In Vitro Evaluation of Sub-Lethal Concentrations of Plant-Derived Antifungal Compounds on FUSARIA Growth and Mycotoxin Production

 ,
,  and
and
Abstract
:1. Introduction
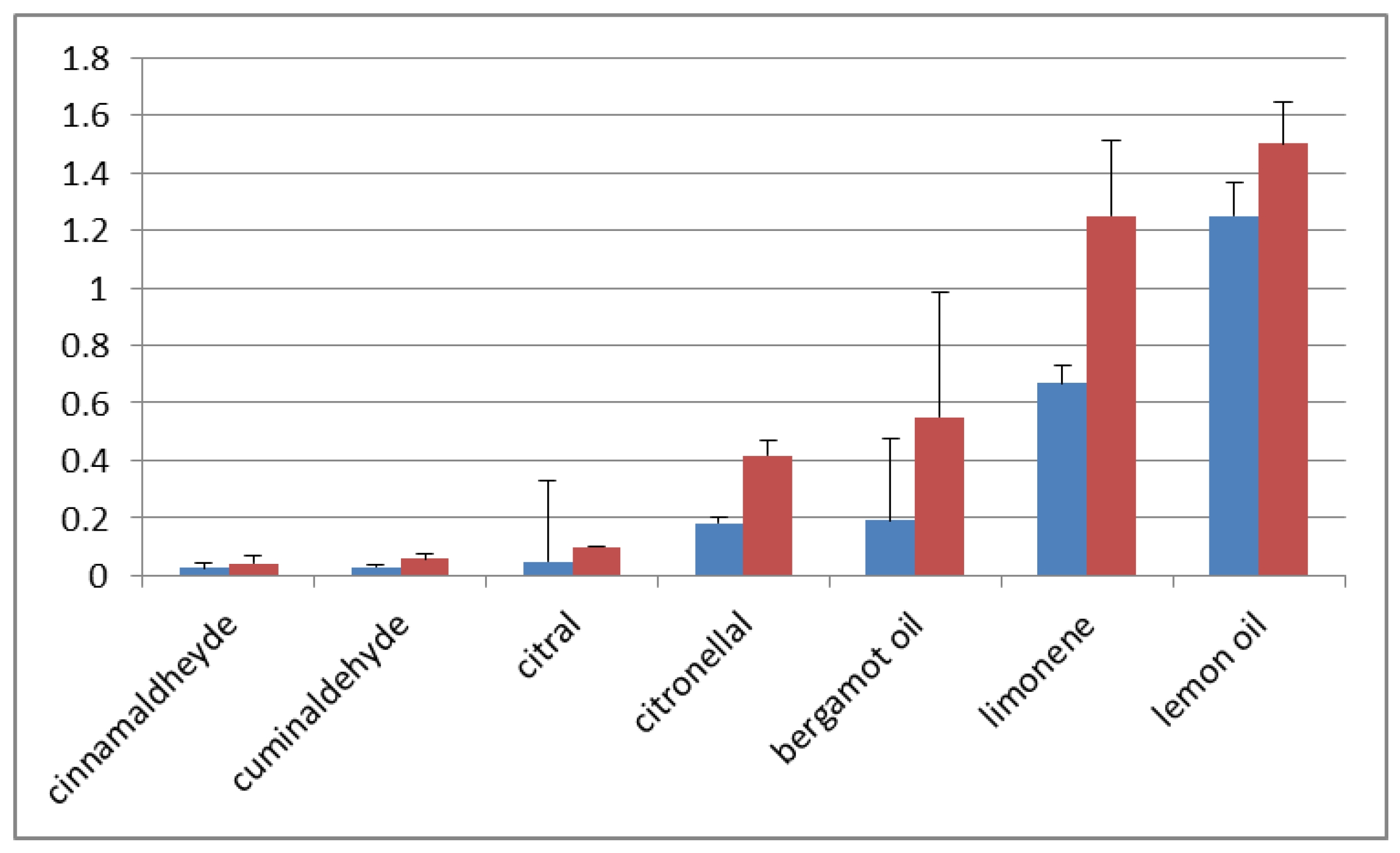
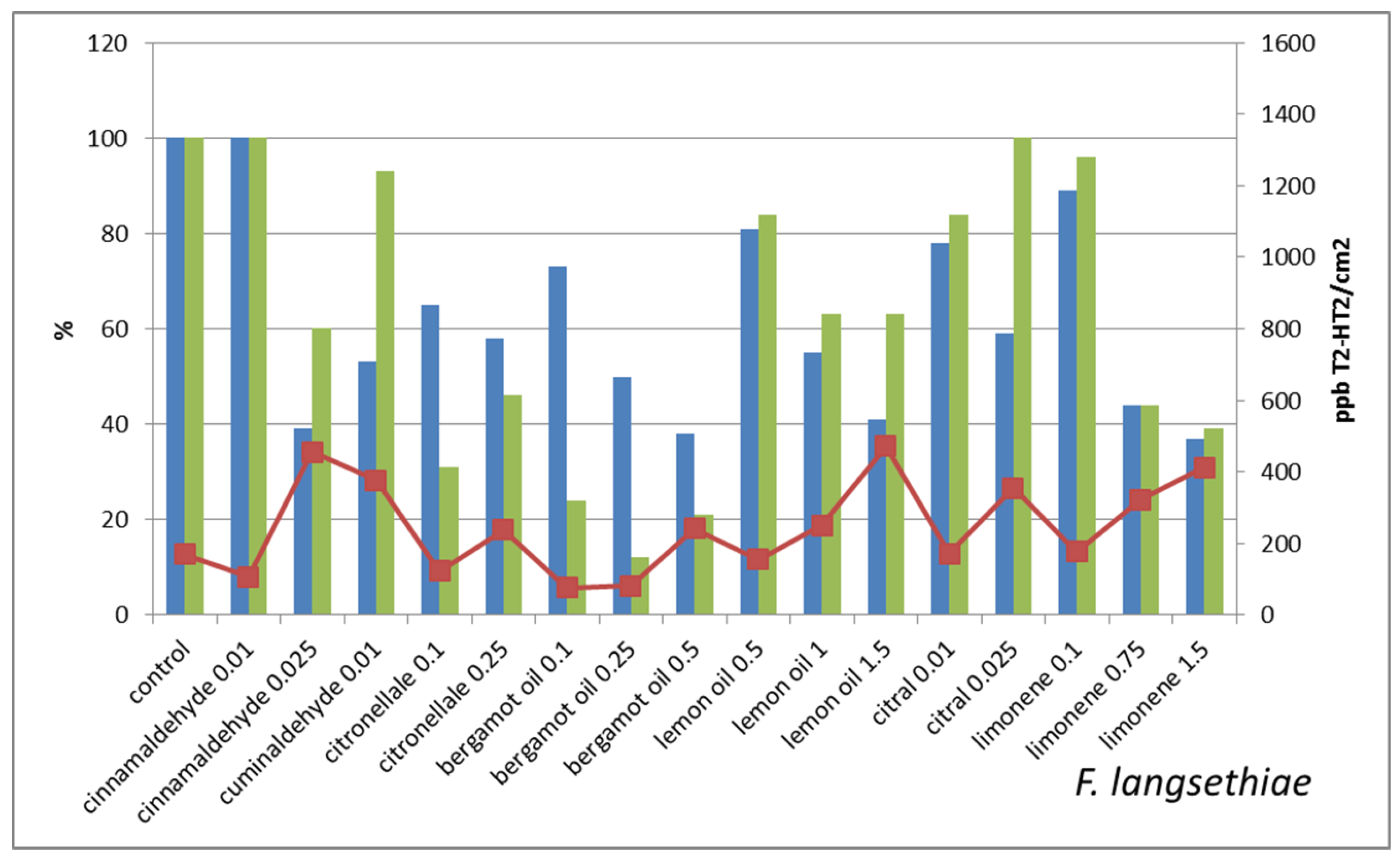
2. Results and Discussion
- the mycotoxin concentration/cm2 in treated is lower than in untreated control;
- the mycotoxin concentration/cm2 in treated is very close to untreated control;
- the mycotoxin concentration/cm2 is two–three times higher in treated in comparison with untreated control;
- the mycotoxin concentration/cm2 in treated is orders of magnitude higher than in untreated control
3. Material and Methods
3.1. Fungal Strains
3.2. Chemicals
3.3. Evaluation of the Effect of Compounds on In Vitro Fungal Growth
3.4. Evaluation of the Effect of Compounds on In Vitro Mycotoxin Production
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium Head Blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in north America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Path. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- De Ruyck, K.; de Boevre, M.; Huybrechts, I.; De Saeger, S. Dietary mycotoxins, co-exposure, and carcinogenesis in humans: Short review. Mut. Res./Rev. Mut. Res. 2015, 766, 32–41. [Google Scholar] [CrossRef]
- Foroud, N.A.; Eudes, F. Trichothecenes in cereal grains. Int. J. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Hohn, T.M.; McCormick, S.P. Trichothecene biosynthesis in Fusarium. Microbiol. Mol. Biol. Rev. 1993, 57, 595–604. [Google Scholar]
- Terzi, V.; Tumino, G.; Stanca, A.M.; Morcia, C. Reducing the incidence of cereal head infection and mycotoxins in small grain cereal species. J. Cereal Sci. 2014, 59, 284–293. [Google Scholar] [CrossRef]
- Mesterhazy, Á. Control of Fusarium Head Blight of wheat by fungicides. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2003; pp. 363–380. [Google Scholar]
- Lang, G.; Buchbauer, G. A review on recent research results (2008–2010) on essential oils as antimicrobials and antifungals. Flavour Fragr. J. 2012, 27, 13–39. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods: A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Morcia, C.; Spini, M.; Malnati, M.; Stanca, A.M.; Terzi, V. Essential oils and their components for the control of phytopathogenic fungi that affect plant health and agri-food quality and safety. In Natural Antimicrobials for Food Safety and Food Quality; Rai, M., Chikindas, M., Eds.; CABI Press: Oxford, UK, 2011; pp. 224–241. [Google Scholar]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorg. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B.; Machial, C.M. Pesticides based on plant essential oils: from traditional practice to commercialization. In Naturally Occurring Bioactive Compounds; Rai, M., Carpinella, M.C., Eds.; Elsevier BV: Amsterdam, the Netherland, 2016; pp. 29–44. [Google Scholar]
