
The Cytotoxicity of the Ajoene Analogue BisPMB in WHCO1 Oesophageal Cancer Cells Is Mediated by CHOP/GADD153


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
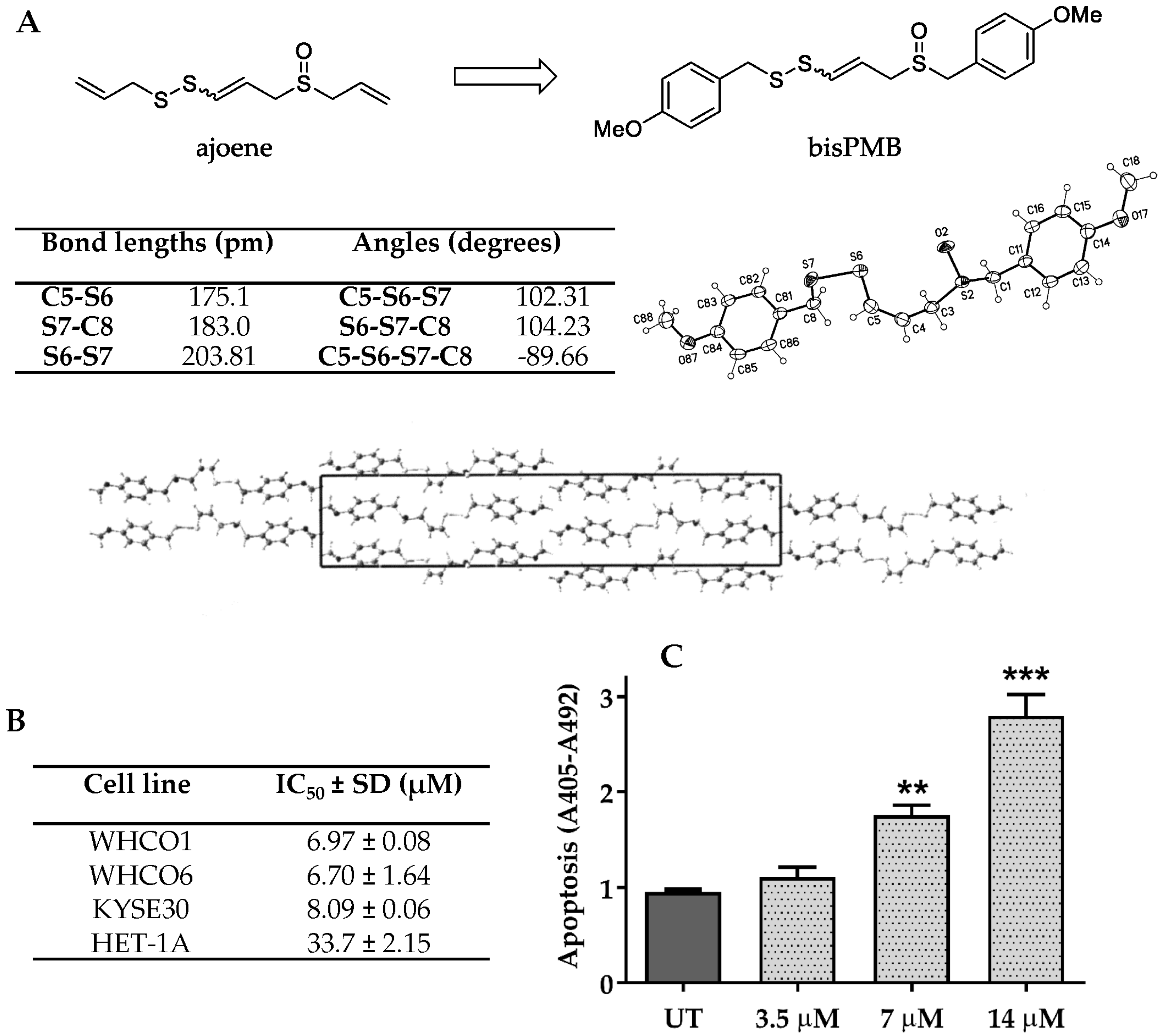
2.1. The Ajoene Analogue BisPMB Is Cytotoxic to Oesophageal Cancer Cells
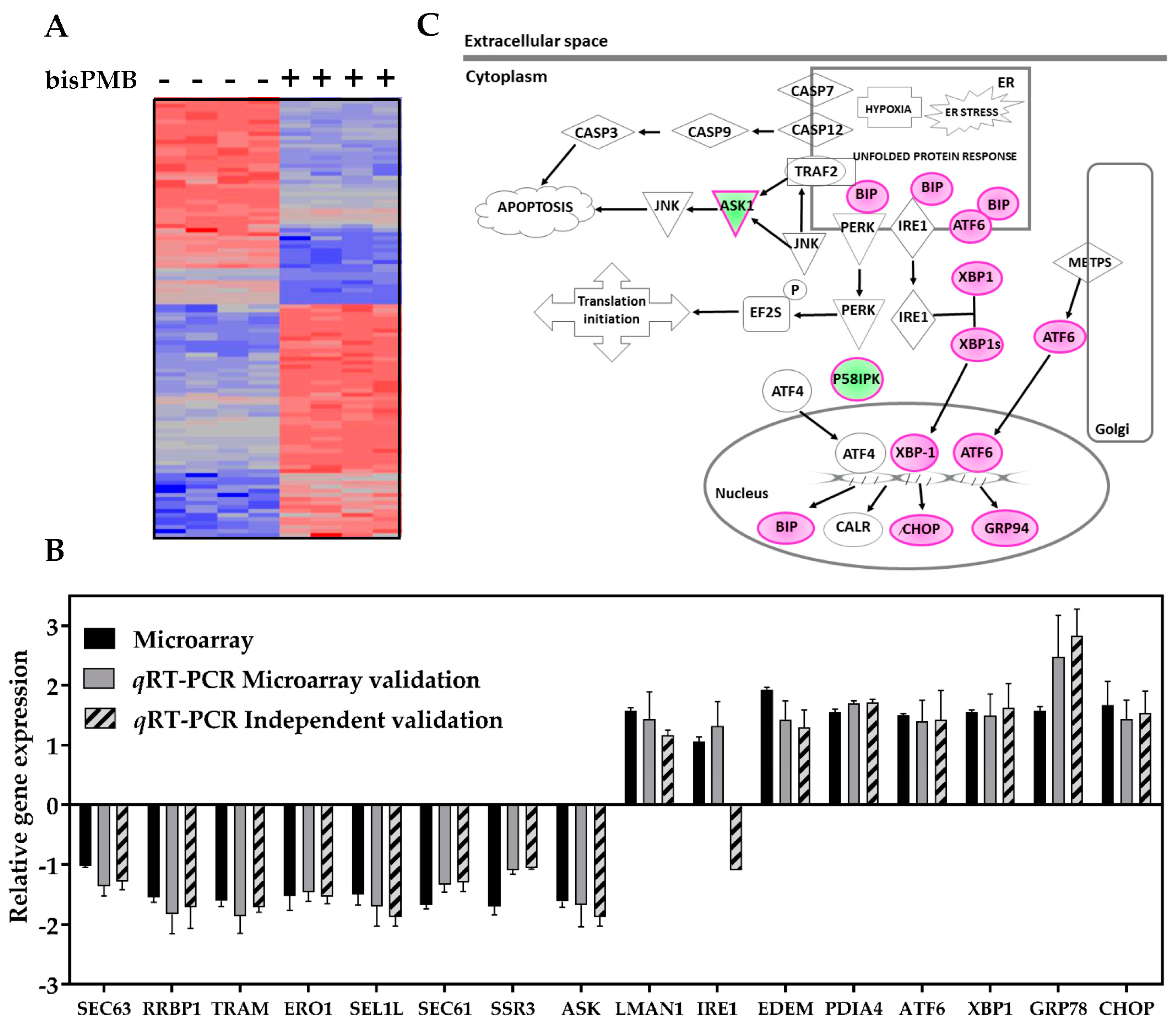
2.2. Transcriptional Profiling of BisPMB-Treated WHCO1 Cells
2.2.1. Validation of Selected Microarray DEGs
2.2.2. Gene Ontology Analysis
2.2.3. Pathway Analysis
2.2.4. Network Analysis
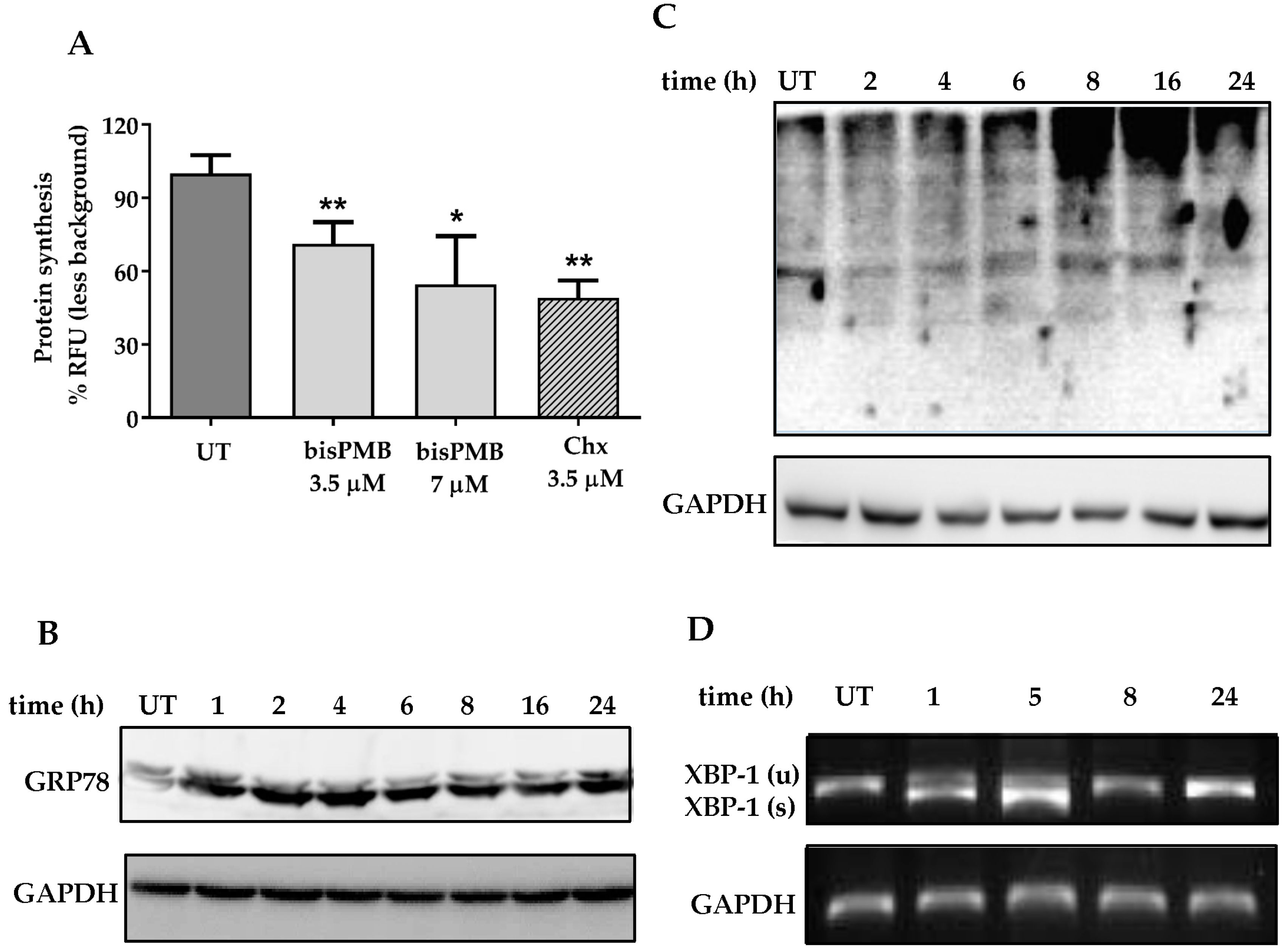
2.3. BisPMB Activates the Unfolded Protein Response
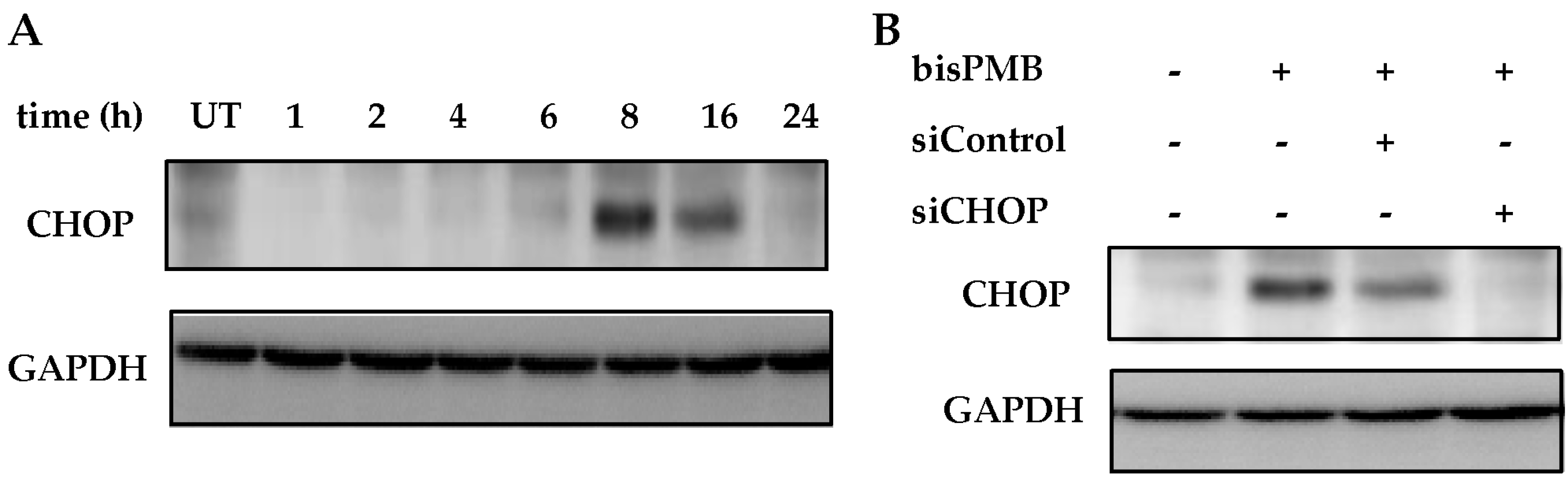
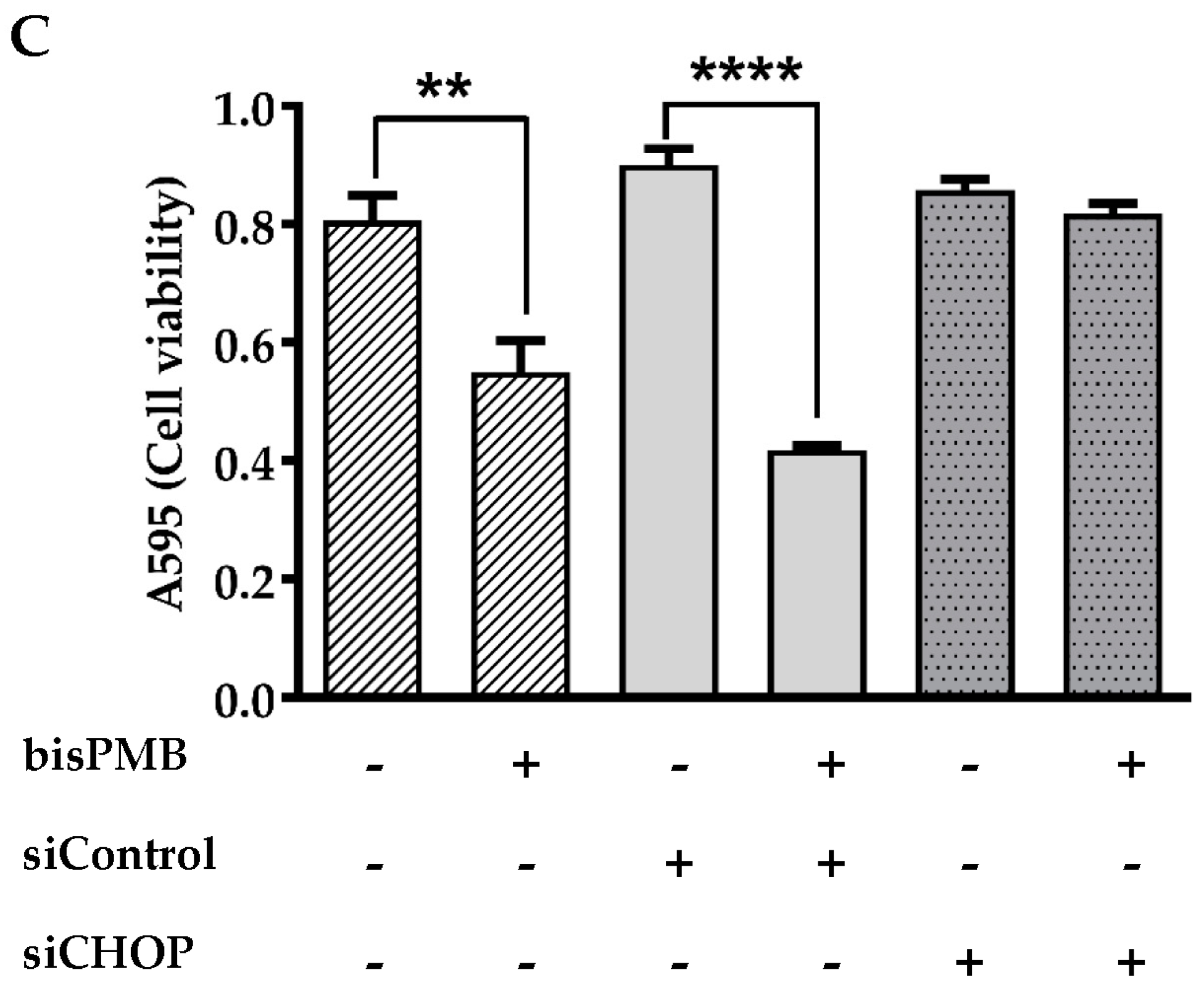
2.4. CHOP/GADD153 Is a Central Regulator of BisPMB-Induced Cytotoxicity
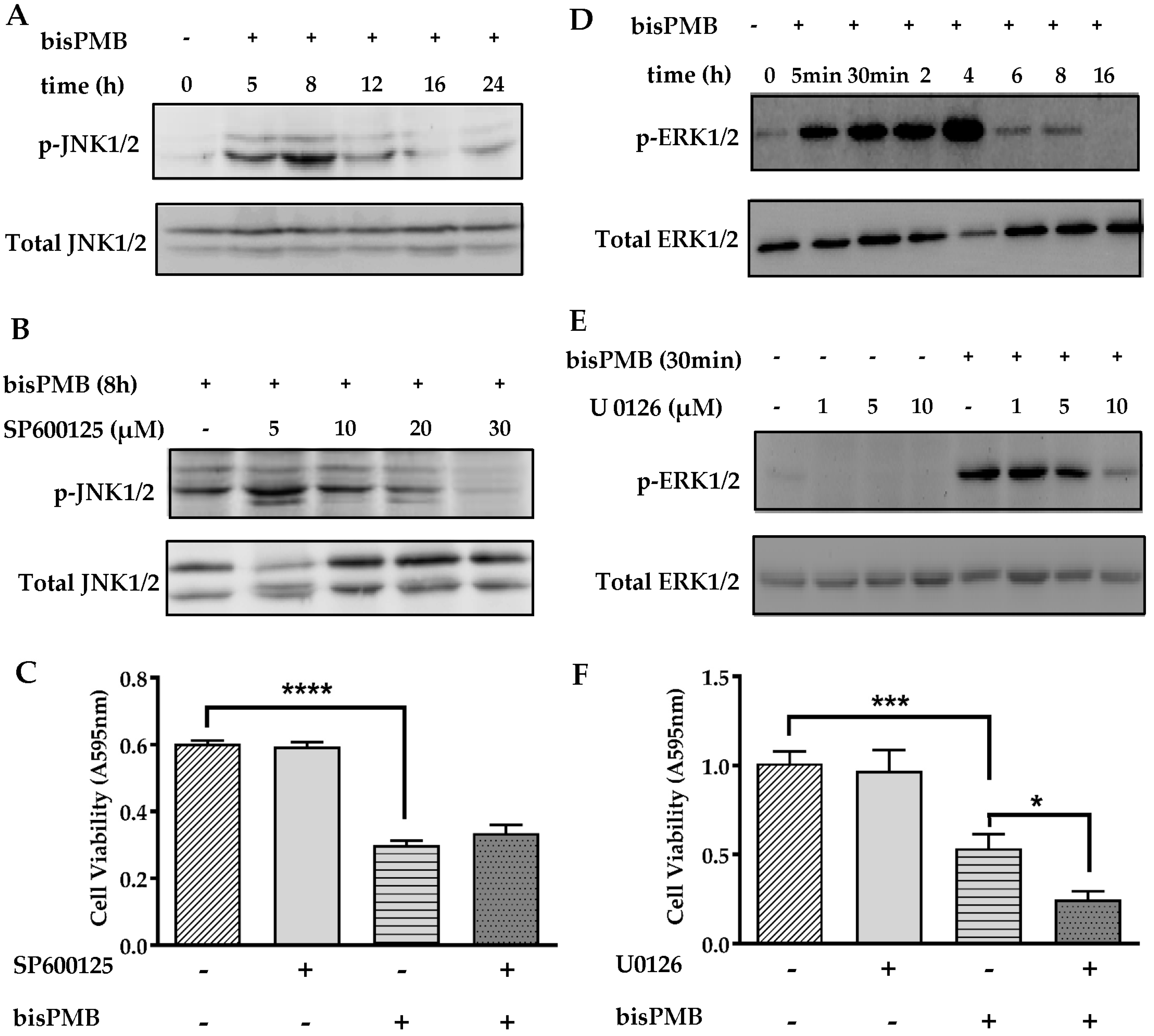
2.5. The Role of the MAPK Signaling Pathways in BisPMB-Induced Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. X-ray Crystallography
4.2. Cell Lines and Treatments
4.3. Cellular Viability Quantification
4.4. Apoptosis Quantification
4.5. Protein Synthesis Quantification
4.6. RNA Preparation and qRT-PCR
4.7. Affymetrix Human Gene ST 2.0 Array and Bioinformatics Analysis
4.8. Transient Knock Down of CHOP/GADD153
4.9. Immunoblot Analysis
4.10. Statistical Analysis
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Fleischauer, A.T.; Arab, L. Garlic and cancer: A critical review of the epidemiologic literature. J. Nutr. 2001, 3S, 1032S–1040S. [Google Scholar]
- Kim, J.Y.; Kwon, O. Garlic intake and cancer risk: An analysis using the Food and Drug Administration's evidence-based review system for the scientific evaluation of health claims. Am. J. Clin. Nutr. 2009, 89, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.N.; Williams, D.B.; Cobiac, L.; Head, R.J. Does garlic reduce risk of colorectal cancer? A systematic review. J. Nutr. 2007, 137, 2264–2269. [Google Scholar] [PubMed]
- Nicastro, H.L.; Ross, S.A.; Milner, J.A. Garlic and Onions: Their Cancer Prevention Properties. Cancer Prev. Res. 2015, 8, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, G.; Kaschula, C.H. The immunomodulation and anti-inflammatory effects of garlic organosulfur compounds in cancer chemoprevention. Anti-Cancer Agent. Med. Chem. 2014, 14, 233–240. [Google Scholar] [CrossRef]
- Scharfenberg, K.; Wagner, R.; Wagner, K.G. The cytotoxicity effect of ajoene, a natural product from garlic, investigated with different cell lines. Cancer Lett. 1990, 53, 103–108. [Google Scholar] [CrossRef]
