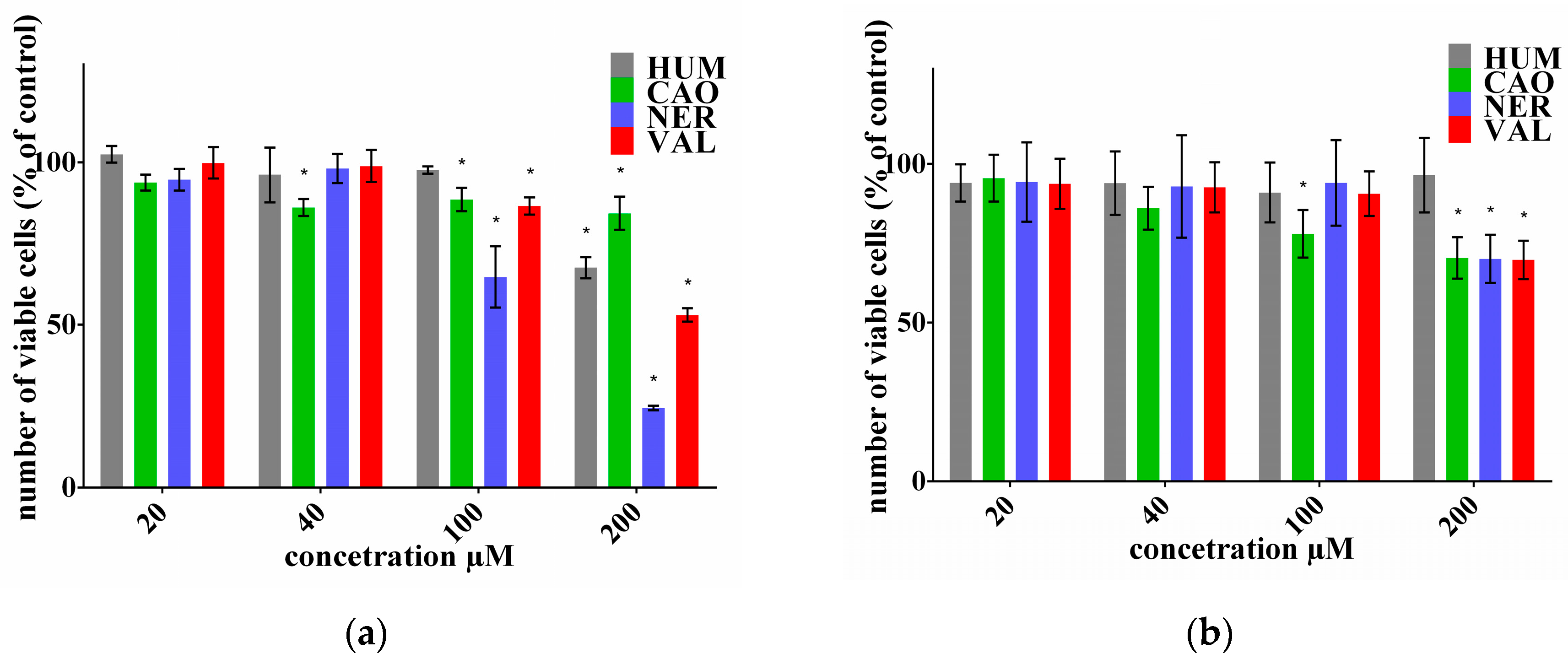
2.1. Effect of Sesquiterpenes on Cancer Cell Proliferation
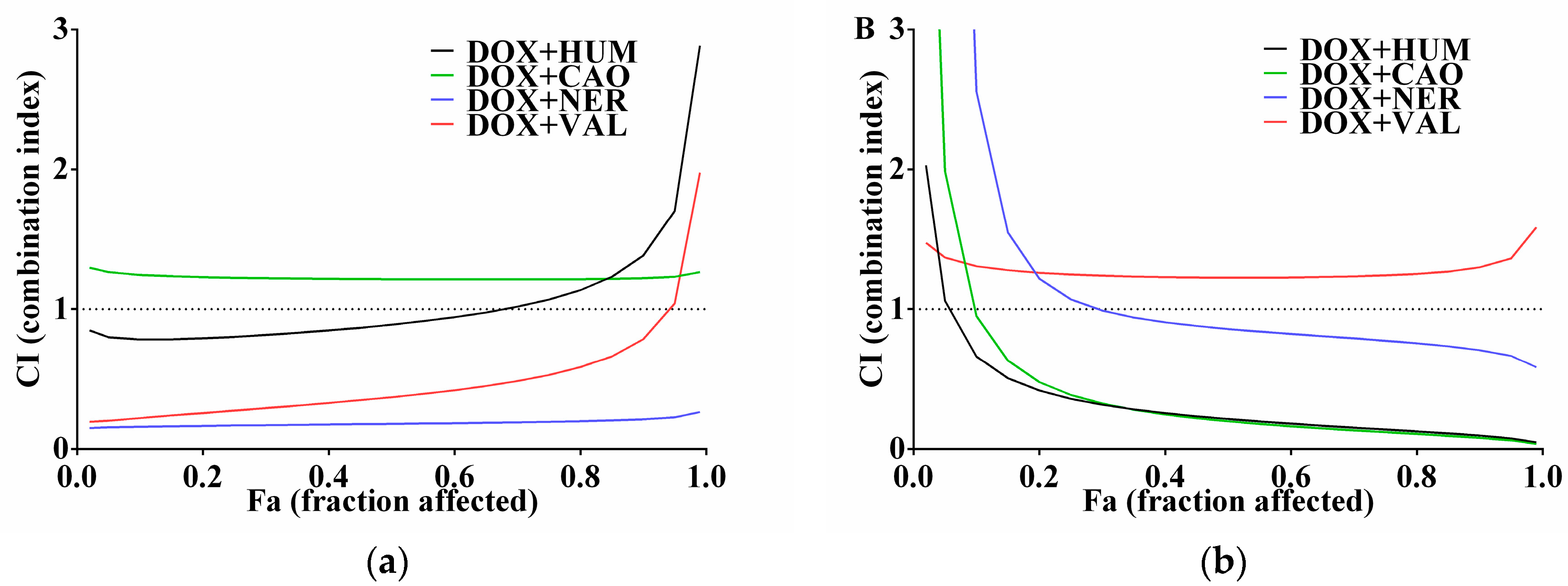
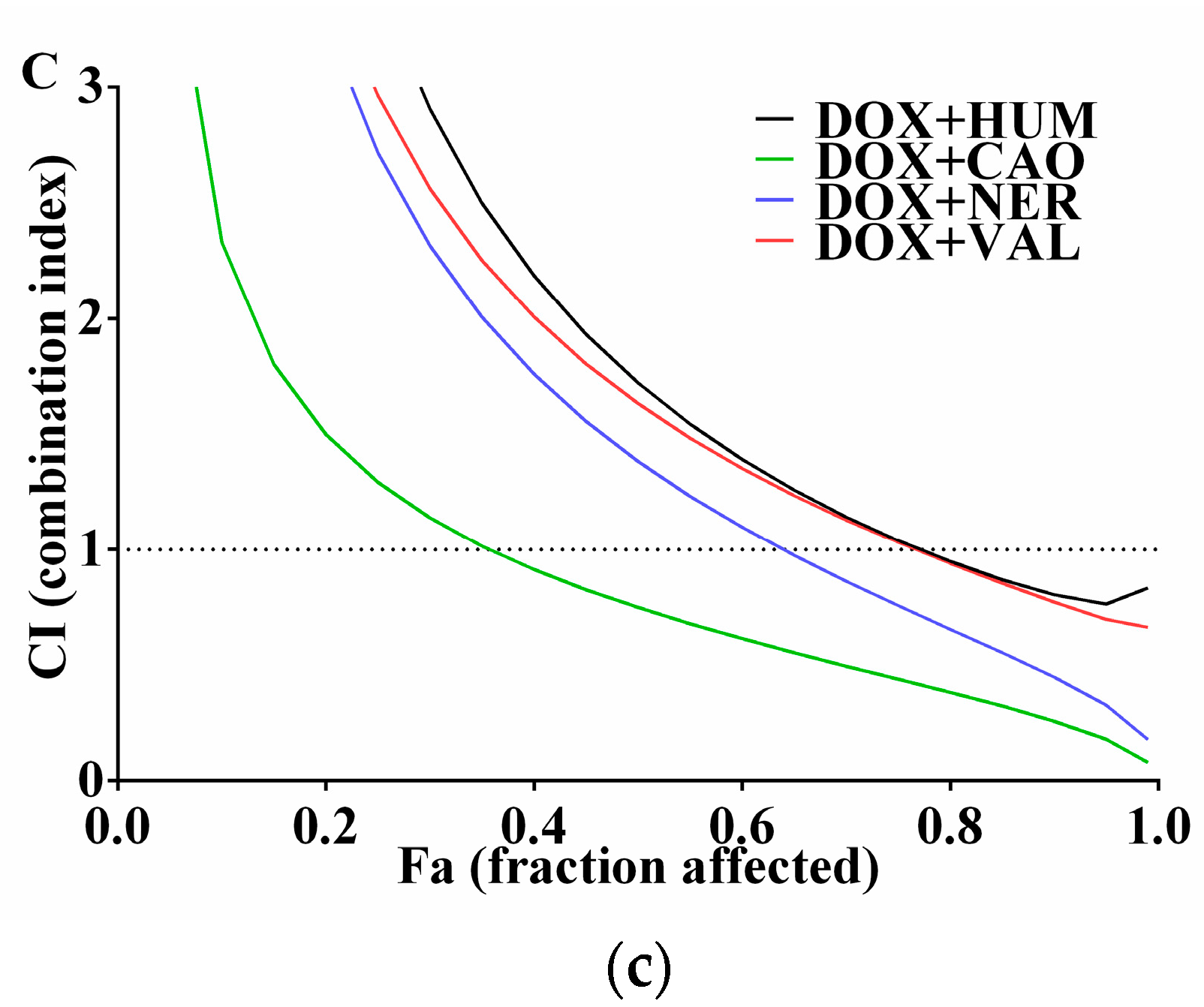
2.2. Effect of Sesquiterpenes in Combinations with DOX on Cancer Cell Proliferation
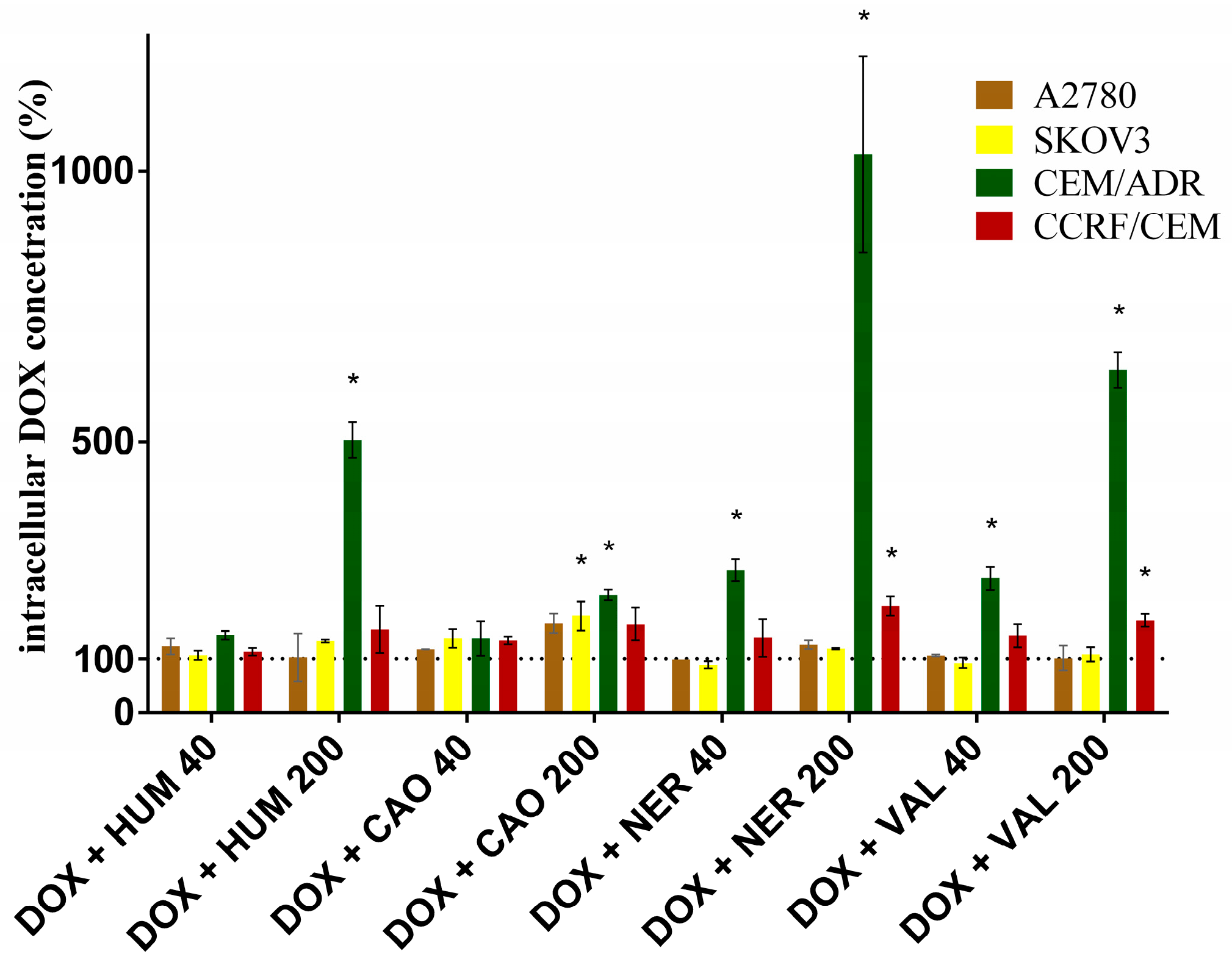
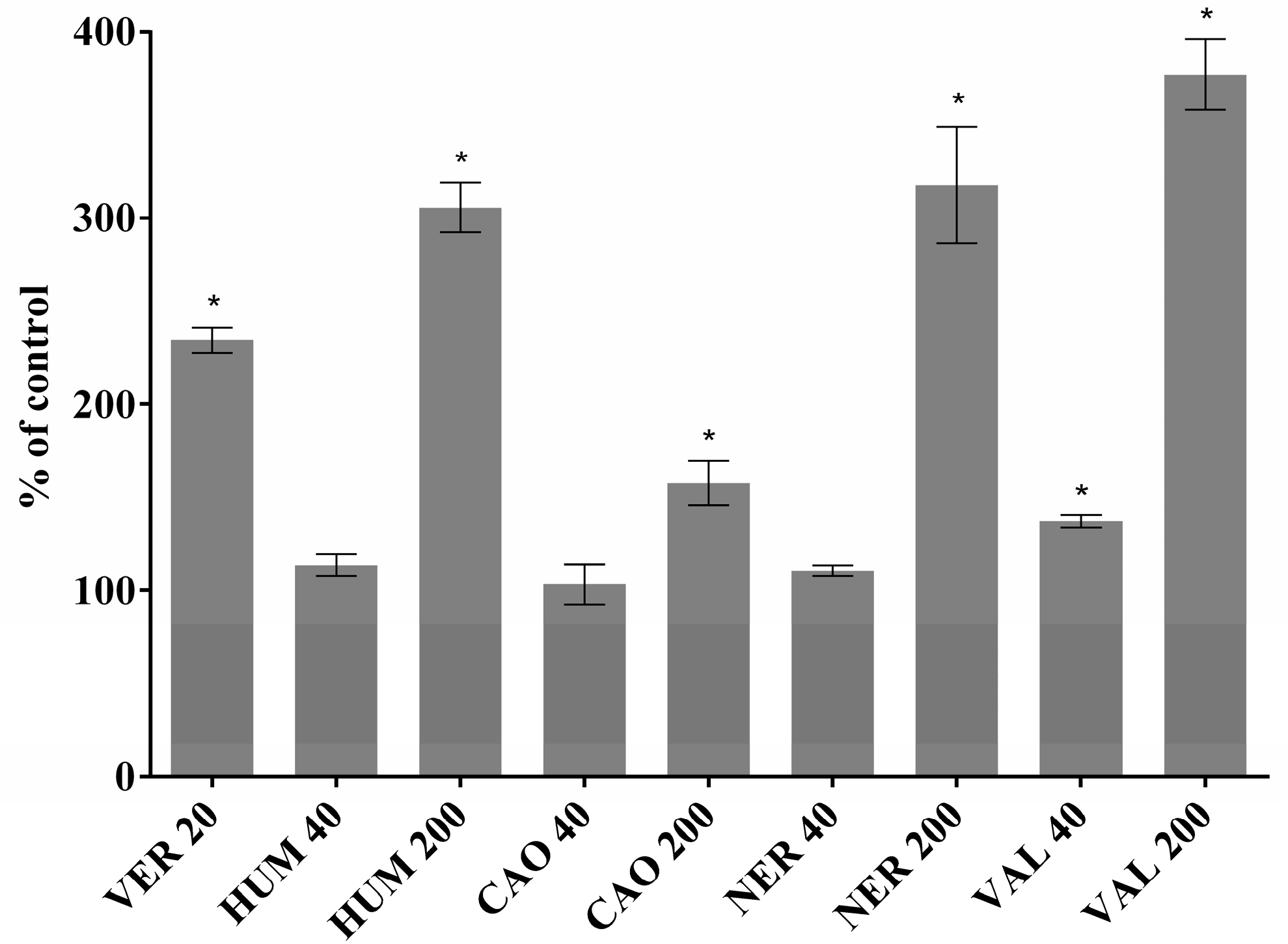
2.3. Effect of Sesquiterpenes on Accumulation of DOX and Rhodamine 123 in Cancer Cells
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Cancer Cell Culture
3.3. Tests of Cell Viability
3.4. The Neutral Red Uptake (NRU) Test
3.5. Alamar Blue (AB)
3.6. Accumulation of DOX in Cells
3.7. Accumulation of RHO123
3.8. Western Blot Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Konig, G.M.; Wright, A.D. New and unusual sesquiterpenes: Kelsoene, prespatane, epi-gamma-gurjunene, and t-cadinthiol, from the tropical marine sponge cymbastela hooperi. J. Org. Chem. 1997, 62, 3837–3840. [Google Scholar] [CrossRef]
- Ayer, W.A.; Browne, L.M. Terpenoid metabolites of mushrooms and related basidiomycetes. Tetrahedron 1981, 37, 2199–2248. [Google Scholar] [CrossRef]
- Finefield, J.M.; Sherman, D.H.; Kreitman, M.; Williams, R.M. Enantiomeric natural products: Occurrence and biogenesis. Angew. Chem. 2012, 51, 4802–4836. [Google Scholar] [CrossRef] [PubMed]
