
Endophytic Bacteria Isolated from Panax ginseng Improves Ginsenoside Accumulation in Adventitious Ginseng Root Culture


Abstract
:1. Introduction
2. Results and Discussion
2.1. Bacterial Endophytes as Elicitors
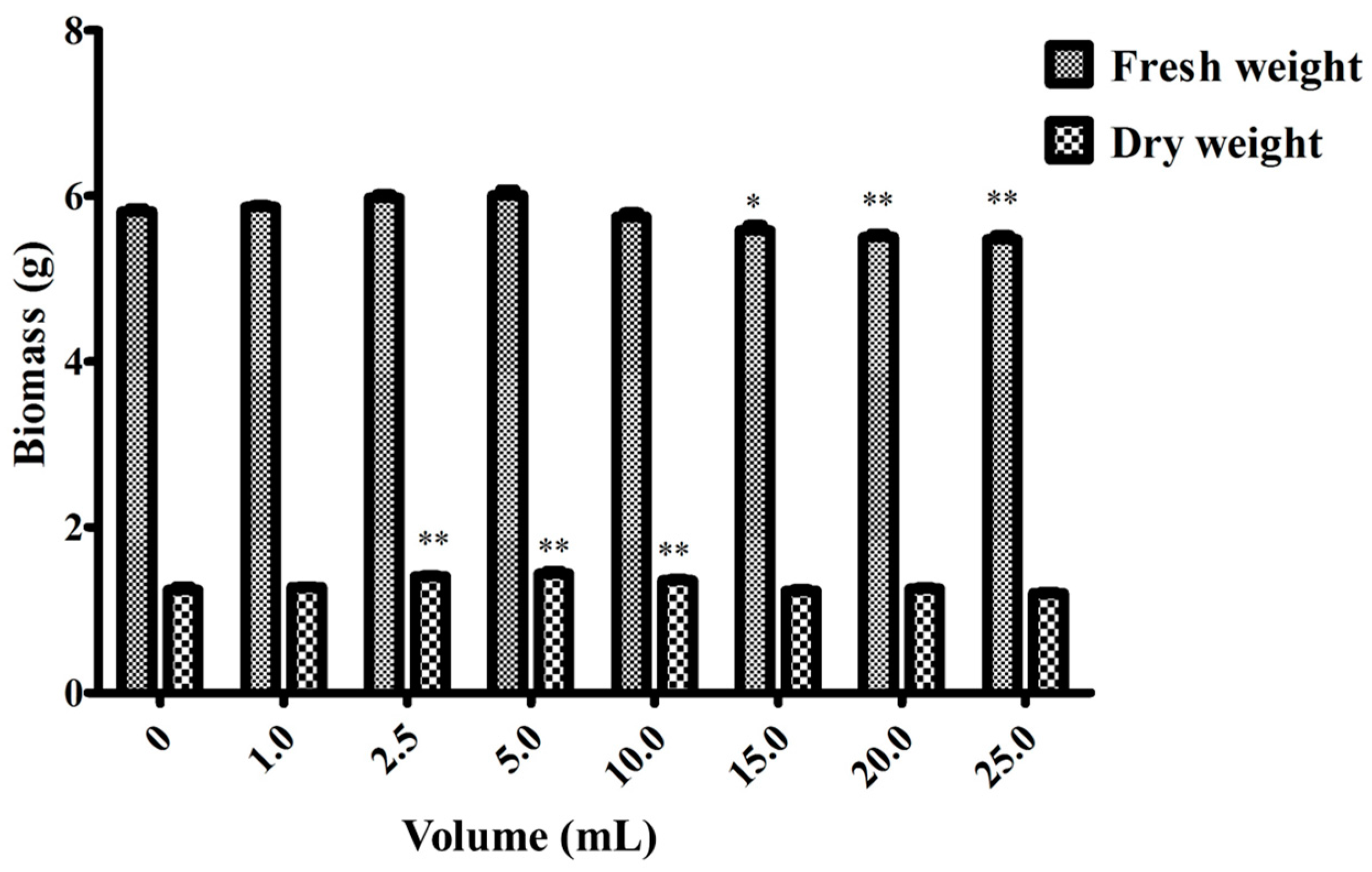
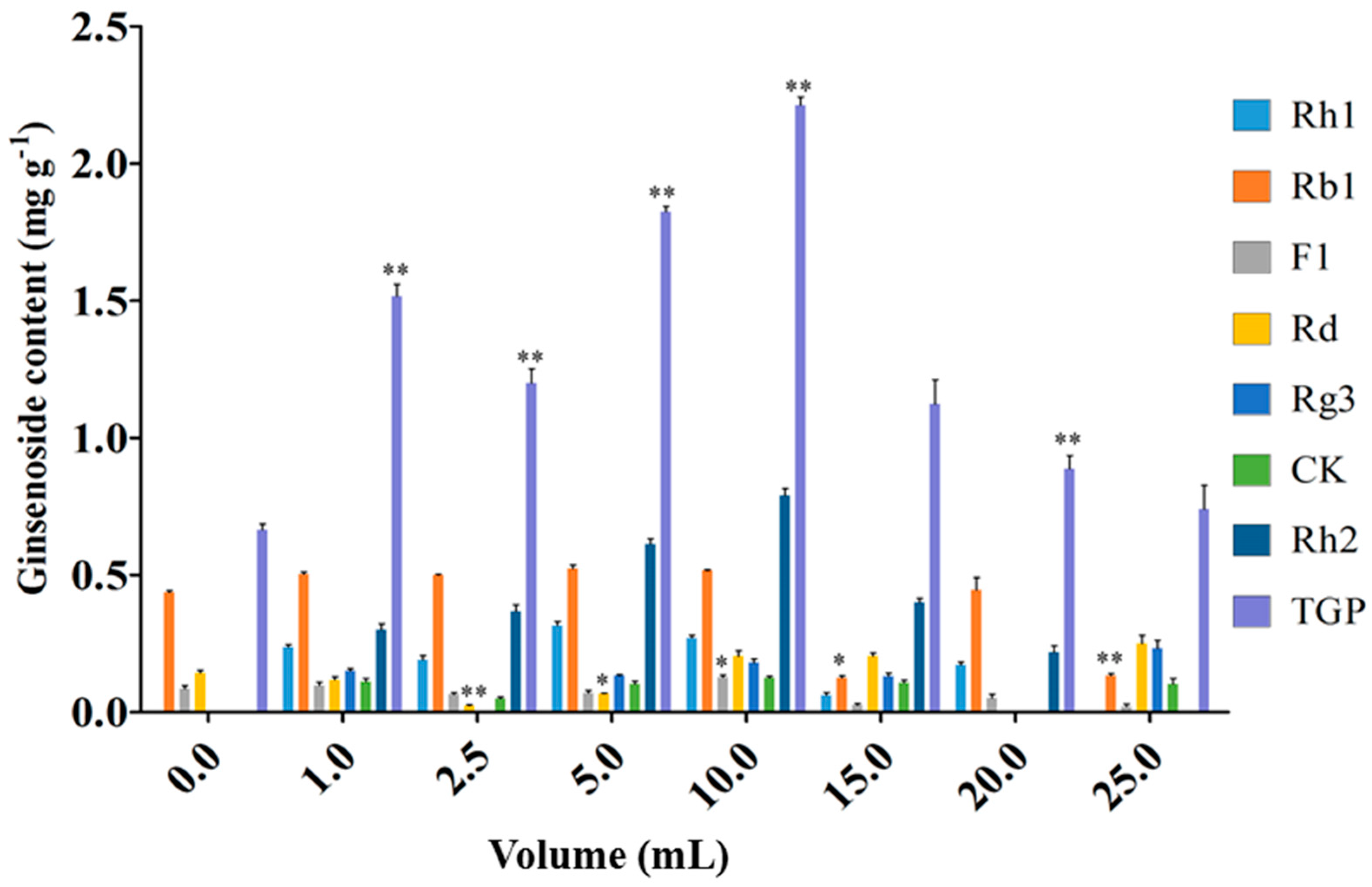
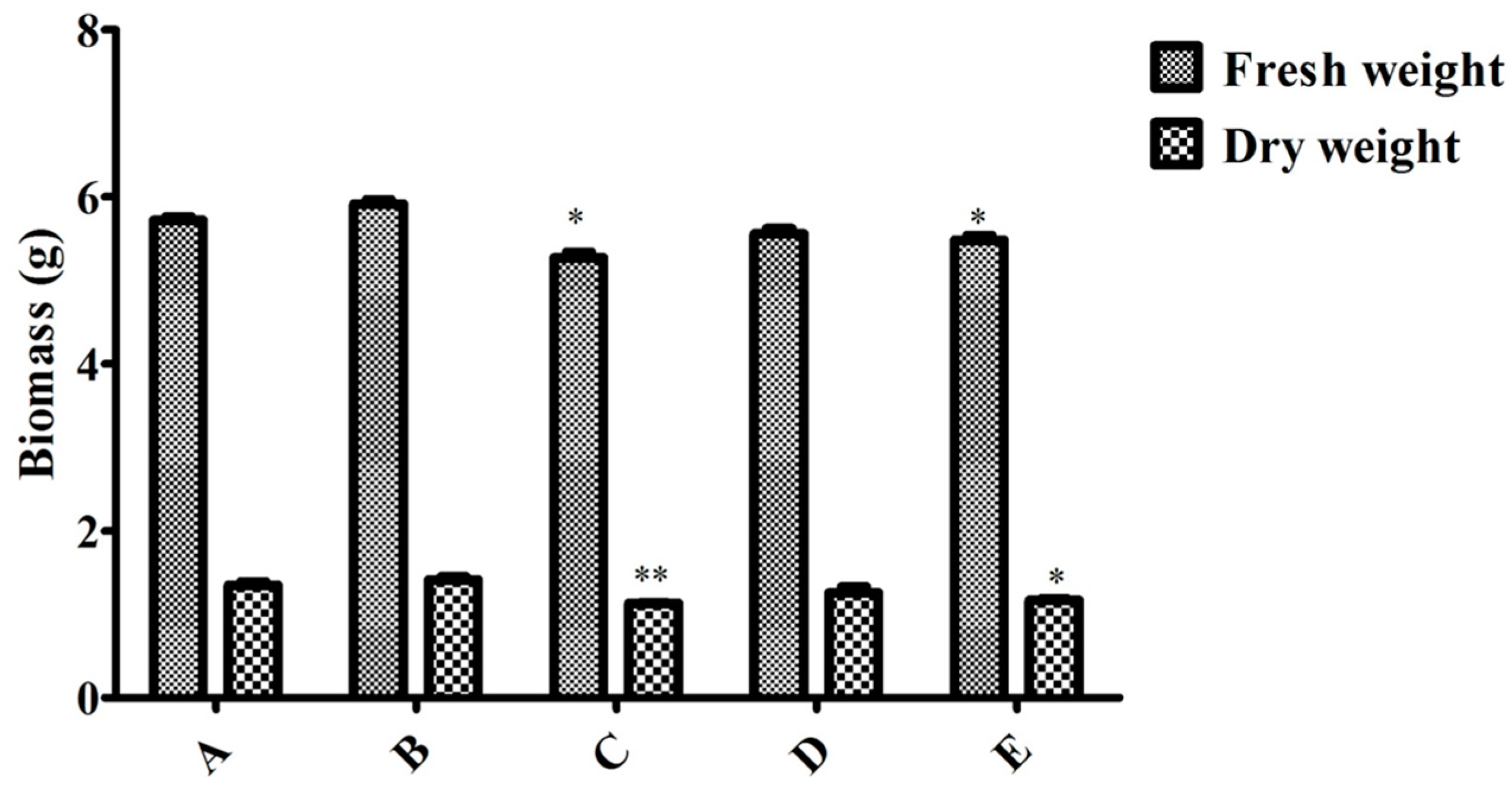
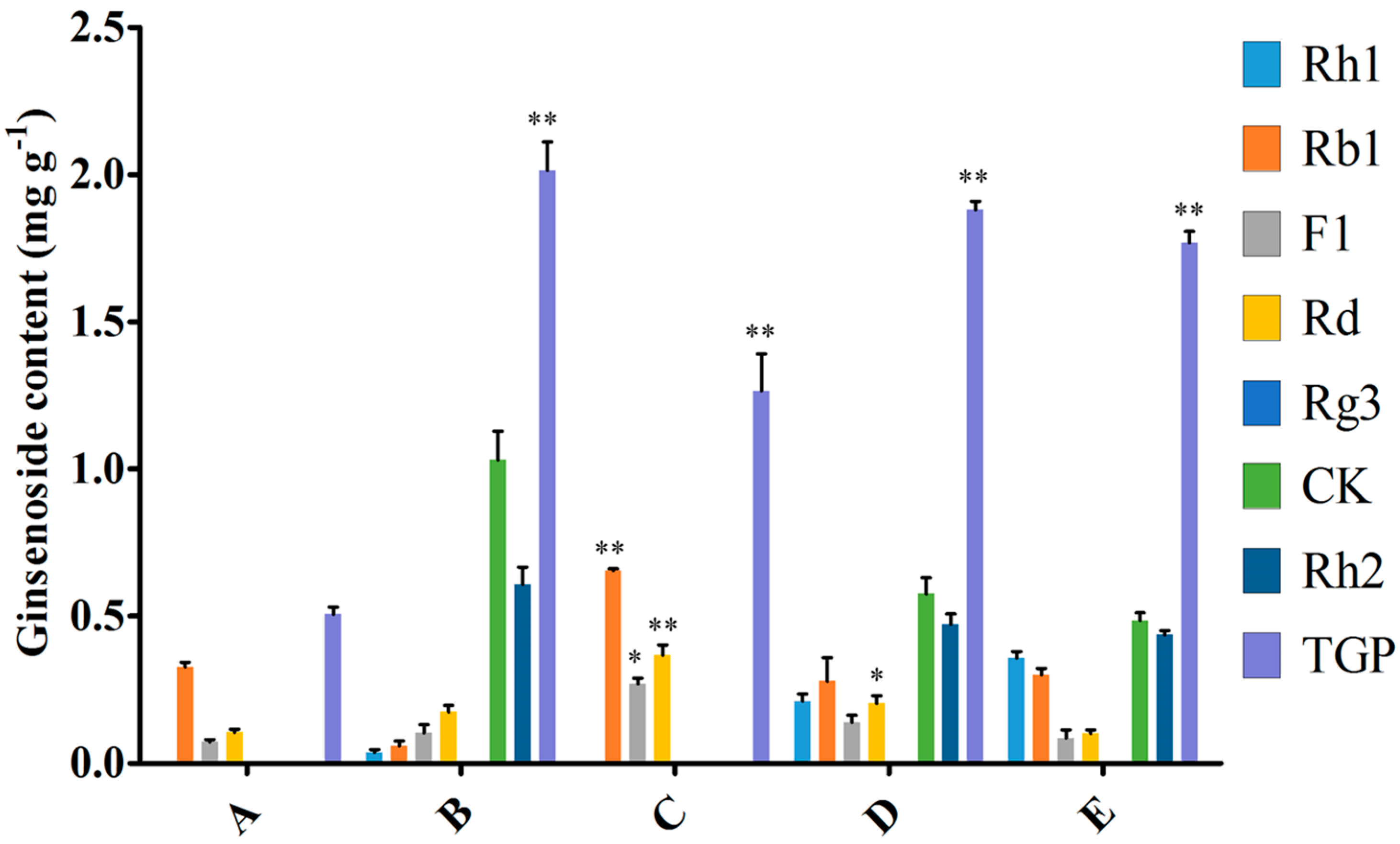
2.2. Effect of the Endophytic Elicitor on Biomass and Ginsenoside Production
2.3. Effect of MeJA, Endophytic Bacteria and Their Combination during Flask Culture
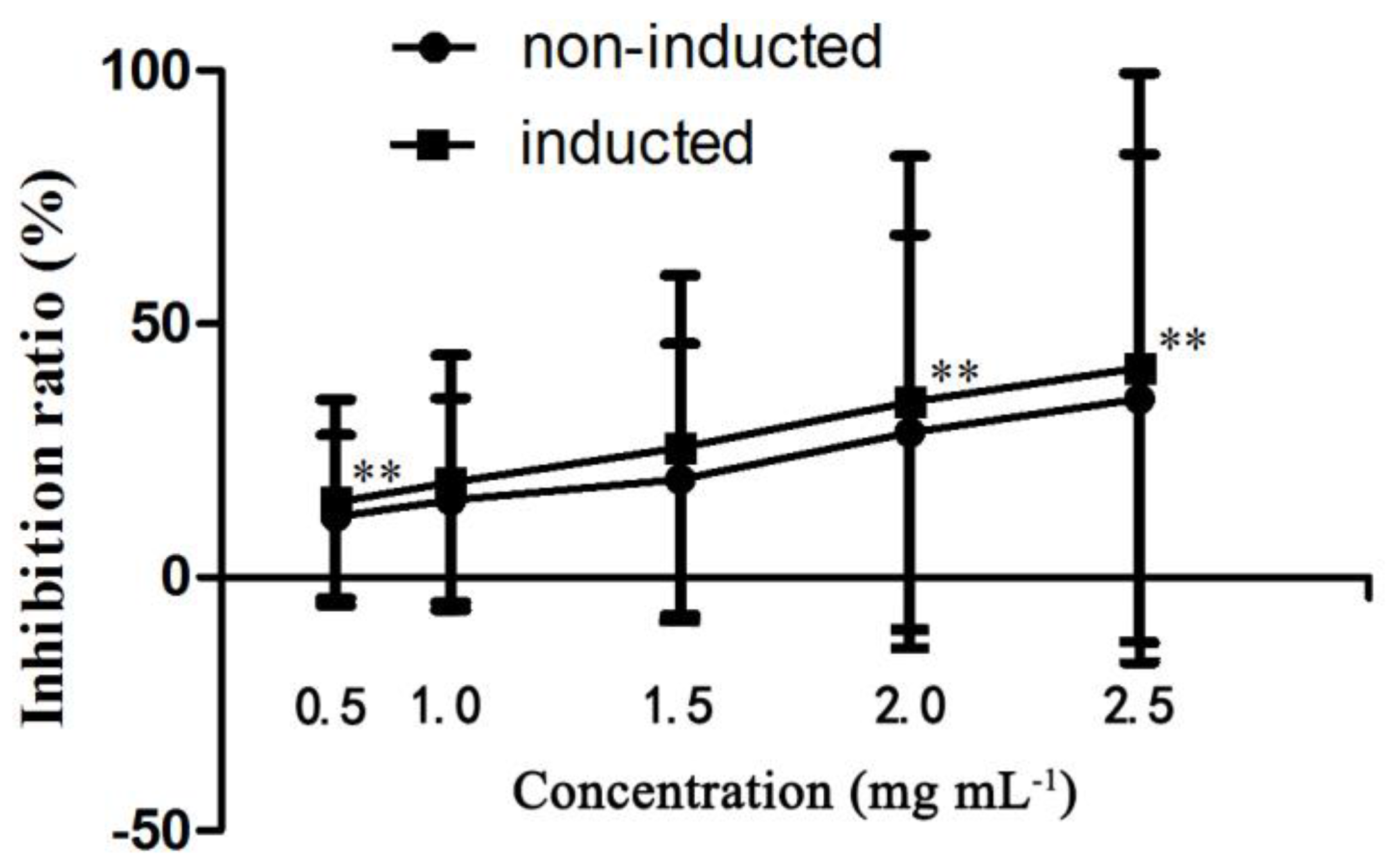
2.4. In Vitro Anticancer Activity Assay
3. Materials and Methods
3.1. Chemicals
3.2. Isolation of Endophytes in Panax ginseng Roots
3.3. Screening Bacterial Endophytes as Elicitors
3.4. Cultivation of Ginseng Adventitious Roots in an Air-Lift Bioreactor
3.5. Endophytic Bacteria Challenge Experiment
3.6. Optimization of the Elicitation Conditions
3.7. Effect of MeJA, Endophytic Bacteria and Their Combination during Flask Culture
3.8. Cell Survival Assay
3.9. Scanning Electron Microscopy (SEM)
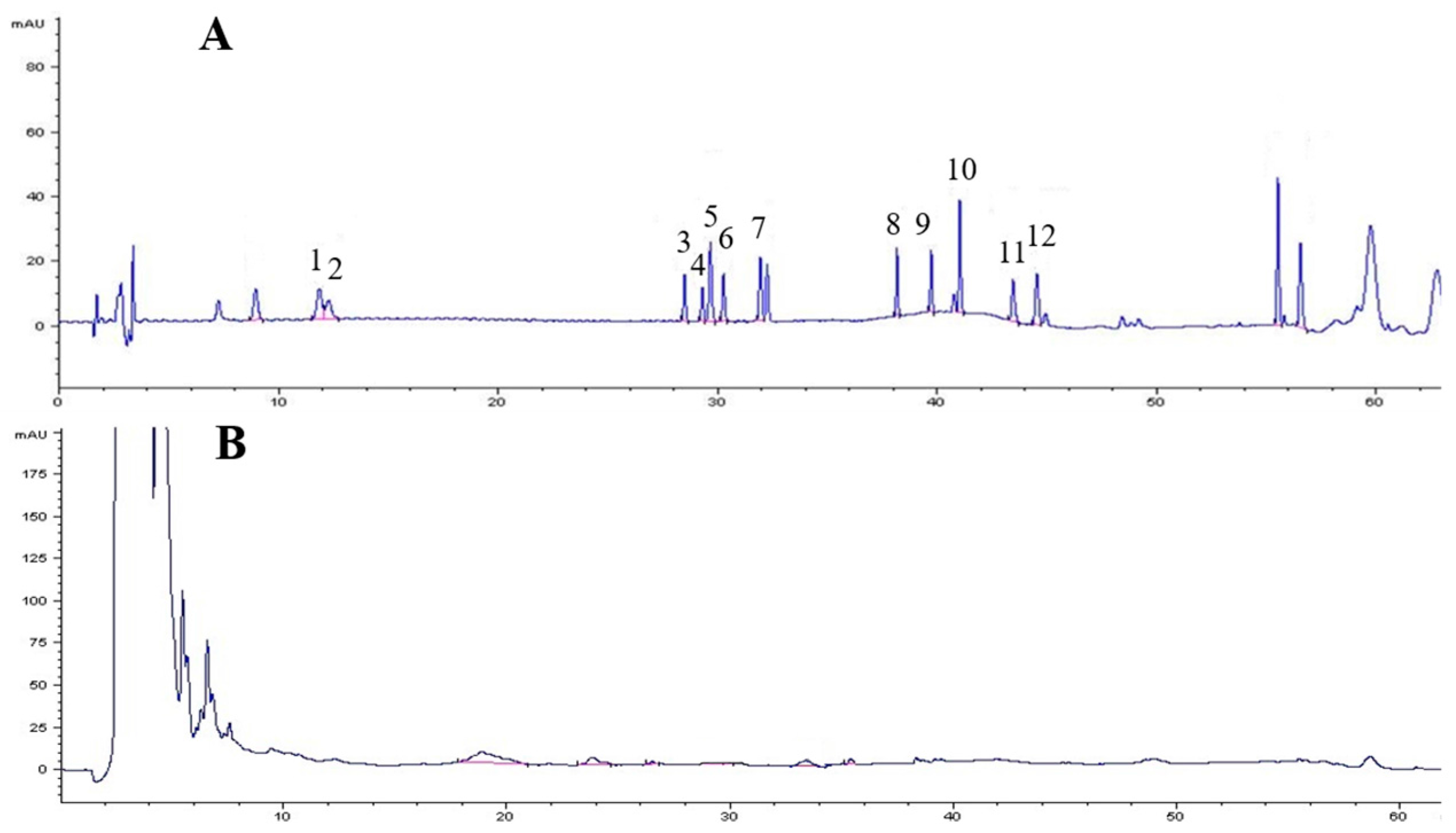
3.10. Analytical Methods
3.11. Standard Curve Preparation
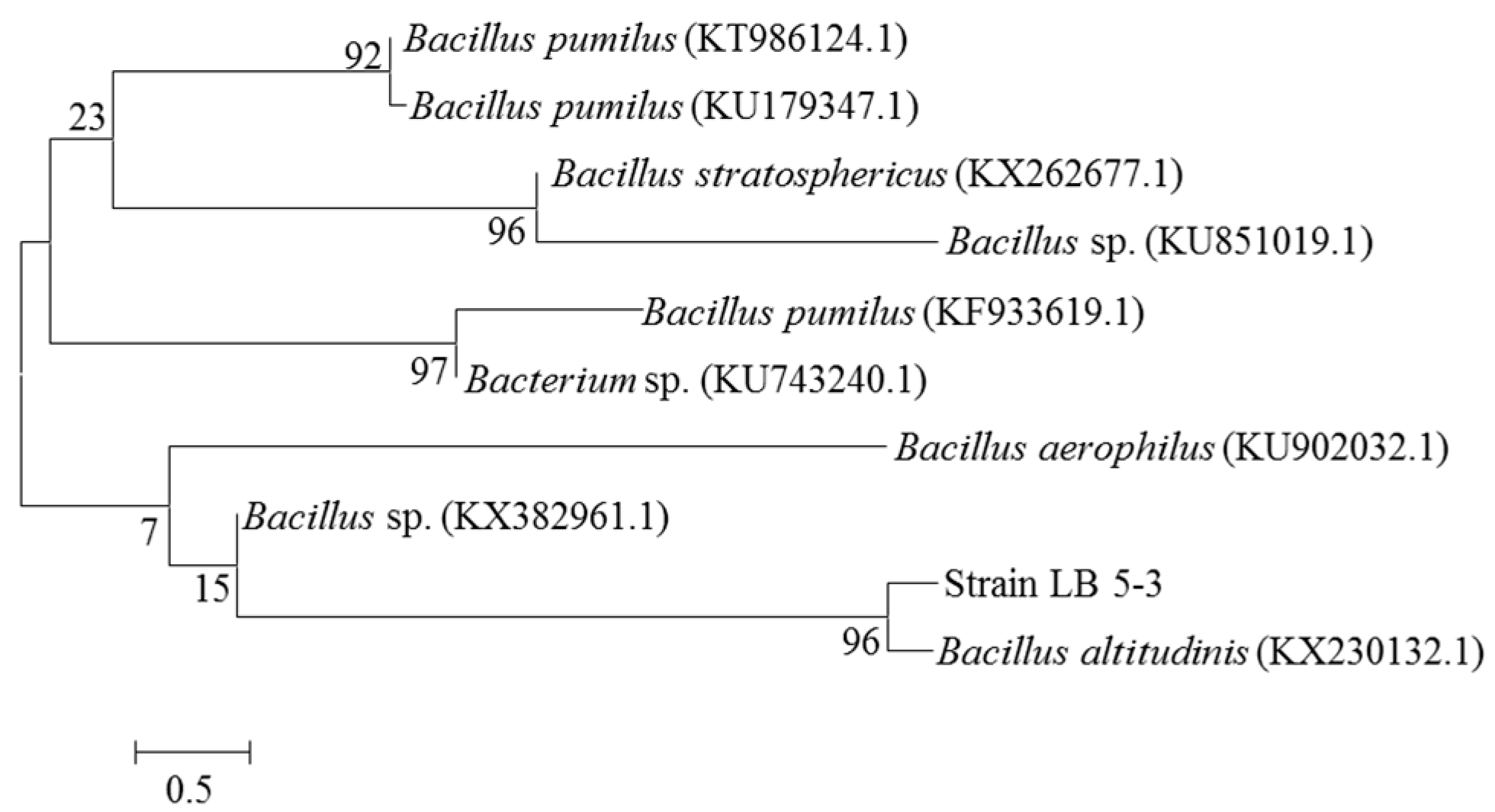
3.12. Phylogenetic Analysis