- Taylor, P.; Noriega, R.; Farah, C.; Abad, M.-J.; Arsenak, M.; Apitz, R. Ajoene inhibits both primary tumor growth and metastasis of B16/BL6 melanoma cells in C57BL/6 mice. Cancer Lett. 2006, 239, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ciu, J.R.; Ye, Y.; Min, J.M.; Zhang, L.H.; Wang, K.; Gares, M.; Cros, J.; Wright, M.; Leung-Tack, J. Antitumor activity of Z-ajoene, a natural compound purified from garlic: antimitotic and microtubule-interaction properties. Carcinogenesis 2002, 23, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Kaschula, C.H.; Hunter, R.; Hassan, H.T.; Stellenboom, N.; Cotton, J.; Zhai, X.Q.; Parker, M.I. Anti-Proliferative Activity of Synthetic Ajoene Analogues on Cancer Cell Lines. Anti-Cancer Agent. Med. Chem. 2011, 11, 260–266. [Google Scholar] [CrossRef]
- Block, E.; Ahmad, S.; Catalfamo, J.L.; Jain, M.K.; Apitz-Castro, R. The chemistry of alkyl thiosulfinate esters. 9. Antithrombotic organosulfur compounds from garlic: Structural, mechanistic, and synthetic studies. J. Am. Chem. Soc. 1986, 108, 7045–7055. [Google Scholar] [CrossRef]
- Yang, C.S.; Smith, T.; Hong, J.Y. Cytochrome P-450 enzymes as targets for chemoprevention against chemical carcinogenesis and toxicity: Opportunities and limitations. Cancer Res. 1994, 54, 1982S–1986S. [Google Scholar] [PubMed]
- Bhuiyan, A.I.; Papajani, V.T.; Paci, M.; Melino, S. Glutathione-garlic sulfur conjugates: Slow hydrogen sulfide releasing agents for therapeutic applications. Molecules 2015, 20, 1731–1750. [Google Scholar] [CrossRef] [PubMed]
- Hosono, T.; Fukao, T.; Ogihara, J.; Ito, Y.; Shiba, H.; Seki, T.; Ariga, T. Diallyl trisulfidesuppresses the proliferation and induces apoptosis of human colon cancer cells through oxidative modification of β-tubulin. J. Biol. Chem. 2005, 280, 41487–41493. [Google Scholar] [CrossRef] [PubMed]
- Gallwitz, H.; Bonse, S.; Martinez-Cruz, A.; Schlichting, I.; Schumacher, K.; Krauth-Siegel, R.L. Ajoene is an Inhibitor and Subversive Substrate of Human Glutathione Reductase and Trypanosoma cruzi Trypanothione Reductase: Crystallographic, Kinetic, and Spectroscopic Studies. J. Med. Chem. 1999, 42, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Rabinkov, A.; Miron, T.; Mirelman, D.; Wilchek, M.; Glozman, S.; Yavin, E.; Weiner, L. S-Allylmercaptoglutathione: The reaction product of allicin with glutathione possesses SH-modifying and antioxidant properties. Biochim. Biophys. Acta 2000, 1499, 144–153. [Google Scholar] [CrossRef]