- Bartikova, H.; Hanusova, V.; Skalova, L.; Ambroz, M.; Bousova, I. Antioxidant, pro-oxidant and other biological activities of sesquiterpenes. Curr. Top. Med. Chem. 2014, 14, 2478–2494. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zhao, Y.P.; Lou, C.H.; Zhao, H.J. Eupalinolide O, a novel sesquiterpene lactone from Eupatorium lindleyanum DC., induces cell cycle arrest and apoptosis in human MDA-MB-468 breast cancer cells. Oncol. Rep. 2016, 36, 2807–2813. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cui, L.; Feng, L.; Zhang, Z.H.; Song, J.; Liu, D.; Jia, X.B. Isoalantolactone inhibits the migration and invasion of human breast cancer MDA-MB-231 cells via suppression of the p38 MAPK/NF-kB signaling pathway. Oncol. Rep. 2016, 36, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Li, M.M.; Wang, G.C.; Shao, F.Y.; Chen, W.B.; Xia, C.; Wang, S.; Li, Y.L.; Zhou, G.X.; Liu, Z. EM23, a natural sesquiterpene lactone from elephantopus mollis, induces apoptosis in human myeloid leukemia cells through thioredoxin- and reactive oxygen species-mediated signaling pathways. Front Pharmacol. 2016, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Van Haaften, C.; Boot, A.; Corver, W.E.; van Eendenburg, J.D.H.; Trimbos, B.J.M.Z.; van Wezel, T. Synergistic effects of the sesquiterpene lactone, EPD, with cisplatin and paclitaxel in ovarian cancer cells. J. Exp. Clin. Cancer Res. 2015, 34, 38. [Google Scholar] [CrossRef] [PubMed]