- Prakash, B.; Kedia, A.; Mishra, P.K.; Dubey, N.K. Plant essential oils as food preservatives to control moulds, mycotoxin contamination and oxidative deterioration of agri-food commodities–Potential and challenges. Food Contr. 2015, 47, 381–391. [Google Scholar] [CrossRef]
- Avila-Sosa, R.; Navarro-Cruz, A.R.; Sosa-Morales, M.E.; Lopez-Malo, A.; Palou, E. Bergamot (Citrus bergamia) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V.R., Ed.; Academic Press: London, UK, 2016; pp. 247–252. [Google Scholar]
- Dugo, G.; Bonaccorsi, I. Citrus Bergamia: Bergamot and Its Derivatives; CCR Press: Boka Raton, FL, USA, 2013. [Google Scholar]
- Stević, T.; Berić, T.; Šavikin, K.; Soković, M.; Gođevac, D.; Dimkić, I.; Stanković, S. Antifungal activity of selected essential oils against fungi isolated from medicinal plant. Ind. Crops Prod. 2014, 55, 116–122. [Google Scholar] [CrossRef]
- Jing, L.; Lei, Z.; Li, L.; Xie, R.; Xi, W.; Guan, Y.; Zhou, Z. Antifungal activity of citrus essential oils. J. Agric. Food Chem. 2014, 62, 3011–3033. [Google Scholar] [CrossRef] [PubMed]
- Calo, J.R.; Crandall, P.G.; O’Bryan, C.A.; Ricke, S.C. Essential oils as antimicrobials in food systems–A review. Food Cont. 2015, 54, 111–119. [Google Scholar] [CrossRef]
- Ramezani, H.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Dargan, J.S. Fungicidal effect of volatile oils from Eucalyptus citriodora and its major constituent citronellal. New Zealand Plant Prot. 2002, 55, 327–330. [Google Scholar]
- Wu, Y.; OuYang, Q.; Tao, N. Plasma Membrane Damage Contributes to Antifungal Activity of Citronellal Against Penicillium Digitatum. J. Food Sci. Technol. 2016, 53, 3853–3858. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-Y.; Wu, X.-M.; Yin, X.-H.; Long, Y.-H.; Li, M. Naturally produced citral can significantly inhibit normal physiology and induce cytotoxicity on Magnaporthe grisea. Pest. Biochem. Physiol. 2015, 118, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Hua, H.; Selvaraj, J.N.; Zhao, Y.; Zhou, L.; Liu, X.; Liu, Y. Growth inhibition and morphological alterations of Fusarium verticillioides by cinnamon oil and cinnamaldehyde. Food Cont. 2014, 46, 343–350. [Google Scholar] [CrossRef]