- Nepravishta, R.; Sabelli, R.; Iorio, E.; Micheli, L.; Paci, M.; Melino, S. Oxidative species and S-glutathionyl conjugates in the apoptosis induction by allyl thiosulfate. FEBS J. 2012, 279, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Sabelli, R.; Iorio, E.; De Martino, A.; Podo, F.; Ricci, A.; Viticchie, G.; Rotilio, G.; Paci, M.; Melino, S. Rhodanese-thioredoxin system and allyl sulfur compounds. FEBS J. 2008, 275, 3884–3899. [Google Scholar] [CrossRef] [PubMed]
- Kaschula, C.H.; Hunter, R.; Cotton, J.; Tuveri, R.; Ngarande, E.; Dzobo, K.; Schäfer, G.; Siyo, V.; Lang, D.; Kusza, D.A.; et al. The garlic compound ajoene targets protein folding in the endoplasmic reticulum of cancer cells. Mol. Carcinog. 2016, 55, 1213–1228. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Hunter, R.; Stellenboom, N.; Kusza, D.A.; Parker, M.I.; Hammouda, A.N.; Jackson, G.; Kaschula, C.H. The cytotoxicity of garlic-related disulphides and thiosulfonates in WHCO1 oesophageal cancer cells is dependent on S-thiolation and not production of ROS. Biochim. Biophys. Acta 2016, 1860, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Dirsch, V.M.; Gerbes, A.L.; Volmar, A.M. Ajoene, a compound of garlic, induces apoptosis in human promyleoleukemic cells, accompanied by generation of reactive oxygen species and activation of nuclear factor kB. Mol. Pharmacol. 1998, 53, 402–407. [Google Scholar] [PubMed]
- Liang, D.; Wu, H.; Wong, M.W.; Huang, D. Diallyl Trisulfide Is a Fast H2S Donor, but Diallyl Disulfide is a Slow One: The Reaction Pathways and Intermediates of Glutathione with Polysulfides. Org. Lett. 2015, 17, 4196–4199. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, N.S.; Anilakumar, K.R.; Singh, O.V. Diallyl disulfide causes caspase-dependent apoptosis in human cancer cells through a Bax-triggered mitochondrial pathway. J. Nutr. Biochem. 2010, 21, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Dirsch, V.M.; Antlsperger, D.S.M.; Hentze, H.; Vollmar, A.M. Ajoene, an experimental anti-leukemic drug: Mechanism of cell death. Leukemia 2002, 16, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Antlsperger, D.S.M.; Dirsch, V.M.; Ferreira, D.; Su, J.-L.; Kuo, M.-L.; Vollmar, A.M. Ajoene-induced cell death in human promyeloleukemic cells does not require JNK but is amplified by the inhibition of ERK. Oncogene 2003, 22, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Park, H.; Zhao, H.Y.; Jeon, R.; Ryu, J.H.; Kim, W.Y. Systemic approaches identify a garlic-derived chemical, Z-ajoene, as a glioblastoma multiforme cancer stem cell-specific targeting agent. Mol. Cell 2014, 37, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Pinto, J.T.; Soh, J.