- Quinn, B.A.; Dash, R.; Sarkar, S.; Azab, B.; Bhoopathi, P.; Das, S.K.; Emdad, L.; Wei, J.; Pellecchia, M.; Sarkar, D.; et al. Pancreatic cancer combination therapy using a BH3 mimetic and a synthetic tetracycline. Cancer Res. 2015, 75, 2305–2315. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Lin, Z.H.; Zhang, B.; Guo, L.; Liu, S.; Li, H.; Zhang, J.B.; Ye, Q.H. Beta-elemene sensitizes hepatocellular carcinoma cells to oxaliplatin by preventing oxaliplatin-induced degradation of copper transporter 1. Sci. Rep. 2016, 6, 21010. [Google Scholar] [CrossRef] [PubMed]
- Ambroz, M.; Bousova, I.; Skarka, A.; Hanusova, V.; Kralova, V.; Matouskova, P.; Szotakova, B.; Skalova, L. The influence of sesquiterpenes from Myrica rubra on the antiproliferative and pro-oxidative effects of doxorubicin and its accumulation in cancer cells. Molecules 2015, 20, 15343–15358. [Google Scholar] [CrossRef] [PubMed]
- Pillozzi, S.; Masselli, M.; de Lorenzo, E.; Accordi, B.; Cilia, E.; Crociani, O.; Amedei, A.; Veltroni, M.; D’Amico, M.; Basso, G.; et al. Chemotherapy resistance in acute lymphoblastic leukemia requires herg1 channels and is overcome by herg1 blockers. Blood 2011, 117, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Mullighan, C.G.; Su, X.; Zhang, J.; Radtke, I.; Phillips, L.A.; Miller, C.B.; Ma, J.; Liu, W.; Cheng, C.; Schulman, B.A.; et