Composition Analysis by UPLC-PDA-ESI (−)-HRMS and Antioxidant Activity Using Saccharomyces cerevisiae Model of Herbal Teas and Green Teas from Hainan

Abstract
:1. Introduction
2. Results
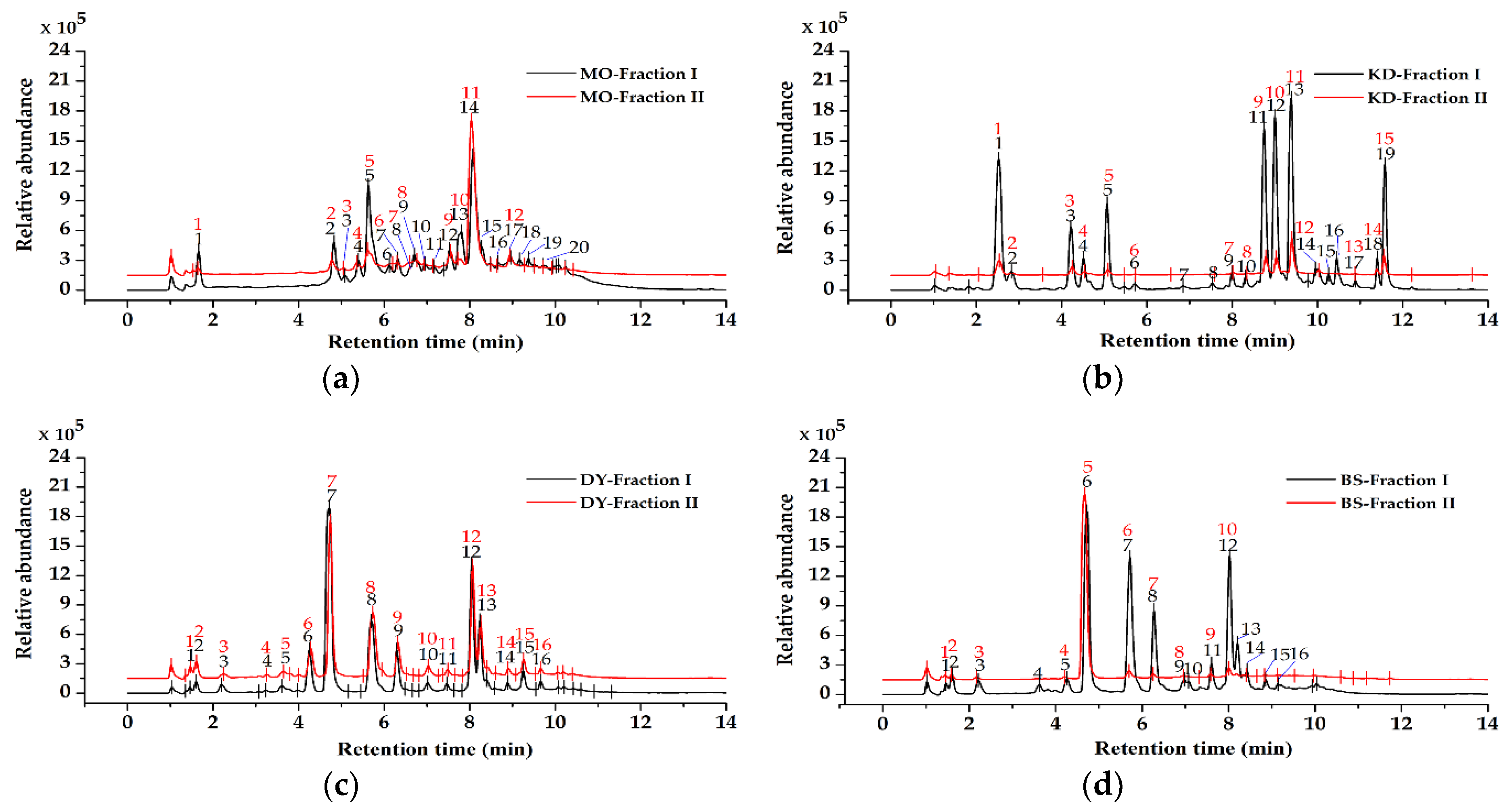
2.1. Results of Phenolic Acids and Flavonoids From the Herbal Teas and Green Teas
2.2. Total Phenolic and Flavonoid Content
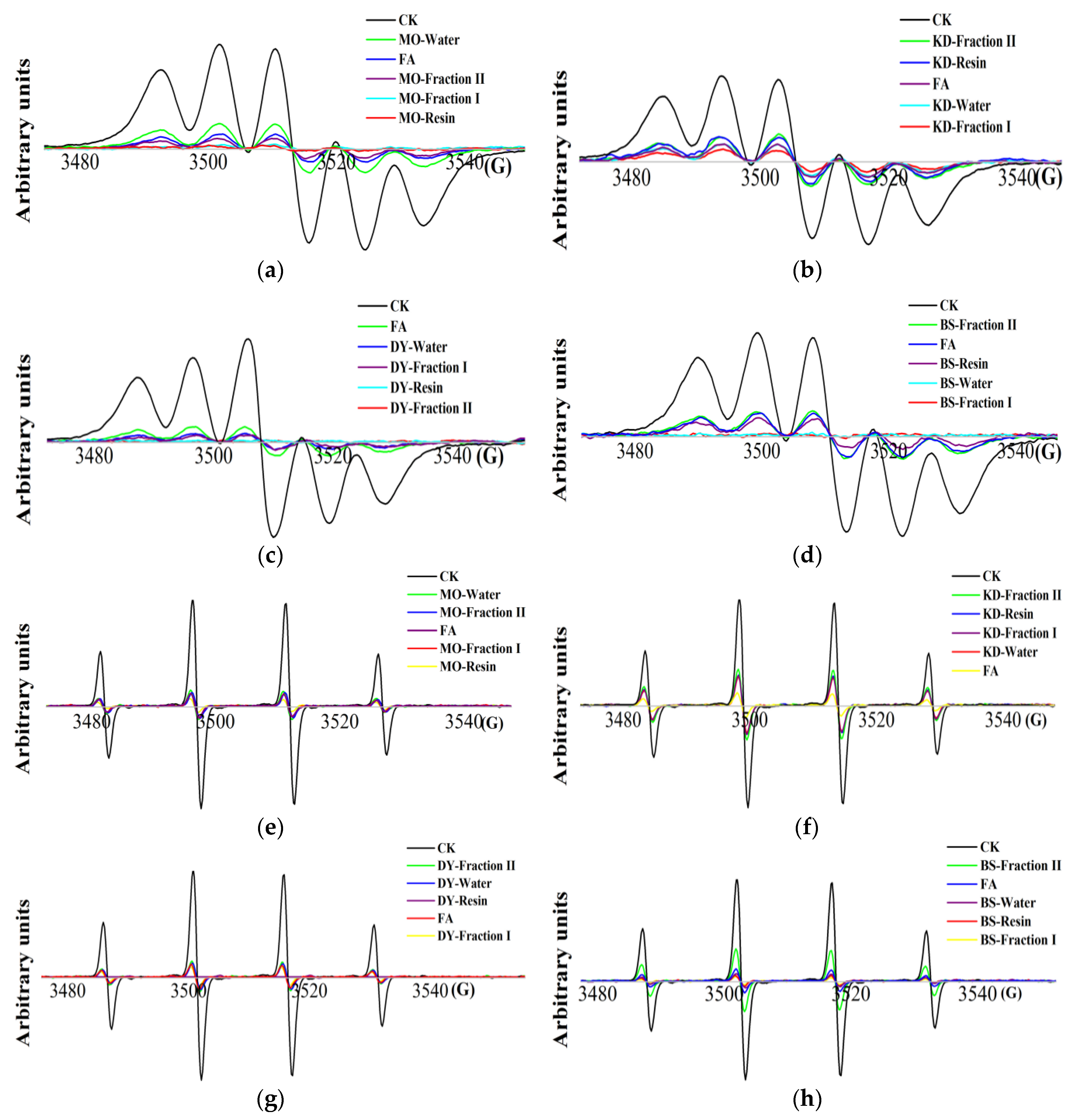
2.3. Detection of DPPH· and HO· Scavenging Activity by EPR Spectroscope
2.4. Protection of S. cerevisiae against Oxidative H2O2 Stress
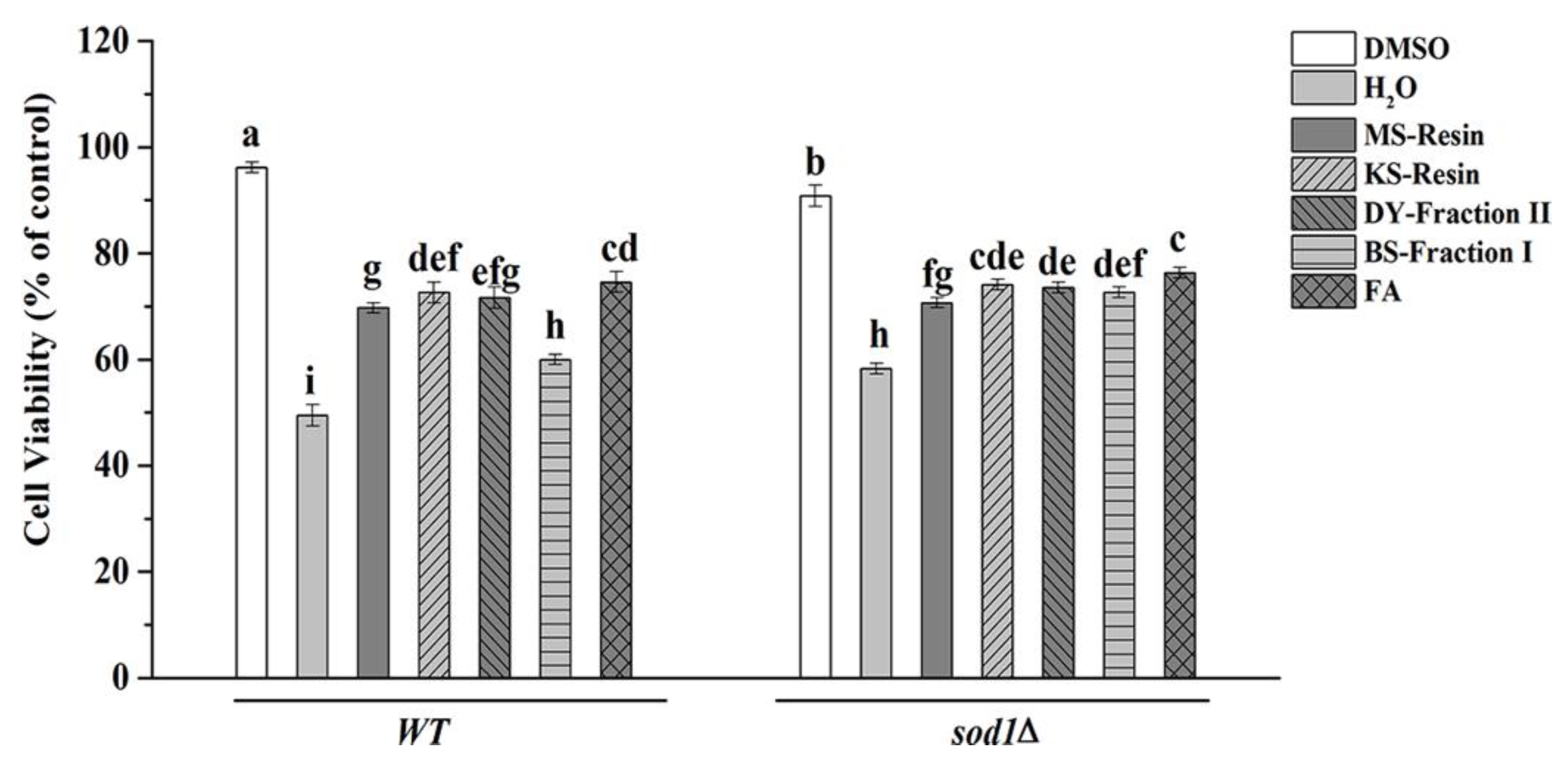
2.4.1. S. cerevisiae Cell Viability
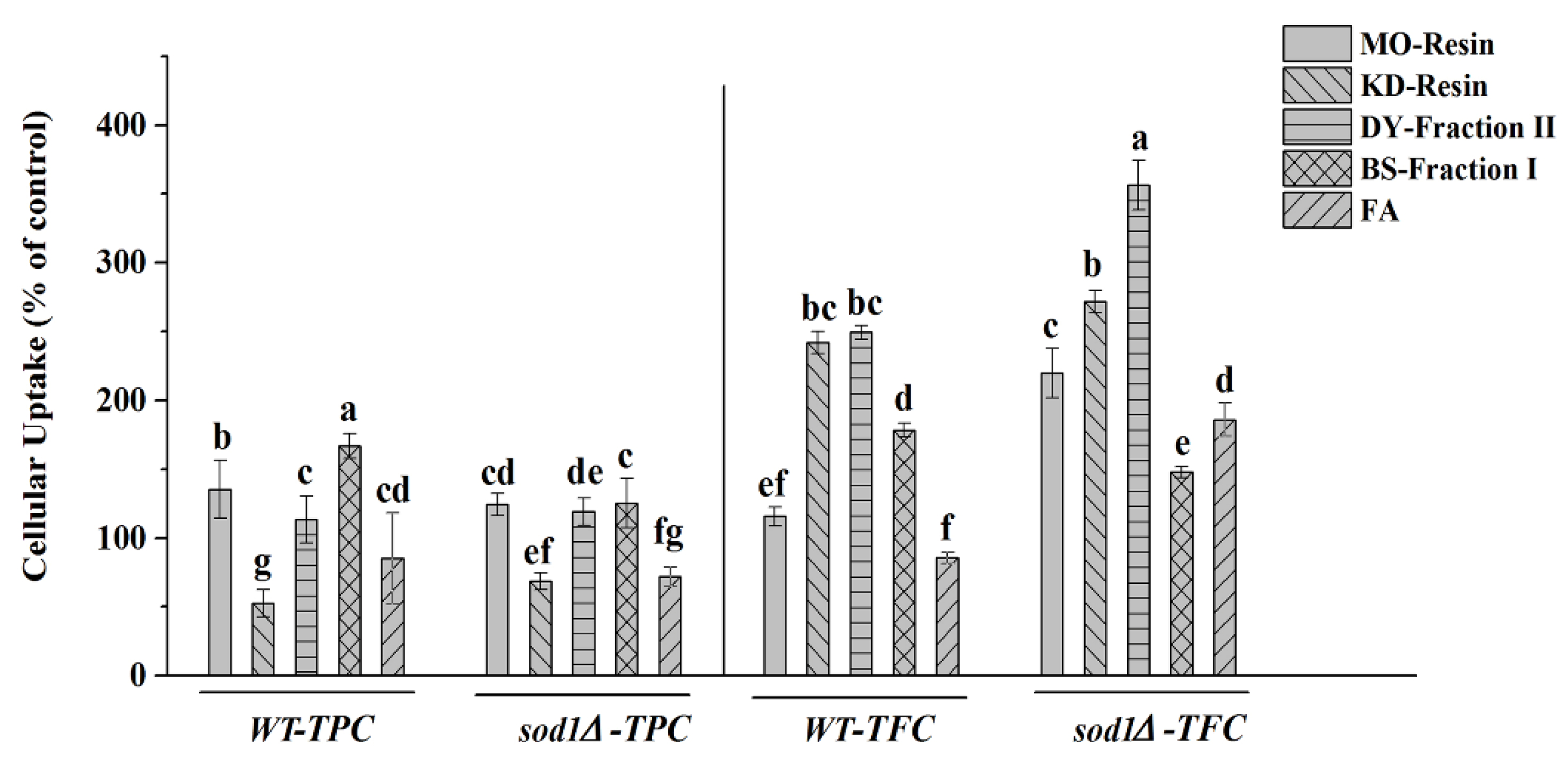
2.4.2. Cellular Uptake of Herbal Teas and Green Teas Extracts
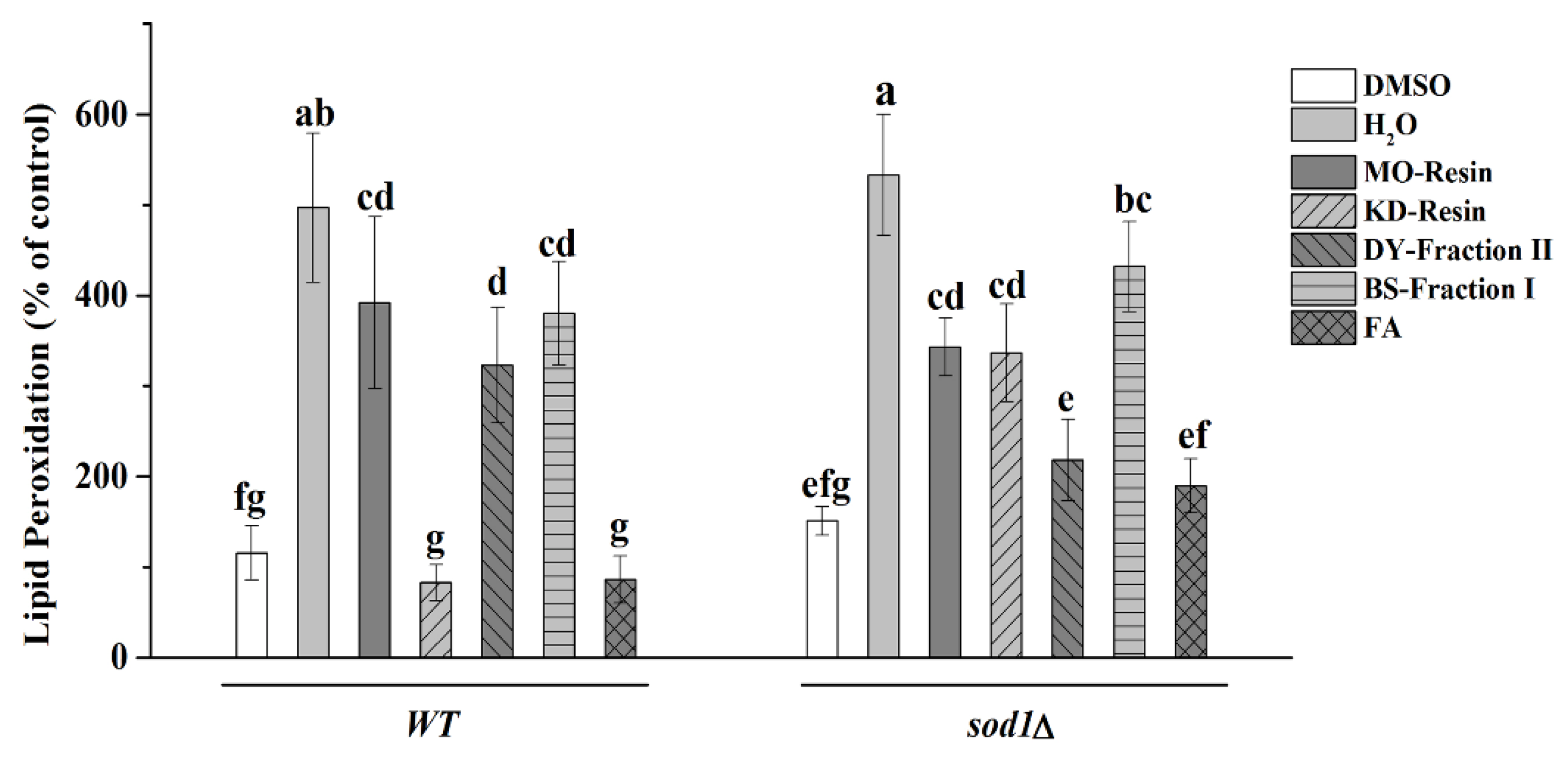
2.4.3. Detection of Lipid Peroxidation
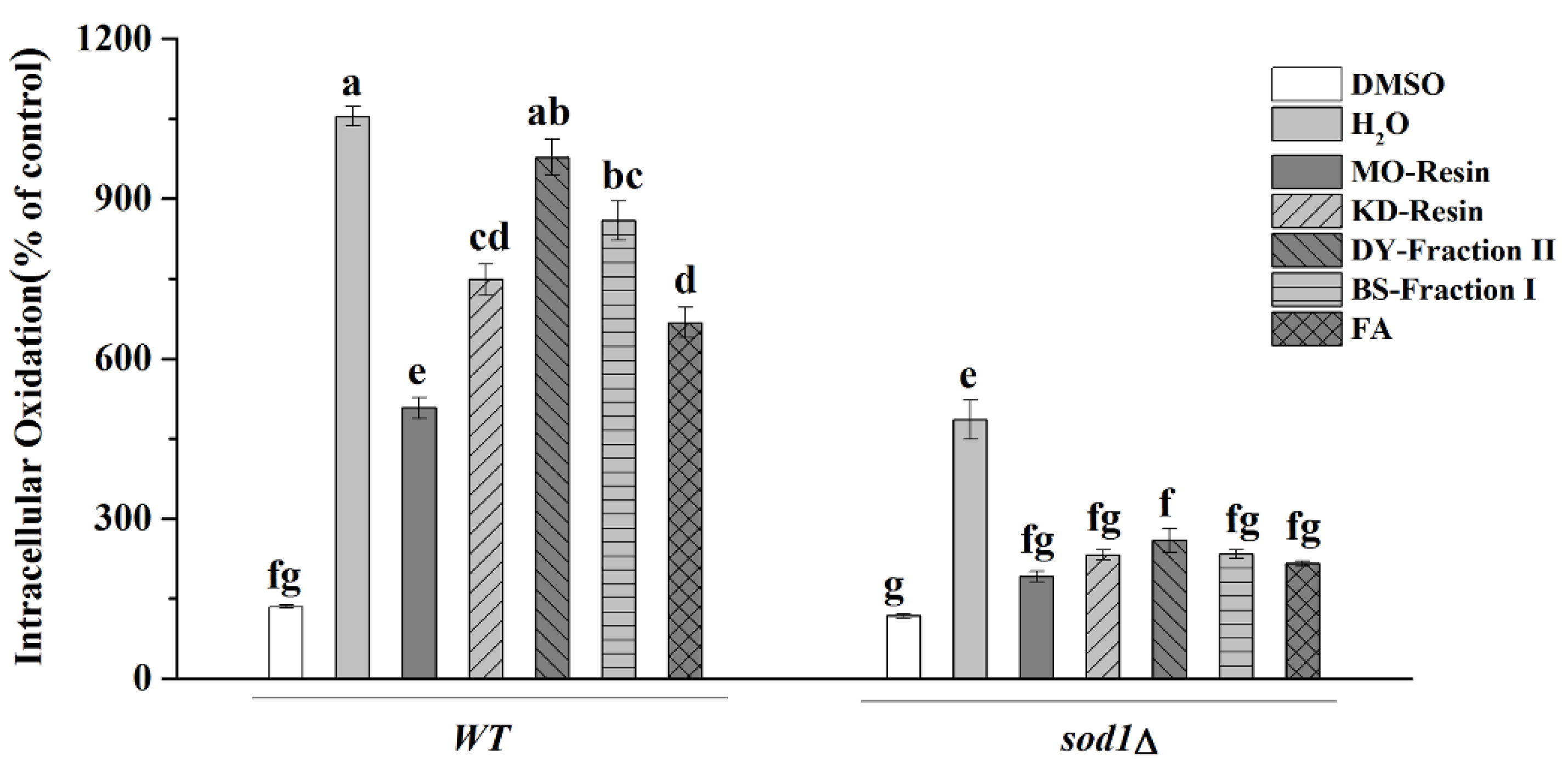
2.4.4. Intracellular Oxidation
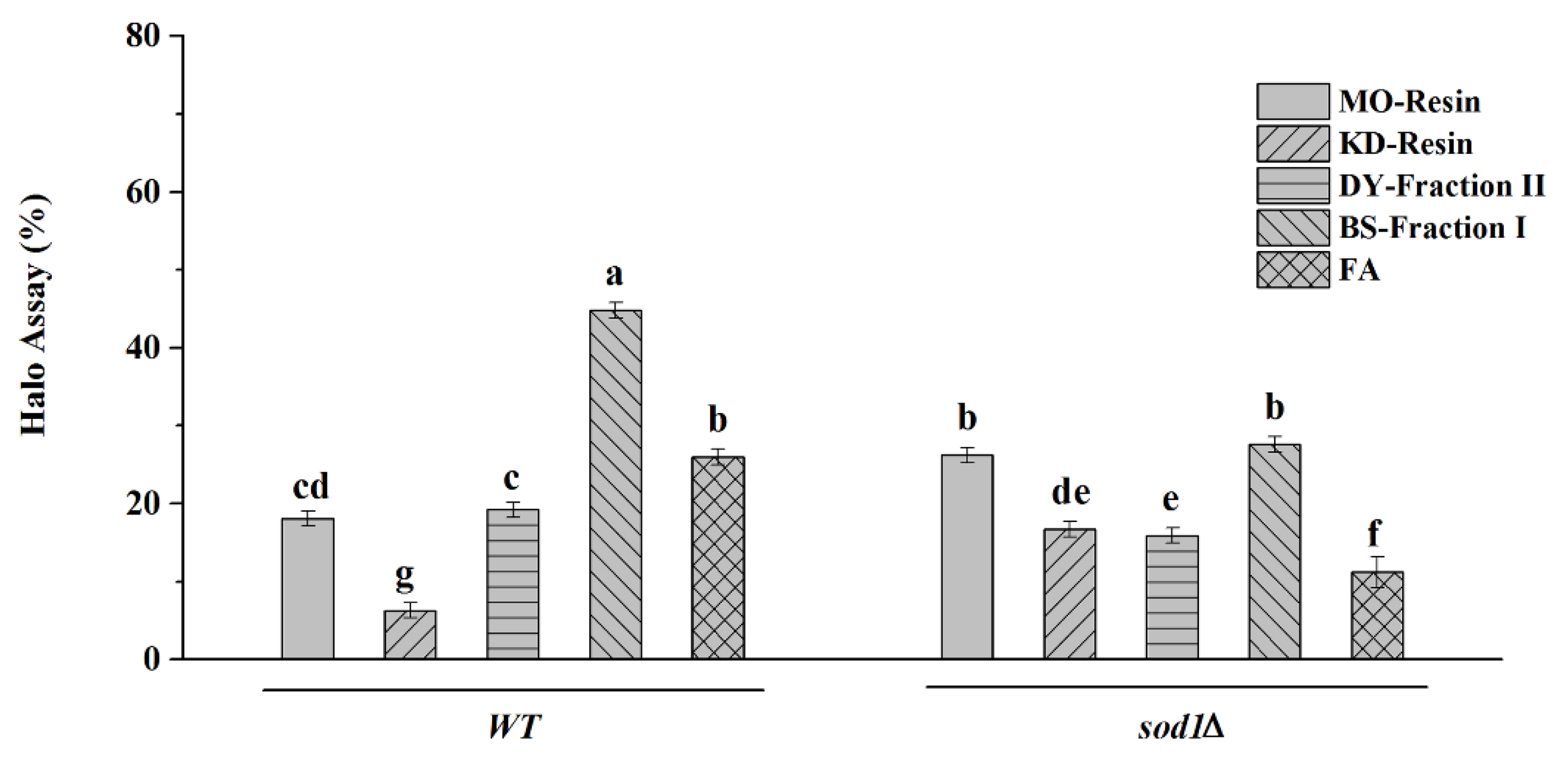
2.4.5. Halo Assay
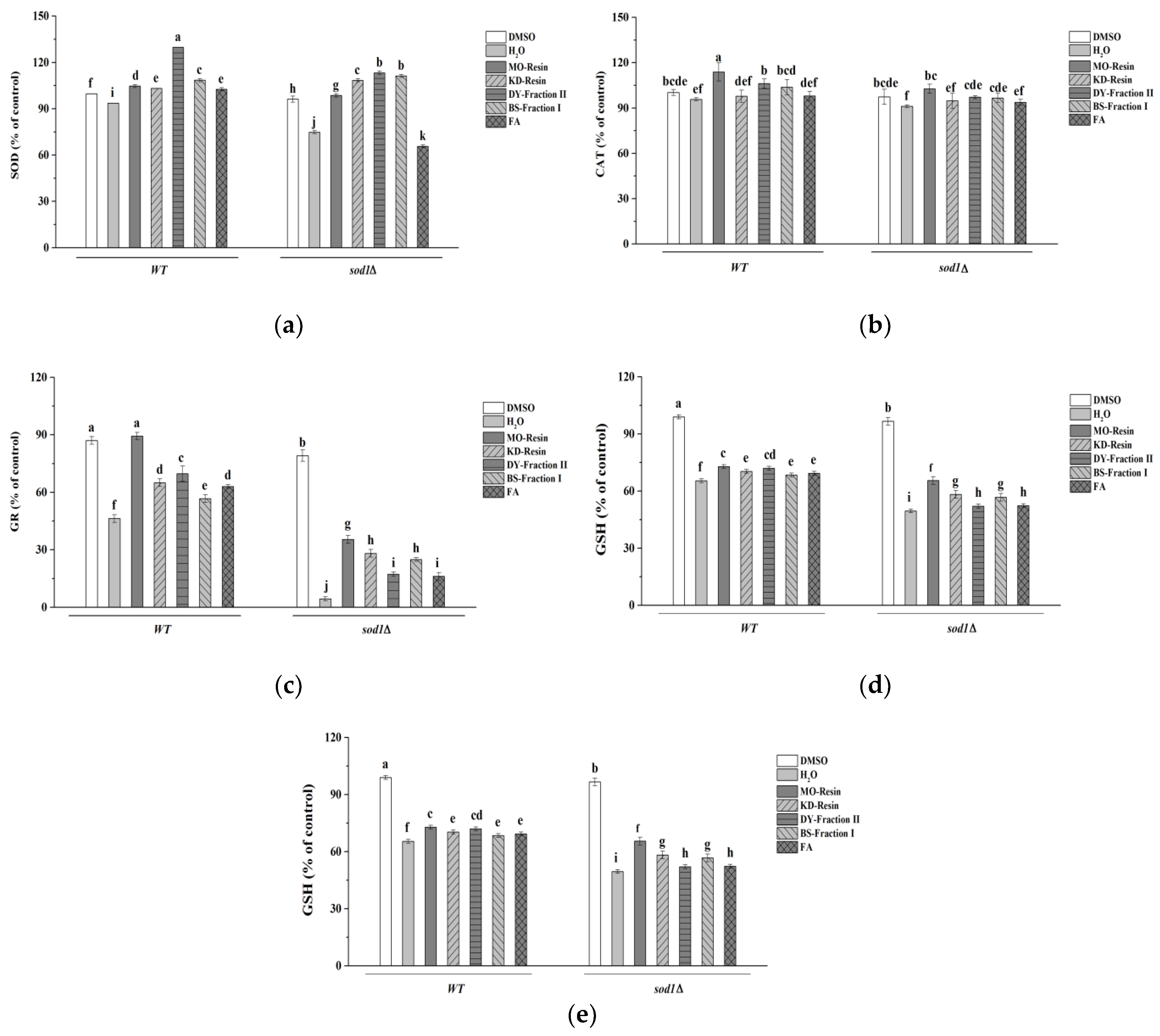
2.4.6. Determination of S. cerevisiae Enzymes Activities
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Chemicals
4.3. Preparation of Herbal Teas and Green Teas Extracts
4.4. Detection of Phenolic Acids and Flavonoids of the Herbal Teas and Green Teas by UPLC-PDA-ESI-(−)-HRMS
4.5. Determination of TPC
4.6. Determination of TFC
4.7. Detection of DPPH· Scavenging Activity by EPR Spectroscopy
4.8. Detection of HO· Scavenging Activity by EPR Spectroscopy
4.9. Protective Effect of Herbal Teas and Green Teas on S. cerevisiae in Oxidative Stress
4.9.1. Determination of S. cerevisiae Cell Viability
4.9.2. Determination of Cellular Uptake by Herbal Teas and Green Teas Extracts
4.9.3. Detection of Lipid Peroxidation
4.9.4. Detection of Intracellular Oxidation
4.9.5. Detection of Halo Assay
4.9.6. Detection of the Enzyme Activity in S. cerevisiae
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MO | Mallotus oblongifolius Muell. Arg. |
| KD | Ilex kudingcha C.J. Tseng |
| DY | Camellia sinensis var. assamica (J. W. Mast.) Kitam. Hainan Dayezhong. |
| BS | Camellia sinensis (L.) O. Ktze. (produced from Hainan Baisha (BS)) |
| TPC | total phenol content |
| TFC | total flavonoids content |
| UPLC-PDA-ESI-(−)-HRMSn | ultra-high pressure liquid chromatography and electrospray ionization coupled with high-resolution mass spectrometry |
| DPPH· | 1-diphenyl-2-picryl-hydrazyl radical |
| HO· | hydroxyl free radical |
| WT | wild-type |
| sod1Δ | isogenic mutant yeast cell, harboring the SOD1 genes knocked out by the KanMX4 gene |
| YPD | yeast extract peptone dextrose |
| H2O2 | hydrogen peroxide |
| DMSO | dimethyl sulfoxide |
| FA | ferulic acid |
| CAT | catalase |
| GR | glutathione reductase |
| SOD | superoxide dismutase |
| GSH | glutathione reduced |
| GSSG | glutathione oxidized |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| ROS | reactive oxygen species |
| O2.− | superoxide anions |
| C | (+)-catechin |
| CG | (−)-catechin gallate |
| EC | (−)-epicatechin |
| EGC | (−)-epigallocatechin |
| ECG | (−)-epicatechin gallate |
| GCG | (−)-gallocatechin gallate |
| EGCG | (−)-epigallocatechin gallate |
| EPR | electron paramagnetic resonance |
| GAE | gallic acid equivalents |
| RE | rutin equivalents |
| DTPA | diethylenetriamine pentaacetic acid |
| DMPO | 5,5-dimethyl-1-proline-N-oxide |
| TCA | trichloroacetic |
| TBA | thiobarbituric acid |
| DCFH-DA | 2′7′-dichlorofluorescein diacetate |
| EDTA | ethylenediaminetetraacetic acid |
| PDA | photo-diode array detector |
| i.d. | Inner diameter |
| LCMS-IT-TOF | liquid chromatography mass spectrometry ion trap time of flight |
References
