Biodetoxification of Phenolic Inhibitors from Lignocellulose Pretreatment using Kurthia huakuii LAM0618T and Subsequent Lactic Acid Fermentation

Abstract
:1. Introduction
2. Results
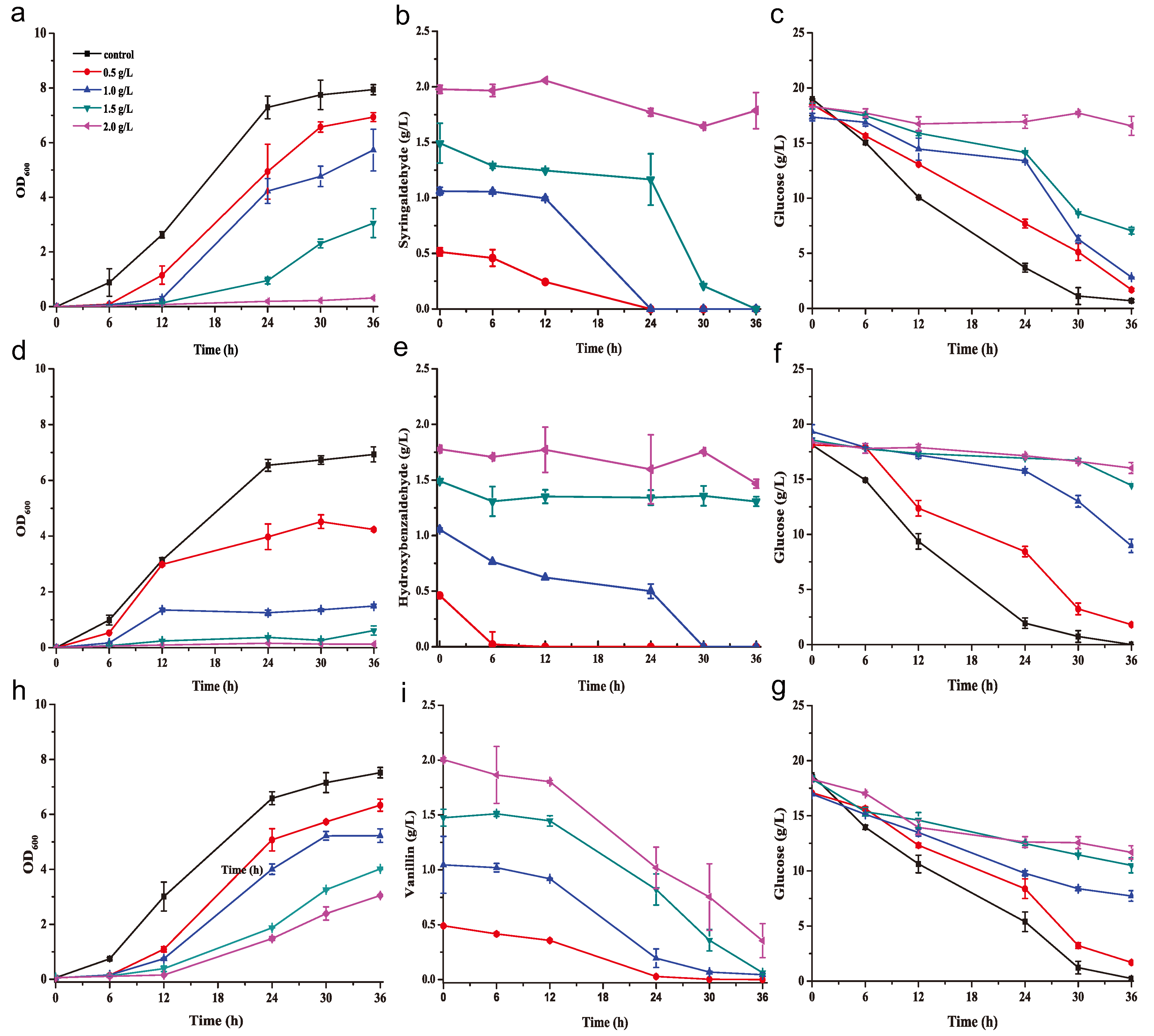
2.1. K. huakuii LAM0618T Degrades Phenolic Inhibitors in a Rich Medium