-W.; Deguchi, A.; Gundersen, G.G.; Palazzo, A.; Yoon, J.-T.; Shirin, H.; Weinstein, I.B. Induction of Apoptosis by the Garlic-Derived Compound S-Allylmercaptocysteine (SAMC) is Associated with Microtubule Depolymerization and c-Jun NH2-Terminal Kinase 1 Activation1. Cancer Res. 2003, 63, 6825–6837. [Google Scholar] [PubMed]
- Filomeni, G.; Aquilano, K.; Rotilio, G.; Ciriolo, M.R. Reactive oxygen species-dependent c-Jun NH2-terminal kinase/c-Jun signaling cascade mediates neuroblastoma cell death induced by diallyl disulfide. Cancer Res. 2003, 63, 5940–5949. [Google Scholar] [PubMed]
- Xiao, D.; Choi, S.; Johnson, D.E.; Vogel, V.G.; Johnson, C.S.; Trump, D.L.; Lee, Y.J.; Singh, S.V. Diallyl trisulfide-induced apoptosis in human prostate cancer cells involves c-Jun N-terminal kinase and extracellular-signal regulated kinase-mediated phosphorylation of Bcl-2. Oncogene 2004, 23, 5594–5606. [Google Scholar] [CrossRef] [PubMed]
- Kelkel, M.; Cerella, C.; Mack, F.; Schneider, T.; Jacob, C.; Schumacher, M.; Dicato, M.; Diederich, M. ROS-independent JNK activation and multisite phosphorylation of Bcl-2 link diallyl tetrasulfide-induced mitotic arrest to apoptosis. Carcinogenesis 2012, 33, 2162–2171. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Hsieh, S.C.; Yang, J.H.; Lin, S.Y.; Sheen, L.Y. Diallyl trisulfide induces apoptosis of human basal cell carcinoma cells via endoplasmic reticulum stress and the mitochondrial pathway. Nutr. Cancer 2012, 64, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Saidu, N.E.; Touma, R.; Asali, I.A.; Jacob, C.; Montenarh, M. Diallyl tetrasulfane activates both the eIF2alpha and Nrf2/HO-1 pathways. Biochim. Biophys. Acta 2013, 1830, 2214–2225. [Google Scholar] [CrossRef] [PubMed]
- Saidu, N.E.; Abu Asali, I.; Czepukojc, B.; Seitz, B.; Jacob, C.; Montenarh, M. Comparison between the effects of diallyl tetrasulfide on human retina pigment epithelial cells (ARPE-19) and HCT116 cells. Biochim. Biophys. Acta 2013, 1830, 5267–5276. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Lawson, L.D.; Milner, J.A. Allyl sulfides from garlic suppress the in vitro proliferation of human A549 lung tumor cells. Nutr. Cancer 1997, 29, 152–156. [Google Scholar]
- Sundaram, S.G.; Milner, J.A. Diallyl disulfide inhibits the proliferation of human tumor cells in culture. Biochim. Biophys. Acta 1996, 1315, 15–20. [Google Scholar] [CrossRef]