- Paulsen, C.E.; Carroll, K. S Orchestrating redox signaling networds through regulatory cysteine switches. ACS Chem. Biol. 2010, 5, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B. Thiol chemistry and specificity in redox signaling. Free Radi. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Sign. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H. Schisandrin A prevents oxidative stress-induced DNA damage and apoptosis by attenuating ROS generation in C2C12 cells. Biomed. Pharmacother. 2018, 10, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Zhang, Y.T.; Guo, Y.R.; Huang, Q.L.; Peng, T.; Xu, Y.; Tang, L.; Chen, F. Antioxidant and anti-inflammatory activities of the phenolic extracts of Sapium sebiferum (L.) Roxb. leaves. J. Ethnopharmacol. 2013, 147, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Nayki, C.; Nayki, U.; Gunay, M.; Kulhan, M.; Cankaya, M.; Humeyra Taskin Kafa, A.; Balci, G. Oxidative and antioxidative status in the endometrium of patients with benign gynecological disorders. J. Gynecol. Obstet. Hum. Reprod. 2017, 46, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Chen, J.; Cai, Y.; Lei, Y.; Chen, L.; Pei, L.; Zhou, D.; Liang, X.; Ruan, J. Antioxidant, free radical scavenging, anti-inflammatory and hepatoprotective potential of the extract from Parathelypteris nipponica (Franch. et Sav.) Ching. J. Ethnopharmacol. 2010, 130, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Oroian, M.; Escriche, I. Antioxidants: Characterization, natural sources, extraction and analysis. Food Res. Int. 2015, 74, 10–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peres, R.G.; Tonin, F.G.; Tavares, M.F.M.; Rodriguez-Amaya, D.B. Determination of catechins in green tea infusions by reduced flow micellar electrokinetic chromatography. Food Chem. 2011, 127, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Wang, L.Q.; Wu, H.T.; Lin, L.B.; Yi, X.N. Comparison of anti-atherosclerotic effects of two different extracts from leaves of Mallotus furetianus. Asian. Pac. J. Trop. Med. 2011, 4, 878–882. [Google Scholar] [CrossRef]
- Li, L.; Xu, L.J.; Ma, G.Z.; Dong, Y.M.; Peng, Y.; Xiao, P.G. The large-leaved Kudingcha (Ilex latifolia Thunb and Ilex kudingcha C.J. Tseng): A traditional Chinese tea with plentiful secondary metabolites and potential biological activities. J. Nat. Med. 2013, 67, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Tistaerta, C.; Dejaeghera, B.; Chataignéb, G.; Rivièreb, C.; Nguyen Hoaic, N.; ChauVanc, M.; Quetin-Leclercqb, J.; Vander Heyden, Y. Potential antioxidant compounds in Mallotus species fingerprints. Part II: Fingerprint alignment, data analysis and peak identification. Analytica Chimica Acta 2012, 721, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Phuong, T.T.; Nguyen, D.S.; Tran, M.N.; Tran, M.H.; Nguyen, H.D.; Nguyen, D.T.; KiHwan, B.; Won, K.O. Antioxidant activity and principles of Vietnam bitter tea Ilex kudingcha. Food Chem. 2009, 113, 139–145. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Huang, W.Y.; Gu, M.T.; Ji, Y.Y.; Zhang, W.M. Antioxidant activity assay and HPLC-MS analysis of polyphenols constituents of baisha green tea. Chin. J. Trop. Crops. 2016, 37, 829–834. [Google Scholar] [CrossRef]
- Gu, X.L.; Li, D.M. Organic planting techniques of Hainan Dayezhong tea plantations. J. Agric. Tech. 2017, 37, 106–108. [Google Scholar] [CrossRef]
- Ellnain-Wojtaszek, M.; Kruczyński, Z.; Kasprzak, J. Investigation of the free radical scavenging activity of Ginkgo biloba L. leaves. Fitoterapia 2003, 7, 1–6. [Google Scholar] [CrossRef]
- Polovka, M.; Brezová, V.; Staško, A. Antioxidant properties of tea investigated by EPR spectroscopy. Biophys. Chem. 2003, 106, 39–56. [Google Scholar] [CrossRef]