al. Deletion of IKZF1 and prognosis in acute lymphoblastic leukemia. N. Engl. J. Med. 2009, 360, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [PubMed]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharm. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.R.; Jamieson, D.; Thomas, H.D.; Brown, C.D.; Boddy, A.V.; Veal, G.J. Characterisation of the roles of ABCB1, ABCC1, ABCC2 and ABCG2 in the transport and pharmacokinetics of actinomycin D in vitro and in vivo. Biochem. Pharmacol. 2013, 85, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Langhansova, L.; Hanusova, V.; Rezek, J.; Stohanslova, B.; Ambroz, M.; Kralova, V.; Vanek, T.; Lou, J.D.; Yun, Z.L.; Yang, J.; et al. Essential oil from Myrica rubra leaves inhibits cancer cell proliferation and induces apoptosis in several human intestinal lines. Ind. Crop. Prod. 2014, 59, 20–26. [Google Scholar] [CrossRef]
- Ambroz, M.; Hanusova, V.; Skarka, A.; Bousova, I.; Kralova, V.; Langhasova, L.; Skalova, L. Essential oil from Myrica rubra leaves potentiated antiproliferative and prooxidative effect of doxorubicin and its accumulation in intestinal cancer cells. Planta Med. 2016, 82, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Barla, A.; Topcu, G.; Oksuz, S.; Tumen, G.; Kingston, D.G.I. Identification of cytotoxic sesquiterpenes from Laurus nobilis L. Food Chem. 2007, 104, 1478–1484. [Google Scholar] [CrossRef]