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wu, L.; Jin, Y.; Yin, C.; Bai, L. Co-Transformation of Panax major ginsenosides Rb1 and Rg1 to minor ginsenosides C-K and F1 by Cladosporium cladosporioides. J. Ind. Microbiol. Biotechnol. 2012, 39, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.D.; Chiu, C.H.; Hsu, Y.J.; Hou, C.W.; Chen, Y.M.; Huang, C.C. Changbai Mountain ginseng (Panax ginseng C.A. Mey) extract supplementation improves exercise performance and energy utilization and decreases fatigue-associated parameters in mice. Molecules 2017, 22, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Kim, Y.J.; Jang, M.G.; Joo, S.C.; Kwon, W.S.; Kim, S.Y.; Jung, S.K.; Yang, D.C. Investigation of ginsenosides in different tissues after elicitor treatment in Panax ginseng. J. Ginseng Res. 2014, 38, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Paek, K.Y.; Chakrabarty, D.; Hahn, E.J. Application of bioreactor systems for large scale production of horticultural and medicinal plants. Plant Cell Tissue Org. 2005, 81, 287–300. [Google Scholar] [CrossRef]
- Choi, S.M.; Son, S.H.; Yun, S.R.; Kwon, O.W.; Seon, J.H.; Paek, K.Y. Pilot-Scale culture of adventitious roots of ginseng in a bioreactor system. Plant Cell Tissue Org. 2000, 62, 187–193. [Google Scholar] [CrossRef]