- Nurul, A.M.A.; Sara, P.; Lluís, F.; Luis, J.; Maria, P.A. Radical Scavenging of White Tea and Its Flavonoid Constituents by Electron Paramagnetic Resonance (EPR) Spectroscopy. J. Agric. Food Chem. 2014, 62, 5743–5748. [Google Scholar] [CrossRef]
- Kim, J.H.; Campbell, B.C.; Yu, J.; Mahoney, N.; Chan, K.L.; Molyneux, R.J.; Bhatnagar, D.; Cleveland, T.E. Examination of fungal stress response genes using Saccharomyces cerevisiae as a model system: Targeting genes affecting aflatoxin biosynthesis by Aspergillus flavus. Appl. Microbiol. Biotech. 2005, 67, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Mager, W.H.; Winderickx, J. Yeast as a model for medical and medicinal research. Trends Pharmacol. Sci. 2005, 26, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Dani, C.; Bonatto, D.; Salavador, M.; Pereira, M.D.; Henriques, J.A.; Eleutherio, E. Antioxidant protection of resveratrol and catechin in Saccharomyces cerevisiae. J. Agric. Food Chem. 2008, 56, 4268–4272. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Li, W.; Koike, K.; Liu, L.; Fu, X.; Lin, L.; Chen, Y.; Nikaido, T. Two new galloylglucosides from the leaves of Mallotus furetianus. Chem. Pharm. Bull. 2004, 52, 776–779. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Kuang, G.; Chen, X.H.; Zeng, R. Identification of chemical composition of leaves and flowers from paeonia rockii by UHPLC-Q-exactive orbitrap HRMS. Molecules 2016, 21, 947. [Google Scholar] [CrossRef] [PubMed]
- Su, B.K.; Hideyuki, M.; Kosaku, T.; Yamasaki, M.; Yamato, O.; Maede, Y.; Katakura, K.; Kobayashi, S.; Trimurningsih, T.; Chairul, C.; et al. Anti-babesial and Anti-plasmodial compounds from Phyllanthus niruri. J. Nat. Prod. 2005, 68, 537–539. [Google Scholar] [CrossRef]
- Lin, L.B.; Fu, X.W.; Ai, C.H.; Shen, J.; Wei, K.; Li, W. Studies on chemical constituents in leaves of Mallotus furetianus I. China J. Chin. Materia Medica 2006, 31, 0477–0480. [Google Scholar]
- Chen, D.L.; Zheng, W.; Feng, J.; Liu, Y.Y. Chemical constituents from Mallotus furetianus. Chin. Tradit. Herbal Drugs 2017, 48, 4851–4855. [Google Scholar] [CrossRef]
- Wu, C.Y.; Xu, H.R.; Julien, H. Determination of catechins and flavonol glycosides in Chinese tea varieties. Food Chem. 2012, 132, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Kim, S.Y.; Lee, S.J.; Han, G.P.; Han, J.S.; Lee, K.R.; Kozukue, N. Distribution of Catechins, Theaflavins, Caffeine, and Theobromine in 77 Teas Consumed in the United States. J. Food Sci. 2005, 70, 550–559. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Shimada, H.; Saka, M.; Yoshizumi, S.; Yamahara, J.; Matsuda, H. Medicinal food stuffs. V. Moroheiya. (1): Absolute stereostructures of corchoionosides A, B, and C, histamine release inhibitors from the leaves of Vietnamese Corchorus olitorius L. (Tiliaceae). Chem. Pharm. Bull. (Tokyo) 1997, 45, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Chen, P.; Ozcan, M.; Harnly, J.M. Chromatographic profiles and identification of new phenolic components of ginkgo biloba leaves and selected products. J. Agric. Food Chem. 2008, 56, 6671–6679. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, S.J.; Imsic, M.; Jones, R.; Trenerry, V.C.; Tomkins, B. Characterization of flavonol conjugates in immature leaves of pak choi [Brassica rapa L. Ssp. chinensis L. (Hanelt.)] by HPLC-DAD and LC-MS/MS. J. Agric. Food Chem. 2006, 54, 4855–4860. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yi, H.; Zhao, Z.X.; Shang, X.Y.; Zhu, M.J.; Kuang, G.J.; Zhu, C.C.; Zhang, L. Simultaneous qualitative and quantitative evaluation of Ilex kudingcha C.J. tseng by using UPLC and UHPLC-qTOF-MS/MS. J. Pharm. Biomed. Anal. 2018, 155, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Sandra, C.G.F.; Paula, C.C. Phenolic screening by HPLC–DAD–ESI/MSn and antioxidant capacity of leaves, flowers and berries of rubus grandifolius Lowe. Ind. Crops Prod. 2015, 73, 28–40. [Google Scholar] [CrossRef]