- Saab, A.M.; Guerrini, A.; Sacchetti, G.; Maietti, S.; Zeino, M.; Arend, J.; Gambari, R.; Bernardi, F.; Efferth, T. Phytochemical analysis and cytotoxicity towards multidrug-resistant leukemia cells of essential oils derived from lebanese medicinal plants. Planta Med. 2012, 78, 1927–1931. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Drug combination studies and their synergy quantification using the chou-talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, H.; Yonehara, S.; Bonavida, B. Overcoming tumor necrosis factor and drug resistance of human tumor cell lines by combination treatment with anti-fas antibody and drugs or toxins. Cancer Res. 1993, 53, 2591–2596. [Google Scholar] [PubMed]
- Kartner, N.; Riordan, J.R.; Ling, V. Cell surface p-glycoprotein associated with multidrug resistance in mammalian cell lines. Science 1983, 221, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, H.; Safrit, J.T.; Bonavida, B. Synergistic effect of tumor necrosis factor-alpha- and diphtheria toxin-mediated cytotoxicity in sensitive and resistant human ovarian tumor cell lines. J. Immunol. 1991, 147, 2609–2616. [Google Scholar] [PubMed]
- Silva, R.; Vilas-Boas, V.; Carmo, H.; Dinis-Oliveira, R.J.; Carvalho, F.; Bastos, M.D.; Remiao, F. Modulation of p-glycoprotein efflux pump: Induction and activation as a therapeutic strategy. Pharmacol. Ther. 2015, 149, 1–123. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yang, Z.P.; Nie, Y.Z.; Shi, Y.Q.; Fan, D.M. Multi-drug resistance in cancer chemotherapeutics: Mechanisms and lab approaches. Cancer Lett. 2014, 347, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Al-Abd, A.M.; El-Dine, R.S.; El-Halawany, A.M. P-glycoprotein inhibitors of natural origin as potential tumor chemo-sensitizers: A review. J. Adv. Res. 2015, 6, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Skarka, A.; Skarydova, L.; Stambergova, H.; Wsol, V. Anthracyclines and their metabolism in human liver microsomes and the participation of the new microsomal carbonyl reductase. Chem. Biol. Interact. 2011, 191, 66–74. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the Sigma-Aldrich. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A2780 | SKOV3 | CCRF/CEM | CEM/ADR | |
|---|---|---|---|---|
| HUM | 78 | − | 224 | 303 |
| CAO | 43 | − | − | − |
| NER | 119 | − | 120 | 232 |
| VAL | 61 | 114 | 203 | 302 |
| DOX | 0.16 | 1.41 | 0.037 | − |
| A2780 | SKOV3 | CCRF/CEM | |
|---|---|---|---|
| HUM + DOX | 1.1 | 0.13 | 0.95 |
| CAO + DOX | 1.2 | 0.11 | 0.38 |