- Sun, Q.; Shang, B.; Wang, L.; Lu, Z.; Liu, Y. Cinnamaldehyde inhibits fungal growth and aflatoxin B1 biosynthesis by modulating the oxidative stress response of Aspergillus flavus. Appl. Microbiol. Biotechnol. 2016, 100, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Marei, G.I.K.; Rasoul, M.A.A.; Abdelgaleil, S.A. Comparative antifungal activities and biochemical effects of monoterpenes on plant pathogenic fungi. Pest. Biochem. Physiol. 2012, 103, 56–61. [Google Scholar] [CrossRef]
- Adegoke, G.O.; Evwiehuroma, F.O.; Afolabi, M.O. African cardamom (Aframomum danielli) oils. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V.R., Ed.; Academic Press: London, UK, 2016; pp. 163–171. [Google Scholar]
- Deising, H.B.; Reimann, S.; Pascholati, S.F. Mechanisms and significance of fungicide resistance. Braz. J. Microbiol. 2008, 39, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, N.; Tokai, T.; Takahashi-Ando, N.; Fujimura, M.; Kimura, M. Genetically engineered Fusarium as a tool to evaluate the effects of environmental factors on initiation of trichothecenes biosynthesis. FEMS Microbiol. Lett. 2007, 275, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Audenart, K.; Callewaert, E.; Höfte, M.; De saeger, S.; Gaesaert, G. Hydrogen peroxide induced by the fungicide prothioconazole triggers deoxynivalenol (DON) production by Fusarium graminearum. BMC Microbiol. 2010, 10, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popiel, D.; Dawidziuk, A.; Koczyk, G.; Mackowiak, A. Multiple factes of response to fungicides-the influence of azole treatment on expression of key mycotoxin biosynthetic genes and candidate resistance factors in the control of resistant Fusarium strains. Eur. J. Plant Pathol. 2017, 147, 773–785. [Google Scholar] [CrossRef]
- Kurita, N.; Koike, S. Synergistic antimicrobial effect of ethanol, sodium chloride, acetic acid and essential oil components. Agric. Biol. Chem. 1983, 47, 67–75. [Google Scholar]
- Morcia, C.; Malnati, M.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Add. Contam. 2012, 3, 415–422. [Google Scholar]
- Lu, M.; Han, Z.; Yao, L. In vitro and in vivo antimicrobial efficacy of essential oils and individual compounds against Phytophthora parasitica var. nicotianae. J. Appl. Microbiol. 2013, 115, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, J.; Shin, S.C.; Lee, S.G.; Park, I.K. Antifungal activity of Myrtaceae essential oils and their components against three phytopathogenic fungi. Flavour Frag. J. 2008, 23, 23–28. [Google Scholar] [CrossRef]
- Edwards, S.G.; Pirgozliev, S.R.; Hare, M.C.; Jenkinson, P. Quantification of trichothecene-producing Fusarium species in harvested grain by competitive PCR to determine efficacies of fungicides against Fusarium Head Blight of winter wheat. Appl. Environ. Microbiol. 2001, 67, 1575–1580. [Google Scholar]