- Tao, W.Q.; Zhou, Z.G.; Zhao, B.; Wei, T.Y. Simultaneous determination of eight catechins and four theaflavins in green, black and oolong tea using new HPLC–MS–MS method. J. Pharm. Biomed. Anal. 2016, 131, 140–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.P.; Ma, R.Y.; Chen, L.; Shi, S.Y.; Cai, P.; Zhang, S.H.; Xiang, H.Y. Antioxidant profiling of vine tea (Ampelopsis grossedentata): Off-line coupling heart-cutting HSCCC with HPLC–DAD–QTOF-MS/MS. Food Chem. 2017, 225, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.Q.; Ma, S.S.; Ren, D.B.; Liu, W.B.; Han, B.S.; Zhang, Y.; Xiao, J.B.; Yi, L.Z.; Deng, B.C. UPLC–Orbitrap–MS/MS combined with chemometrics establishes variations in chemical components in green tea from Yunnan and Hunan origins. Food Chem. 2018, 266, 534–544. [Google Scholar] [CrossRef]
- Jiang, G.B.; Xu, L.; Cao, F.L.; Li, G.Z.; Lei, M. Electron Paramagnetic Resonance (EPR) studies on free radical scavenging capacity of EGB and EGB Cigwerette. Spectra Anal. 2017, 37, 1322–1328. [Google Scholar] [CrossRef]
- Zhou, L.F.; Zhu, X.; Zhao, H.L.; Lin, M.B.; Shi, X.M. Study on antioxidant activities of traditional chinese medicines by electron paramagnetic resonance technique. Nat. Prod. Res. Dev. 2014, 26, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.Q.; Chang, S.K.C.; Zhang, Y. Comparison of a-amylase, α-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.Q.; Chang, S.K.C. Digestive enzyme inhibition activity of the phenolic substances in selected fruits, vegetables and tea as compared to black legumes. J. Funct. Foods 2017, 38, 644–655. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Song, W.O.; Fernandez, M.L.; Bruno, R.S.; Chun, O.K. Development and validation of an algorithm to establish a total antioxidant capacity database of the US diet. Int. J. Food. Sci. Nutr. 2010, 61, 600–623. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.J.; Chang, S.K.C. A comparative study on phenolic profiles and antioxidant activities of legumes as affected by extraction solvents. J. Food Sci. 2007, 72, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.H.; Kim, T.W.; Kim, C.G.; Park, N.H. Antioxidant activity of various solvent fractions from edible brown alga, eisenia bicyclis and its active compounds. J. Food Sci. 2013, 78, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.H.; Huang, K.X.; Yang, X.L.; Xu, H.B. Free radical scavenging and antioxidant activities of flavonoids extracted from the radix of Scutellaria baicalensis Georgi. Biochim. Biophys. Acta 1999, 1472, 643–650. [Google Scholar] [CrossRef]
- Ma, Z.R.; Zhao, B.L.; Yuan, Z.B. Application of electrochemical and spin trapping techniques in the investigation of hydroxyl radicals. Anal. Chim. Acta 1999, 389, 213–218. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Bbiology and Medicine, 3rd ed; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Fernandes, P.N.; Mannarino, S.C.; Silva, C.G.; Pereira, M.D.; Panek, A.D.; Eleutherio, E.C.A. Oxidative stress response in eukaryotes: Effect of glutathione, superoxide dismutase and catalase on adaptation to peroxide and menadione stresses in Saccharomyces cerevisiae. Redox Rep. 2007, 12, 236–244. [Google Scholar] [CrossRef] [PubMed]
- França, M.B.; Panek, A.D.; Eleutherio, E.C. The role of cytoplasmic catalase in dehydration tolerance of saccharomyces cerevisiae. Cell Stress Chaperon 2005, 10, 167–170. [Google Scholar] [CrossRef]