- Chen, W.-C.; Hsu, S.-S.; Chou, C.-T.; Kuo, C.-C.; Huang, J.-K.; Fang, Y.-C.; Chang, H.-T.; Tsai, J.-Y.; Liao, W.-C.; Wang, B.-W.; et al. Effect of diallyl disulfide on Ca2+ movement and viability in PC3 human prostate cancer cells. Toxicol. In Vitro 2011, 25, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-S.; Chen, G.-W.; Hsia, T.-C.; Ho, H.-C.; Ho, C.-C.; Lin, M.-W.; Lin, S.-S.; Yeh, R.-D.; Ip, S.-W.; Lu, H.-F.; et al. Diallyl disulfide induces apoptosis in human colon cancer cell line (COLO 205) through the induction of reactive oxygen species, endoplasmic reticulum stress, caspases casade and mitochondrial-dependent pathways. Food. Chem. Toxicol. 2009, 47, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Banik, N.L.; Patel, S.J.; Ray, S.K. Garlic compounds induced calpain and intrinsic caspase cascade for apoptosis in human malignant neuroblastoma SH-SY5Y cells. Apoptosis 2007, 12, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Su, S.-J.; Chang, K.-L.; Huang, M.-W.; Kuo, S.-Y. The garlic ingredient diallyl sulfide induces Ca2+ mobilization in Madin-Darby canine kidney cells. Food Chem. Toxicol. 2009, 47, 2344–2350. [Google Scholar] [CrossRef] [PubMed]
- Park, E.K.; Kwon, K.B.; Park, K.I.; Park, B.H.; Jhee, E.C. Role of Ca2+ in diallyl disulfide-induced apoptotic cell death of HCT-15 cells. Exp. Mol. Med. 2002, 34, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, S.G.; Milner, J.A. Diallyl disulfide induces apoptosis of human colon tumor cells. Carcinogenesis 1996, 17, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Kaschula, C.H.; Hunter, R.; Stellenboom, N.; Caira, M.R.; Winks, S.; Ogunleye, T.; Richards, P.; Cotton, J.; Zilbeyaz, K.; Wang, Y.; et al. Structure-Activity Studies into the Anti-Proliferation Activity of Ajoene Derivatives in WHCO1 Oesophageal Cancer Cells. Eur. J. Med. Chem. 2012, 50, 236–254. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.; Kaschula, C.H.; Parker, M.I.; Caira, M.R.; Richards, P.; Travis, S.; Taute, F.; Qwebani, T. Substituted ajoenes as novel anti-cancer agents. Bioorg. Med. Chem. Lett. 2008, 18, 5277–5279. [Google Scholar] [CrossRef] [PubMed]
- Veale, R.B.; Thornley, A.L. Increased single class low-affinity EGF receptors expressed by human oesophageal squamous carcinoma cell lines. S. Afr. J. Sci. 1989, 85, 375–379. [Google Scholar]
- Shimada, Y.; Imamura, M.; Wagata, T.; Yamaguchi, N.; Tobe, T. Characterization of 21 newly established esophageal cancer cell lines. Cancer 1992, 69, 277–284. [Google Scholar] [CrossRef]