- Yu, K.W.; Gao, W.; Hahn, E.J.; Paek, K.Y. Jasmonic acid improves ginsenoside accumulation in adventitious root culture of Panax ginseng C.A. Meyer. Biochem. Eng. J. 2002, 11, 211–215. [Google Scholar] [CrossRef]
- Yu, K.W.; Murthy, H.N.; Jeong, C.S.; Hahn, E.J.; Paek, K.Y. Organic germanium stimulates the growth of ginseng adventitious roots and ginsenoside production. Process Biochem. 2005, 40, 2959–2961. [Google Scholar] [CrossRef]
- Ali, M.B.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Differential responses of anti-oxidants enzymes, lipoxygenase activity, ascorbate content and the production of saponins in tissue cultured root of mountain Panax ginseng C.A. Mayer and Panax quinquefolium L. in bioreactor subjected to methyl jasmonate stress. Plant Sci. 2005, 169, 83–92. [Google Scholar]
- Ali, M.B.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate and salicylic acid elicitation induces ginsenosides accumulation, enzymatic and non-enzymatic antioxidant in suspension culture Panax ginseng roots in bioreactors. Plant Cell Rep. 2006, 25, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, Q.; Zang, P.; Li, X.; Ji, Q.; He, Z.; Zhao, Y.; Yang, H.; Zhao, X.; Zhang, L. An endophytic bacterium isolated from Panax ginseng C.A. Meyer enhances growth, reduces morbidity, and stimulates ginsenoside biosynthesis. Phytochem. Lett. 2015, 125, 132–138. [Google Scholar] [CrossRef]