- Cai, Y.Z.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef] [PubMed]
- Ozsoy, N.; Can, A.; Yanardag, R.; Akev, N. Antioxidant activity of Smilax excelsa L. leaf extracts. Food Chem. 2007, 110, 571–583. [Google Scholar] [CrossRef]
- Oprea, E.; Ruta, L.L.; Nicolau, I.; Popa, C.V.; Neagoe, A.D.; Farcasanu, I.C. Vaccinium corymbosum L. (blueberry) extracts exhibit protective action against cadmium toxicity in Saccharomyces cerevisiae cells. Food Chem. 2014, 152, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.M.; Dean, R.T.; Jessup, W. Membrane proteins are critical targets in free radical mediated cytolysis. Biochim. Biophys. Acta 1988, 946, 281–288. [Google Scholar] [CrossRef]
- Atiba, A.S.; Abbiyesuku, F.M.; Niranatiba, T.A.; Oparinde, D.P.; Ajose, O.A.; Akindele, R.A. Free Radical Attack on Membrane Lipid and Antioxidant Vitamins in the Course of Pre-Eclamptic Pregnancy. Ethiop. J. Health. Sci. 2004, 24, 35–42. [Google Scholar] [CrossRef]
- Rota, C.; Fann, Y.C.; Mason, R.P. Phenoxyl free radical formation during the oxidation of the fluorescent dye 2′,7′-dichlorofluorescein by horseradish peroxidase Possible consequences for oxidative stress measurements. J. Biol. Chem. 1999, 274, 28161–28168. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Meng, D.; Zhang, P.; Wang, X.X.; Du, G.; Brennan, C.; Li, S.M.; Ho, C.T.; Zhao, H. Antioxidant protection of nobiletin, 5-demethylnobiletin, tangeretin, and 5-demethyltangeretin from citrus peel in Saccharomyces cerevisiae. J. Agric. Food Chem. 2018, 66, 3155–3160. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.W.; Zhang, J.; Townsend, D.M.; Tew, K.D. Oxidative stress, redox regulation and diseases of cellular differentiation. Biochim. Biophys. Acta. 1850, 1850, 1607–1621. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.H.; Shao, L.J.; Spitz, D.R. Chapter one—Reactive oxygen species in normal and tumor stem cells. Adv. Cancer Res. 2014, 122, 1–67. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, N.; Victor, V.M. Molecular strategies for targeting antioxidants to mitochondria: Therapeutic implications. Antioxid. Redox. Signal. 2015, 22, 686–729. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Zhang, P.; Zhang, L.L.; Wang, H.; Ho, C.T.; Li, S.M.; Shahidi, F.; Zhao, H. Detection of cellular redox reactions and antioxidant activity assays. J. Func. Foods 2017, 37, 467–479. [Google Scholar] [CrossRef]
- Yang, Y.C.; Li, C.K.; Lin, A.H.; Yeh, Y.W.; Yao, H.T.; Li, C.C.; Liu, K.L.; Chen, H.W. Induction of glutathione synthesis and heme oxygenase 1 by the flavonoids butein and phloretin is mediated through the ERK/Nrf2 pathway and protects against oxidative stress. Free Rad. Biol. Med. 2011, 51, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Lee, J.H.; Jegal, K.H.; Cho, J.L.; Kim, Y.W.; Kim, S.C. Oxyresveratrol abrogates oxidative stress by activating ERK–Nrf2 pathway in the liver. Chem.-Biol. Interact. 2016, 245, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.X.; Sun, Y.; Laura, T.; Liang, X.F.; Ye, H.; Zeng, X.X. Determination of polyphenolic content and antioxidant activity of kudingcha made from Ilex kudingcha C.J. Tseng. Food Chem. 2009, 112, 35–41. [Google Scholar] [CrossRef]
- Zhang, X.M. Extraction and Purification of the Polyphenols in Mallotus oblongifolius (miq.) muell.-arg and Studied the Effect for Osteoporosis Prevention and Treatment in Ovariectomized Rats. Master’s Thesis, Shanxi Academy of Traditional Chinese Medicine, Taiyuan, China, 2014. [Google Scholar]