- Hasan, H.A.H. Fungicide inhibition of aflatoxins, diacetoxyscirpenol and zearalenone production. Folia Microbiol. 1993, 38, 295–298. [Google Scholar] [CrossRef]
- Matthies, A.; Buchenauer, H. Investigations on the action of different active ingredients on the biosynthesis of mycotoxins in Fusarium culmorum and Fusarium graminearum. In Modern Fungicides and Antifungal Compounds; Lyr, H., Russell, P.E., Sisler, H.D., Eds.; Intercept Ltd.: Andover, UK, 1996; pp. 199–204. [Google Scholar]
- D’Mello, J.P.F.; Macdonald, A.M.C.; Postel, D.; Dijksma, W.T.P.; Dujardin, A.; Placinta, C.M. Pesticide use and mycotoxin production in Fusarium and Aspergillus phytopathogens. Eur. J. Plant Pathol. 1998, 104, 741–751. [Google Scholar] [CrossRef]
- Moss, M.O.; Frank, J.M. Influence of the fungicide tridemorph on T2 toxin production of Fusarium sporotrichioides. Trans. Br. Mycol. Soc. 1985, 84, 585–590. [Google Scholar] [CrossRef]
- Placinta, C.M.; Macdonald, A.M.C.; D’Mello, J.B.F.; Harling, R. The Influence of Carbendazim on Mycotoxin Production in Fusarium sporotrichioides. In Proceedings of the Brighton Crop Protection Conference 1996: Pests and Diseases, Brighton, UK, 18–21 November 1996; British Crop Protection Council: Farnham, UK, 1996; pp. 415–416. [Google Scholar]
- Magan, N.; Hope, R.; Colleate, A.; Baxter, E.S. Relationship between Growth and Mycotoxin Production by Fusarium species, Biocides and Environment. Eur. J. Plant Pathol. 2002, 108, 685–690. [Google Scholar] [CrossRef]
- Kulik, T.; Loiko, M.; Jestoi, M.; Perkowski, J. Sublethal concentrations of azoles induce tri transcript levels and trichothecene production in Fusarium graminearum. FEMS Microbiol. Lett. 2012, 335, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz Cabral, L.; Fernández Pinto, V.; Patriarca, A. Application of plant derived compounds to control fungal spoilage and mycotoxin production in foods. Int. J. Food Microbiol. 2013, 166, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sumalan, R.-M.; Alexa, E.; Poiana, M.-A. Assessment of inhibitory potential of essential oils on natural mycoflora and Fusarium mycotoxins production in wheat. Chem. Central J. 2013, 7, 32. [Google Scholar] [CrossRef] [PubMed]
- Hope, R.; Cairns-Fuller, V.; Aldred, D.; Magan, N. Use of antioxidants and essential oils for controlling mycotoxins in grain. BCPC Crop Sci. Technol. 2005, 5B, 429–436. [Google Scholar]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marí, S. Effect of Equisetum arvense and Stevia rebaudiana extracts on growth and mycotoxin production by Aspergillus flavus and Fusarium verticillioides in maize seeds as affected by water activity. Int. J. Food Microbiol. 