- Wang, J.W.; Zhang, Z.; Tan, R.X. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp. Biotechnol. Lett. 2001, 23, 857–860. [Google Scholar] [CrossRef]
- Song, X.L.; Wu, H.; Piao, X.C.; Yin, Z.H.; Yin, C.R. Microbial transformation of ginsenosides extracted from Panax ginseng adventitious roots in an airlift bioreactor. Electron. J. Biotechnol. 2017, 26, 20–26. [Google Scholar] [CrossRef]
- Venugopalan, A.; Srivastava, S. Enhanced camptothecin production by ethanol addition in the suspension culture of the endophyte, Fusarium solani. Bioresour. Technol. 2015, 188, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.F.M.; Coutte, F.; Ravallec, R.; Dhulster, P.; Tournier-Couturier, L.; Jacques, P. An improvement of surfactin production by B. subtilis BBG131 using design of experiments in microbioreactors and continuous process in bubbleless membrane bioreactor. Bioresour. Technol. 2016, 218, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. Elicitors’ action modes and their application in plant tissue culture. Chin. Bull. Bot. 1999, 16, 11–18. [Google Scholar]
- Venugopalan, A.; Potunuru, U.R.; Dixit, M.; Srivastava, S. Effect of fermentation parameters, elicitors and precursors on camptothecin production from the endophyte Fusarium solani. Bioresour. Technol. 2016, 206, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.T.; Li, D.F.; Ding, K.; Che, R.Q.; Xu, J.W.; Zhao, P.; Li, T.; Ma, H.X.; Yu, X.Y. Production of biomass and lipids by the oleaginous microalgae Monoraphidium sp. QLY-1 through heterotrophic cultivation and photo-chemical modulator induction. Bioresour. Technol. 2016, 211, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, S.; Kim, Y.J.; Yang, D.C. Production of ginseng saponins: Elicitation strategy and signal transductions. Appl. Microbiol. Biotechnol. 2015, 99, 6987–6996. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Jin, M.Y.; Paek, K.Y.; Piao, X.C.; Lian, M.L. An efficient strategy for enhancement of bioactive compounds by protocorm-like body culture of Dendrobium candidum. Ind. Crops Prod. 2016, 84, 121–130. [Google Scholar] [CrossRef]
- Kim, Y.S.; Hahn, E.J.; Murthy, H.N.; Paek, K.Y. Adventitious root growth and ginsenoside accumulation in Panax ginseng cultures as affected by methyl jasmonate. Biotechnol. Lett. 2004, 26, 1619–1622. [Google Scholar] [CrossRef] [PubMed]