- Zhao, Z.D.; Zhu, J.Z.; Xie, Y.L.; Dou, Z.F. Analysis of flavonoids and Hydroxycinnamic Acid Derivatives in Feng Liao of Chinese Herbal Medicine Using UPLC-PDA-ESI(-)-HRMSn Method. Asian J. Org. Med. Chem. 2017, 2, 52–61. [Google Scholar] [CrossRef]
- Silva, C.G.; Herdeiro, R.S.; Mathias, C.J.; Panek, A.D.; Silveira, C.S.; Rodrigues, V.P.; Renno, M.N.; Falcao, D.Q.; Cerqueira, D.M.; Minto, A.B.; et al. Evaluation of antioxidant activity of Brazilian plants. Pharmacol. Res. 2005, 52, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Slatnar, A.; Jakopic, J.; Stampar, F.; Veberic, R.; Jamnik, P. The effect of bioactive compounds on in vitro and in vivo antioxidant activity of different berry juices. PLoS ONE. 2012, 7, 47880–47885. [Google Scholar] [CrossRef] [PubMed]
- Steels, E.L.; Learmonth, R.P.; Watson, K. Stress tolerance and membrane lipid unsaturation in Saccharomyces cerevisiae grown aerobically or anaerobically. Microbiology 1994, 140, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.D.; Herdeiro, R.S.; Fernandes, P.N.; Eleutherio, E.C.A.; Panek, A.D. Targets of oxidative stress in yeast sod mutants. Biochim. Biophys. Acta 2003, 1620, 245–251. [Google Scholar] [CrossRef]
- Shitamukai, A.; Mizunuma, M.; Hirata, D.; Takahashi, H.; Miyakawa, T. A positive screening for drugs that specifically inhibit Ca2+-signaling activity on the basis of the growth promoting effect on a yeast mutant with a peculiar phenotype. Biosci. Biotechnol. Biochem. 2000, 64, 1942–1946. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds of MO | TR (min) | %RSD | [M − H]− (m/z) | Error (ppm) | Max (nm) | Formula | Tentatively Identified Compound |
| 1 | 1.657 | 0.277 | 169.0141 | 0.592 | 240 271 | C7H6O5 | Gallic acid [22] |
| 2 | 4.821 | 0.243 | 583.1126 | Unknown | 276 352 | Unknown | Unknown |
| 3 | 5.079 | 0.262 | 583.1138 | Unknown | 208 276 | Unknown | Unknown |
| 4 | 5.395 | 0.266 | 484.0755a | 5.164 | 281 341 | C20H20O14 | 1,6-Digalloyl glucose [23] |
| 5 | 5.629 | 0.295 | 633.0718 | 2.369 | 223 267 | C27H22O18 | 1-O-galloyl-6-O-luteoyl-α-glucose [24] |
| 6 | 6.122 | 0.239 | 388.2395a | Unknown | 246 269 | Unknown | Unknown |
| 7 | 6.314 | 0.247 | 415.2388a | 1.440 | 254 279 | C29H50O | β-sitosterol [25] |
| 8 | 6.645 | 0.278 | 275.0263 | 2.545 | 208 263 | C13H8O7 | 3,4,8,9,10-pentahydroxy-dibenzo[b,d]pyran-6-one [22] |
| 9 | 6.699 | 0.287 | 611.1583a | 3.927 | 273 208 | C27H30O16 | kaempferol-3,7-di-O-β-d-glucopyranoside [26] |
| 10 | 6.949 | 0.273 | 305.0678 | 3.606 | 276 208 | C15H14O7 | (−)-Gallocatechin (GC) [27,28] |
| 11 | 7.154 | 0.189 | 385.0843 | 5.713 | 271 208 | C19H30O8 | (6S,9R)-roseoside [29] |
| 12 | 7.53 | 0.241 | 771.197 | 2.464 | 278 211 | C33H40O21 | Quercetin-3-O-rutinoside-7-O-glucoside [30] |
| 13 | 7.801 | 0.226 | 593.1513 | 0.169 | 270 214 | C27H30O15 | Kaempferol-3-O-robinobioside [31] |
| 14 | 8.073 | 0.183 | 301.0378 | 7.972 | 246 217 | C15H10O7 | Quercetin |
| 15 | 8.274 | 0.131 | 431.0985 | 0.232 | 255 211 | C21H20O10 | Vitexin |
| 16 | 8.659 | 0.134 | 593.1507 | 0.843 | 266 208 | C27H30O15 | Kaempferol-3-O-rutinoside [31] |
| 17 | 8.952 | 0.118 | 584.2413b | 5.341 | 248 268 | C26H34O12 | Apigenin [22] |
| 18 | 9.160 | 0.100 | 477.0520 | Unknown | 273 208 | Unknown | Unknown |
| 19 | 9.368 | 0.098 | 482.0433 | Unknown | 275 208 | Unknown | Unknown |
| 20 | 9.716 | 0.080 | 485.0407 | Unknown | 228 210 | Unknown | Unknown |
| Compounds of KD | TR (min) | %RSD | [M − H]− (m/z) | Error (ppm) | Max (nm) | Formula | Tentatively Identified Compound |
| 21 | 2.521 | 1.666 | 360.1128 | Unknown | 253 326 | Unknown | Unknown |
| 22 | 2.82 | 1.534 | 353.0885 | 1.983 | 218 324 | C16H18O9 | 3-CQA [32] |
| 23 | 4.213 | 1.178 | 353.0908 | 8.496 | 215 322 | C16H18O9 | 5-CQA [32] |