2012, 153, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Ricelli, A.; Zjalic, S.; Fabbri, A.A.; Fanelli, C. Natural function of mycotoxins and control of their biosynthesis in fungi. Appl. Microbiol. Biotechnol. 2010, 87, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Ferruz, E.; Atanasova-Pénichon, V.; Bonnin-Verdal, M.; Marchegay, G.; Pinson-Gadais, L.; Ducos, C.; Lorán, S.; Ariño, A.; Barreau, C.; Richard-Forget, F. Effects of Phenolic Acids on the Growth and Production of T-2 and HT-2 Toxins by Fusarium langsethiae and F. sporotrichioides. Molecules 2016, 21, 449. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Probit Analysis; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Crop Prospects and Food Situation. Available online: http://www.fao.org/3/a-i6903e.pdf (accessed on 28 July 2017).
Sample Availability: Samples of the fungal isolates are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fusarium sporotrichioides | Fusarium graminearum | Fusarium langsethiae | ||||
|---|---|---|---|---|---|---|
| EC50 | EC100 | EC50 | EC100 | EC50 | EC100 | |
| cuminaldehyde | 0.031 | 0.075 | 0.046 | 0.075 | 0.015 | 0.025 |
| cinnamaldehyde | 0.036 | 0.05 | 0.02 | 0.025 | 0.02 | 0.05 |
| lemon oil | 1.5 | >1.5 | 1.3 | >1.5 | 0.95 | >1.5 |
| citral | 0.067 | 0.15 | 0.05 | 0.1 | 0.027 | 0.05 |
| limonene | 1 | >1.5 | 0.5 | 0.75 | 0.5 | >1.5 |
| bergamot oil | 0.12 | 0.25 | 0.22 | 0.7 | 0.23 | 0.7 |
| citronellal | 0.32 | 0.5 | 0.12 | 0.25 | 0.11 | 0.5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morcia, C.; Tumino, G.; Ghizzoni, R.; Bara, A.; Salhi, N.; Terzi, V. In Vitro Evaluation of Sub-Lethal Concentrations of Plant-Derived Antifungal Compounds on FUSARIA Growth and Mycotoxin Production. Molecules 2017, 22, 1271. https://doi.org/10.3390/molecules22081271
Morcia C, Tumino G, Ghizzoni R, Bara A, Salhi N, Terzi V. In Vitro Evaluation of Sub-Lethal Concentrations of Plant-Derived Antifungal Compounds on FUSARIA Growth and Mycotoxin Production. Molecules. 2017; 22(8):1271. https://doi.org/10.3390/molecules22081271
Chicago/Turabian StyleMorcia, Caterina, Giorgio Tumino, Roberta Ghizzoni, Assetou Bara, Nesrine Salhi, and Valeria Terzi. 2017. "In Vitro Evaluation of Sub-Lethal Concentrations of Plant-Derived Antifungal Compounds on FUSARIA Growth and Mycotoxin Production" Molecules 22, no. 8: 1271. https://doi.org/10.3390/molecules22081271
APA StyleMorcia, C., Tumino, G., Ghizzoni, R., Bara, A., Salhi, N., & Terzi, V. (2017). In Vitro Evaluation of Sub-Lethal Concentrations of Plant-Derived Antifungal Compounds on FUSARIA Growth and Mycotoxin Production. Molecules, 22(8), 1271. https://doi.org/10.3390/molecules22081271