- Creelman, R.A.; Mullet, J.E. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. USA 1995, 92, 4114–4119. [Google Scholar] [CrossRef] [PubMed]
- Kuźma, Ł.; Bruchajzer, E.; Wysokińska, H. Methyl jasmonate effect on diterpenoid accumulation in Salvia sclarea hairy root culture in shake flasks and sprinkle bioreactor. Enzyme Microb. Technol. 2009, 44, 406–410. [Google Scholar] [CrossRef]
- Wang, J.; Gao, W.Y.; Zhang, J.; Huang, T.; Wen, T.T.; Huang, L.Q. Combination effect of lactoalbumin hydrolysate and methyl jasmonate on ginsenoside and polysaccharide production in Panax quinquefolium L. cells cultures. Acta Physiol. Plant 2011, 33, 861–866. [Google Scholar] [CrossRef]
- Thanh, N.T.; Murthy, H.N.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate elicitation enhanced synthesis of ginsenoside by cell suspension cultures of Panax ginseng in 5-L balloon type bubble bioreactors. Appl. Microbiol. Biotechnol. 2005, 67, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, W.B.; Li, X.Y.; Piao, X.C.; Jiang, J.; Lian, M.L. Pathogenic fungal elicitors enhance ginsenoside biosynthesis of adventitious roots in Panax quinquefolius during bioreactor culture. Ind. Crops Prod. 2016, 94, 729–735. [Google Scholar] [CrossRef]