- Tanaka, N.; Kimura, H.; Faried, A.; Sakai, M.; Sano, A.; Inose, T.; Sohda, M.; Okada, K.; Nakajima, M.; Miyazaki, T.; et al. Quantitative analysis of cisplatin sensitivity of human esophageal squamous cancer cell lines using in-air micro-PIXE. Cancer Sci. 2010, 101, 1487–1492. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic reticulum stress: Cell life and death decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.; Teodoro, T.; Volchuk, A. Endoplasmic reticulum stress: Signaling the unfolded protein response. Physiology 2007, 22, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D. A direct approach to false discovery rates. J. R. Stat. Soc. Ser. 2002, 64, 479–498. [Google Scholar] [CrossRef]
- Gebhardt, R.; Beck, H.; Wagner, K.G. Inhibition of chloesterol biosynthesis by allicin and ajoene in rat hepatocytes and HepG2 cells. Biochim. Biophys. Acta 1994, 1213, 57–62. [Google Scholar] [CrossRef]
- Sendl, A.; Schliack, M.; Loser, R.; Stanislaus, F.; Wagner, H. Inhibition of cholesterol synthesis in vitro by extracts and isolated compounds prepared from garlic and wild garlic. Atherosclerosis 1992, 94, 79–85. [Google Scholar] [CrossRef]
- Meusser, B.; Hirsch, C.; Jarosch, E.; Sommer, T. ERAD: The long road to destruction. Nat. Cell Biol. 2005, 7, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Vembar, S.S.; Brodsky, J.L. One step at a time: endoplasmic reticulum-associated degradation. Nat. Rev. Mol. Cell Biol. 2008, 9, 944–957. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods 2005, 35, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Novoa, I.; Zhang, Y.; Zeng, H.; Wek, R.; Schapira, M.; Ron, D. Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol. Cell 2000, 6, 1099–1108. [Google Scholar] [CrossRef]
- Oyadomari, S.; Mori, M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004, 11, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Darling, N.J.; Cook, S.J. The role of MAPK signalling pathways in the response to endoplasmic reticulum stress. Biochim. Biophys. Acta 2014, 1843, 2150–2163. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.J. Signal transduction by the JNK group of MAP kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef]