| NER + DOX | 0.19 | 0.76 | 0.65 |
| VAL + DOX | 0.58 | 1.25 | 0.94 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambrož, M.; Matoušková, P.; Skarka, A.; Zajdlová, M.; Žáková, K.; Skálová, L. The Effects of Selected Sesquiterpenes from Myrica rubra Essential Oil on the Efficacy of Doxorubicin in Sensitive and Resistant Cancer Cell Lines. Molecules 2017, 22, 1021. https://doi.org/10.3390/molecules22061021
Ambrož M, Matoušková P, Skarka A, Zajdlová M, Žáková K, Skálová L. The Effects of Selected Sesquiterpenes from Myrica rubra Essential Oil on the Efficacy of Doxorubicin in Sensitive and Resistant Cancer Cell Lines. Molecules. 2017; 22(6):1021. https://doi.org/10.3390/molecules22061021
Chicago/Turabian StyleAmbrož, Martin, Petra Matoušková, Adam Skarka, Martina Zajdlová, Kateřina Žáková, and Lenka Skálová. 2017. "The Effects of Selected Sesquiterpenes from Myrica rubra Essential Oil on the Efficacy of Doxorubicin in Sensitive and Resistant Cancer Cell Lines" Molecules 22, no. 6: 1021. https://doi.org/10.3390/molecules22061021
APA StyleAmbrož, M., Matoušková, P., Skarka, A., Zajdlová, M., Žáková, K., & Skálová, L. (2017). The Effects of Selected Sesquiterpenes from Myrica rubra Essential Oil on the Efficacy of Doxorubicin in Sensitive and Resistant Cancer Cell Lines. Molecules, 22(6), 1021. https://doi.org/10.3390/molecules22061021