- Kim, D.Y.; Yuan, H.D.; Chung, I.K.; Chung, S.H. Compound K, intestinal metabolite of ginsenoside, attenuates hepatic lipid accumulation via AMPK activation in human hepatoma cells. J. Agric. Food Chem. 2009, 57, 1532–1537. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.H.; Kim, H.; Hong, M.J.; Yang, M.H.; Kim, J.W.; Yoo, H.; Yang, H.; Park, J.H.; Sung, S.H.; Kim, H.P.; et al. Stereoisomer-Specific anticancer activities of ginsenoside Rg3 and Rh2 in HepG2 cells: Disparity in cytotoxicity and autophagy-inducing effects due to 20(S)-epimers. Biol. Pharm. Bull. 2015, 38, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Rodriguez-Tudela, J.L.; Chryssanthou, E.; Petrikkou, E.; Mosquera, J.; Denning, D.W.; Cuenca-Estrella, M. Interlaboratory evaluation of hematocytometer method of inoculum preparation for testing antifungal susceptibilities of filamentous fungi. J. Clin. Microbiol. 2003, 41, 5236–5237. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the ginseng adventitious root extracts are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ginsenoside | Linear Range (mg mL−1) | Linear Equation | Correlation Coefficient (r2) |
|---|---|---|---|
| Rh1 | 0.002–2.000 | y = 2429.3x + 21.901 | 0.999 |
| Rb1 | 0.004–4.000 | y = 1598.8x + 35.703 | 0.9988 |
| F1 | 0.004–4.000 | y = 2630.8x + 10.136 | 0.9992 |