- Mhaidat, N.M.; Zhang, X.D.; Jiang, C.C.; Hersey, P. Docetaxel-induced apoptosis of human melanoma is mediated by activation of c-Jun NH2-terminal kinase and inhibited by the mitogen-activated protein kinase extracellular signal-regulated kinase 1/2 pathway. Clin. Cancer Res. 2007, 13, 1308–1314. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, P.; Schremser, E.J.; Cooper, G.M. B-Raf inhibits programmed cell death downstream of cytochrome c release from mitochondria by activating the MEK/Erk pathway. Mol. Cell. Biol. 1999, 19, 5308–5315. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Ellis, M.; Erlichman, C.; Lutzker, S.; Zwiebel, J. Development of rational drug combinations with investigational targeted agents. Oncologist 2010, 15, 496–499. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Miyazaki, S.; Matsuyama, S.; Takeda, M.; Kawano, M.; Nakagawa, H.; Nishimura, K.; Matsuo, S. Selection of autophagy or apoptosis in cells exposed to ER-stress depends on ATF4 expression pattern with or without CHOP expression. Biol. Open 2013, 2, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Caira, M.R.; Hunter, R.; Bourne, S.A.; Smith, V.J. Preparation, thermal behavior and solid-state structures of inclusion complexes of permethylated-β-cyclodextrin with the garlic-derived antithrombotics (E)- and (Z)-ajoene. Supramol. Chem. 2004, 16, 395–403. [Google Scholar] [CrossRef]
- Van Wart, H.E.; Lewis, A.; Scheraga, H.A.; Saeva, F.D. Disulfide bond dihedral angles from Raman spectroscopy. PNAS 1973, 70, 2619–2623. [Google Scholar] [CrossRef] [PubMed]
- Shefter, E.; Kotick, M.P.; Bardos, T.J. Crystal and molecular structure of 5-[1-(2‘-deoxy-alpha-d-ribofuranosyl)uracilyl] disulfide. J. Pharm. Sci. 1967, 56, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Schubert, U.; Anton, L.C.; Gibbs, J.; Norbury, C.C.; Yewdell, J.W.; Bennink, J.R. Rapid degradation of a large fraction of newly synthesized proteins by proteasomes. Nature 2000, 404, 770–774. [Google Scholar] [PubMed]
- Schroder, M.; Kaufman, R.J. ER stress and the unfolded protein response. Mutat. Res. 2005, 569, 29–63. [Google Scholar] [CrossRef] [PubMed]
- Urano, F.; Wang, X.; Bertolotti, A.; Zhang, Y.; Chung, P.; Harding, H.P.; Ron, D. Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science 2000, 287, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Xi, Chen.; Gentao, L.; Peixin, Y.; Xi, C.; Jianxiang, Z.; Daoyin, D.; Peixin, Y. Endoplasmic reticulum stress-induced CHOP inhibits PGC-1α and causes mitochondroial disfunction in diabetic embryopathy. Toxicol. Sci. 2017. [Google Scholar] [CrossRef]
- Seiji, T.; Shigetoshi, Y.; Motohiro, M.; Jun-ichi, K.; Masataka, M.; Tomomi, G. CHOP is involved in neuronal apoptosis induced by neurotrophic factor deprivation. FEBS Lett. 2006, 580, 3462–3468. [Google Scholar]