| 24 | 4.507 | 1.162 | 353.0885 | 1.983 | 218 324 | C16H18O9 | 4-CQA [32] |
| 25 | 5.058 | 1.026 | 179.0365 | 8.378 | 217 321 | C9H8O4 | Caffeic acid [32] |
| 26 | 5.707 | 0.436 | 337.0928 | 0.297 | 223 293 | C16H18O8 | 4-O-p-coumaroylquinic acid [33] |
| 27 | 6.84 | 0.847 | 367.1049 | 3.012 | 221 324 | C17H20O9 | 3-FQA [32] |
| 28 | 7.525 | 0.797 | 367.1040 | 3.584 | 220 326 | C17H20O9 | 5-FQA [32] |
| 29 | 7.99 | 0.620 | 609.1508 | 7.716 | 203 255 | C27H30O16 | Rutin [32] |
| 30 | 8.309 | 0.514 | 463.0901 | 4.103 | 203 255 | C21H20O12 | Quercetin-3-O-galactoside [32] |
| 31 | 8.743 | 0.370 | 515.1203 | 1.553 | 224 279 | C25H24O12 | 3,4-diCQA [32] |
| 32 | 8.999 | 0.321 | 515.1201 | 1.165 | 224 277 | C25H24O12 | 3,5-diCQA [32] |
| 33 | 9.376 | 0.227 | 515.1204 | 1.747 | 225 275 | C25H24O12 | 4,5-diCQA [32] |
| 34 | 9.95 | 0.155 | 453.3355 | 4.191 | 218 324 | C30H46O3 | Betulonic acid [32] |
| 35 | 10.253 | 0.124 | 529.1347 | 0.945 | 221 326 | C26H26O12 | Macroanthoin G [32] |
| 36 | 10.444 | 0.105 | 529.1336 | 3.024 | 219 330 | C26H26O12 | Unknown |
| 37 | 10.88 | 0.090 | 543.1523 | Unknown | 221 326 | Unknown | Unknown |
| 38 | 11.395 | 0.071 | 647.3196a | Unknown | 260 326 | Unknown | Unknown |
| 39 | 11.574 | 0.065 | 469.3701a | 8.496 | 251 326 | C30H44O4 | a-Kudinlactone [32] |
| Compounds of DY | TR (min) | %RSD | [M − H]− (m/z) | Error (ppm) | Max (nm) | Formula | Tentatively Identified Compound |
| 40 | 1.452 | 0.332 | 343.0685 | 4.081 | 203 270 | C14H16O10 | 5-GQA [27] |
| 41 | 1.596 | 0.339 | 169.0149 | 4.142 | 215 271 | C7H6O5 | Gallic acid [22] |
| 42 | 2.194 | 0.609 | 305.067 | 0.983 | 204 271 | C15H14O7 | (−)-Gallocatechin (GC) [27,28] |
| 43 | 3.226 | 0.686 | 353.0888 | 2.832 | 206 274 | C16H18O9 | 3-CQA [32] |
| 44 | 3.605 | 0.452 | 305.0671 | 1.311 | 204 270 | C15H14O7 | (−)-Epigallocatechin(EGC) |
| 45 | 4.249 | 0.433 | 289.0711 | 2.422 | 204 217 | C15H14O6 | (+)-Catechin (C) |
| 46 | 4.707 | 0.304 | 195.0727a | 2.051 | 224 273 | C8H10N4O2 | Caffeine |
| 47 | 5.695 | 0.216 | 289.0733 | 5.189 | 220 277 | C15H14O6 | (−)-Epicatechin(EC) |
| 48 | 6.289 | 0.270 | 457.0771 | 1.094 | 204 274 | C22H18O11 | (−)-Epigallocatechin-3-gallate (EGCG) |
| 49 | 7.01 | 0.466 | 563.1143 | 9.234 | 200 272 | C29H24O12 | Theaflavin [34] |
| 50 | 7.458 | 0.239 | 471.3439 | 8.698 | 206 275 | C30H48O4 | Pomolic acid [35] |
| 51 | 8.042 | 0.195 | 441.0842 | 3.401 | 219 275 | C22H18O10 | (−)-Epicatechin-3-gallate (ECG) |
| 52 | 8.23 | 0.195 | 441.0831 | 0.907 | 204 276 | C22H18O10 | (−)-Catechin-3-gallate (CG) |
| 53 | 8.88 | 0.322 | 593.1499 | 2.192 | 198 266 | C27H30O15 | Kaempferol-3-O-rutinoside [31] |
| 54 | 9.244 | 0.246 | 455.0978 | 1.318 | 203 278 | C23H20O10 | (−)-Catechin-3-O-(4-O-methyl) gallate [36] |
| 55 | 9.655 | 0.242 | 455.0979 | 1.099 | 205 278 | C23H20O10 | (−)-Epicatechin-3-O-(4-O-methyl) gallate [36] |
| Compounds of BS | TR (min) | %RSD | [M − H]−(m/z) | Error(ppm) | Max(nm) | Formula | Tentatively Identified Compound |
| 56 | 1.458 | 0.439 | 343.0645 | 7.579 | 213 273 | C14H16O10 | 5-GQA [27] |
| 57 | 1.595 | 0.374 | 343.067 | 0.291 | 214 271 | C14H16O10 | 3-GQA [27] |
| 58 | 2.202 | 0.644 | 305.0652 | 4.917 | 203 272 | C15H14O7 | (−)-Gallocatechin (GC) [27,28] |
| 59 | 3.616 | 0.545 | 305.0663 | 1.311 | 204 270 | C15H14O7 | (−)-Epigallocatechin (EGC) |
| 60 | 4.255 | 0.457 | 289.0729 | 3.805 | 204 280 | C15H14O6 | (+)-Catechin (C) |
| 61 | 4.718 | 0.441 | 195.0725a | 3.076 | 222 273 | C8H10N4O2 | Caffeine |
| 62 | 5.712 | 0.384 | 457.0771 | 1.094 | 216 274 | C22H18O11 | (−)-Epigallocatechin-3-gallate (EGCG) |
| 63 | 6.269 | 0.332 | 457.0763 | 2.844 | 218 274 | C22H18O11 | (−)-Gallocatechin-3-gallate (GCG) |