| Rd | 0.004–4.000 | y = 1604.2x + 20.314 | 0.9989 |
| Rg3 | 0.002–2.000 | y = 2064.8x + 42.395 | 0.9987 |
| CK | 0.002–1.500 | y = 2961.7x + 12.920 | 0.9992 |
| Rh2 | 0.002–1.500 | y = 2945.2x + 42.138 | 0.999 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Wu, H.; Yin, Z.; Lian, M.; Yin, C. Endophytic Bacteria Isolated from Panax ginseng Improves Ginsenoside Accumulation in Adventitious Ginseng Root Culture. Molecules 2017, 22, 837. https://doi.org/10.3390/molecules22060837
Song X, Wu H, Yin Z, Lian M, Yin C. Endophytic Bacteria Isolated from Panax ginseng Improves Ginsenoside Accumulation in Adventitious Ginseng Root Culture. Molecules. 2017; 22(6):837. https://doi.org/10.3390/molecules22060837
Chicago/Turabian StyleSong, Xiaolin, Hao Wu, Zhenhao Yin, Meilan Lian, and Chengri Yin. 2017. "Endophytic Bacteria Isolated from Panax ginseng Improves Ginsenoside Accumulation in Adventitious Ginseng Root Culture" Molecules 22, no. 6: 837. https://doi.org/10.3390/molecules22060837
APA StyleSong, X., Wu, H., Yin, Z., Lian, M., & Yin, C. (2017). Endophytic Bacteria Isolated from Panax ginseng Improves Ginsenoside Accumulation in Adventitious Ginseng Root Culture. Molecules, 22(6), 837. https://doi.org/10.3390/molecules22060837