- Yang, S.H.; Sharrocks, A.D.; Whitmarsh, A.J. MAP kinase signalling cascades and transcriptional regulation. Gene 2013, 513, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Chen, R.; Li, H. Cross-talk between PI3K/Akt and MEK/ERK pathways mediates endoplasmic reticulum stress-induced cell cycle progression and cell death in human hepatocellular carcinoma cells. Int. J. Oncol. 2009, 34, 1749–1757. [Google Scholar] [PubMed]
- Samatar, A.A.; Poulikakos, P.I. Targeting RAS-ERK signalling in cancer: Promises and challenges. Nat. Rev. Drug Discov. 2014, 13, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Boelens, J.; Lust, S.; Offner, F.; Bracke, M.E.; Vanhoecke, B.W. The endoplasmic reticulum: a target for new anticancer drugs. In Vivo 2007, 21, 215–226. [Google Scholar] [PubMed]
- Sheldrick, G.M. A short history of SHELX. Acta Crystallogr. 2008, 64, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, G.A.; Seto, J.; Patil, S.; Nudelman, G.; Sealfon, S.C. Getting started in biological pathway construction and analysis. PLoS Comput. Biol. 2008, 4, e16. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors on request. |






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siyo, V.; Schäfer, G.; Hunter, R.; Grafov, A.; Grafova, I.; Nieger, M.; Katz, A.A.; Parker, M.I.; Kaschula, C.H. The Cytotoxicity of the Ajoene Analogue BisPMB in WHCO1 Oesophageal Cancer Cells Is Mediated by CHOP/GADD153. Molecules 2017, 22, 892. https://doi.org/10.3390/molecules22060892
Siyo V, Schäfer G, Hunter R, Grafov A, Grafova I, Nieger M, Katz AA, Parker MI, Kaschula CH. The Cytotoxicity of the Ajoene Analogue BisPMB in WHCO1 Oesophageal Cancer Cells Is Mediated by CHOP/GADD153. Molecules. 2017; 22(6):892. https://doi.org/10.3390/molecules22060892
Chicago/Turabian StyleSiyo, Vuyolwethu, Georgia Schäfer, Roger Hunter, Andriy Grafov, Iryna Grafova, Martin Nieger, Arieh A. Katz, M. Iqbal Parker, and Catherine H. Kaschula. 2017. "The Cytotoxicity of the Ajoene Analogue BisPMB in WHCO1 Oesophageal Cancer Cells Is Mediated by CHOP/GADD153" Molecules 22, no. 6: 892. https://doi.org/10.3390/molecules22060892
APA StyleSiyo, V., Schäfer, G., Hunter, R., Grafov, A., Grafova, I., Nieger, M., Katz, A. A., Parker, M. I., & Kaschula, C. H. (2017). The Cytotoxicity of the Ajoene Analogue BisPMB in WHCO1 Oesophageal Cancer Cells Is Mediated by CHOP/GADD153. Molecules, 22(6), 892. https://doi.org/10.3390/molecules22060892