| 64 | 6.971 | 0.186 | 563.1193 | 0.355 | 203 271 | C29H24O12 | Theaflavin [34] |
| 65 | 7.072 | 0.291 | 479.0838 | 1.461 | 205 265 | C21H20O13 | Myricetin 3-O-β-l-galactopyranoside [35] |
| 66 | 7.597 | 0.228 | 771.2002 | 1.686 | 203 256 | C33H40O21 | Quercetin-3-O-rutinoside-7-O-glucoside [30] |
| 67 | 8.018 | 0.211 | 441.0829 | 0.453 | 220 274 | C22H18O10 | (−)-Epicatechin-3-gallate (ECG) |
| 68 | 8.199 | 0.200 | 441.0834 | 1.587 | 204 275 | C22H18O10 | (−)-Catechin-3-gallate (CG) |
| 69 | 8.418 | 0.180 | 463.0884 | 0.432 | 200 266 | C21H20O12 | Quercetin-3-O-galactoside [32] |
| 70 | 8.849 | 0.120 | 593.1534 | 3.709 | 199 266 | C27H30O15 | Kaempferol-3-O-rutinoside [31] |
| 71 | 9.134 | 0.103 | 447.0925 | 1.789 | 203 267 | C21H20O11 | Kaempferol-3-O-glucoside [27] |
| Varieties | MO | KD | DY | BS | ||||
|---|---|---|---|---|---|---|---|---|
| TPC (mg GAE/g) | TFC (mg RE/g) | TPC (mg GAE/g) | TFC (mg RE/g) | TPC (mg GAE/g) | TFC (mg RE/g) | TPC (mg GAE/g) | TFC (mg RE/g) | |
| Water extract | 353.83 ± 6.49b * | 215.36 ± 7.00a | 186.41 ± 3.47c | 742.00 ± 4.65b * | 326.55 ± 3.21c * | 556.82 ± 26.48c | 251.26 ± 5.30c | 298.67 ± 6.43c |
| Resin extract | 458.83 ± 5.42a | 215.70 ± 2.92a | 221.69 ± 0.84b | 786.82 ± 1.56ab * | 551.12 ± 5.24b * | 782.37 ± 9.79b * | 428.83 ± 3.74b | 499.41 ± 0.76b |
| Fraction I | 449.26 ± 7.49a | 288.30 ± 0.31a | 279.12 ± 1.83a | 858.30 ± 2.79a * | 659.83 ± 1.71a * | 974.22 ± 5.31a * | 526.26 ± 3.00a | 602.00 ± 2.41a |
| Fraction II | 433.98 ± 3.78a | 237.56 ± 6.83a | 158.12 ± 4.35d | 446.82 ± 2.01c | 669.55 ± 4.74a * | 960.15 ± 5.87a * | 199.83 ± 4.39d | 204.59 ± 7.91d |
| Components | IC50 (μg/mL) DPPH· Scavenging Activity | IC50 (mg/mL) HO· Scavenging Activity |
|---|---|---|
| MO-water | 13.93 ± 0.60 de | 1.97 ± 0.05 de |
| MO-resin | 6.54 ± 0.71 fg | 1.30 ± 0.17 efg |
| MO-fraction I | 8.33 ± 0.45 f | 1.21 ± 0.46 efg |
| MO-fraction II | 8.22 ± 0.98 f | 1.62 ± 0.30 def |
| KD-water | 35.04 ± 0.40 b | 5.16 ± 0.88 c |
| KD-resin | 18.75 ± 1.30 c | 4.95 ± 0.41 c |
| KD-fraction I | 20.26 ± 0.63 c | 6.60 ± 0.87 b |
| KD-fraction II | 47.85 ± 1.93 a | 8.11 ± 0.33 a |
| DY-water | 4.71 ± 0.15 gh | 2.15 ± 0.13 d |
| DY-resin | 4.57 ± 0.09 gh | 1.81 ± 0.40 def |
| DY-fraction I | 12.25 ± 1.76 e | 1.58 ± 0.24 def |
| DY-fraction II | 4.13 ± 0.70 h | 1.44 ± 0.08 def |
| BS-water | 15.48 ± 1.48 d | 1.31 ± 0.36 efg |
| BS-resin | 15.93 ± 0.96 d | 1.11 ± 0.05 fg |
| BS-fraction I | 12.20 ± 1.55 e | 0.63 ± 0.17 g |
| BS-fraction II | 19.74 ± 1.95 c | 1.58 ± 0.26 def |
| FA | 5.77 ± 1.23 gh | 1.37 ± 0.13 ef |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Wang, L.; Luo, Y. Composition Analysis by UPLC-PDA-ESI (−)-HRMS and Antioxidant Activity Using Saccharomyces cerevisiae Model of Herbal Teas and Green Teas from Hainan. Molecules 2018, 23, 2550. https://doi.org/10.3390/molecules23102550
Li H, Wang L, Luo Y. Composition Analysis by UPLC-PDA-ESI (−)-HRMS and Antioxidant Activity Using Saccharomyces cerevisiae Model of Herbal Teas and Green Teas from Hainan. Molecules. 2018; 23(10):2550. https://doi.org/10.3390/molecules23102550
Chicago/Turabian StyleLi, Hua, Lanying Wang, and Yanping Luo. 2018. "Composition Analysis by UPLC-PDA-ESI (−)-HRMS and Antioxidant Activity Using Saccharomyces cerevisiae Model of Herbal Teas and Green Teas from Hainan" Molecules 23, no. 10: 2550. https://doi.org/10.3390/molecules23102550
APA StyleLi, H., Wang, L., & Luo, Y. (2018). Composition Analysis by UPLC-PDA-ESI (−)-HRMS and Antioxidant Activity Using Saccharomyces cerevisiae Model of Herbal Teas and Green Teas from Hainan. Molecules, 23(10), 2550. https://doi.org/10.3390/molecules23102550