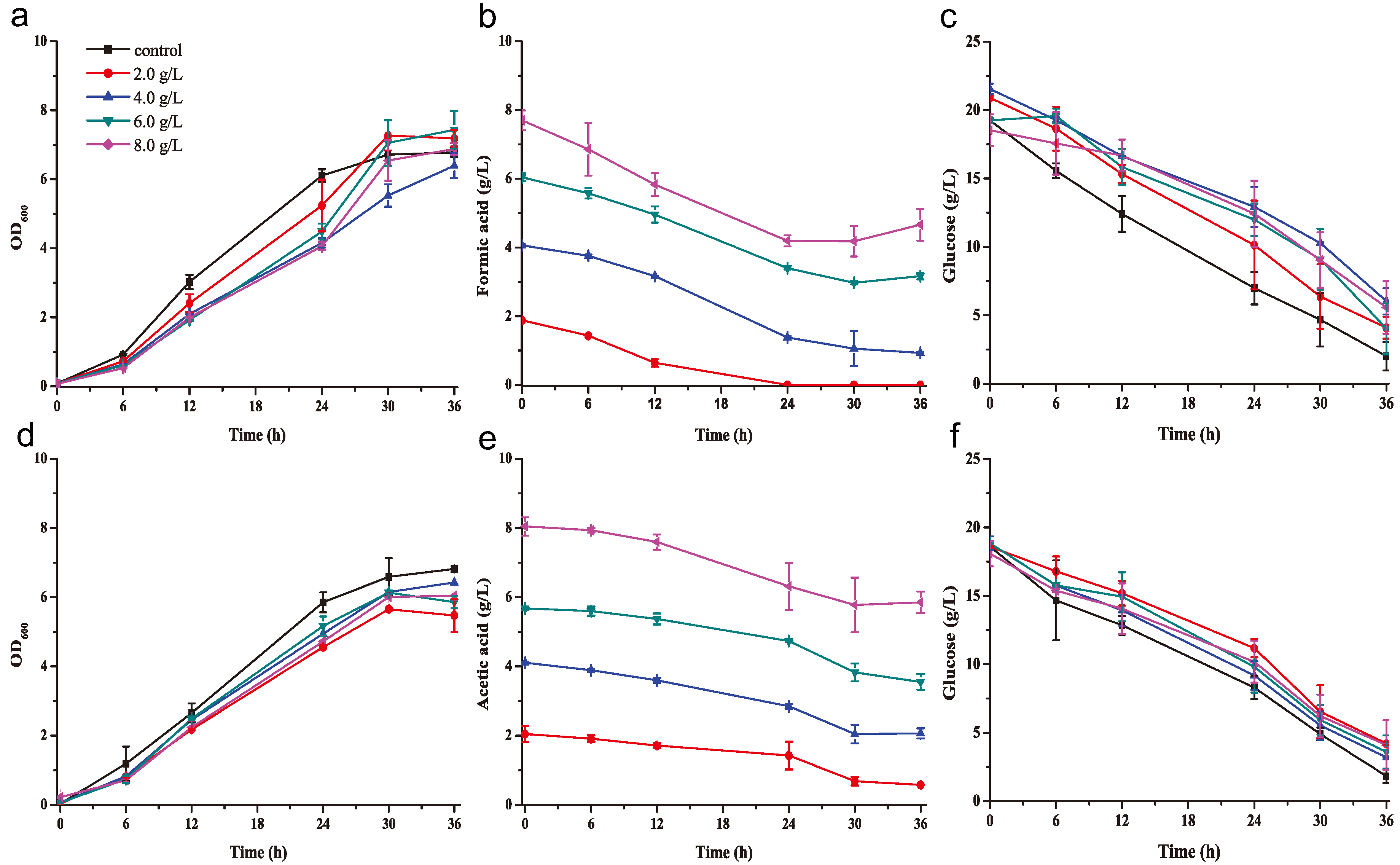
2.2. K. huakuii LAM0618T Degrades Formate and Acetate in a Rich Medium
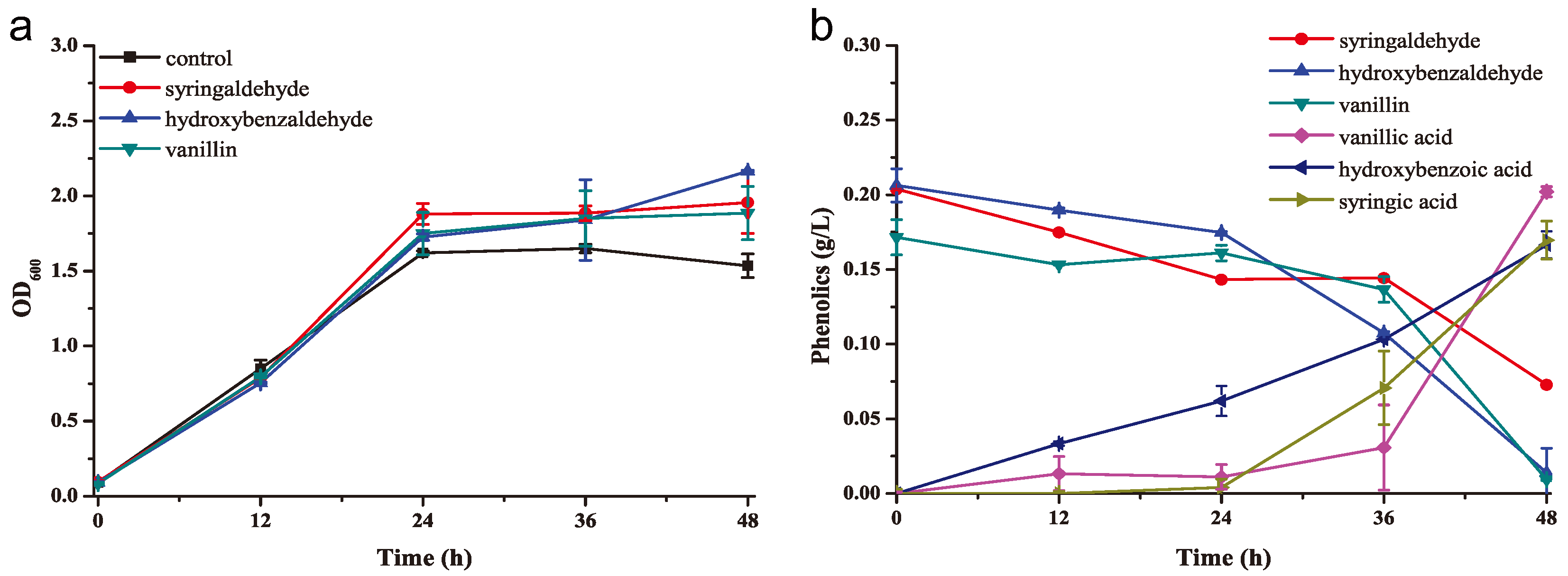
2.3. K. huakuii LAM0618T Trzansformed Phenolic Inhibitors into Less Toxic Acids
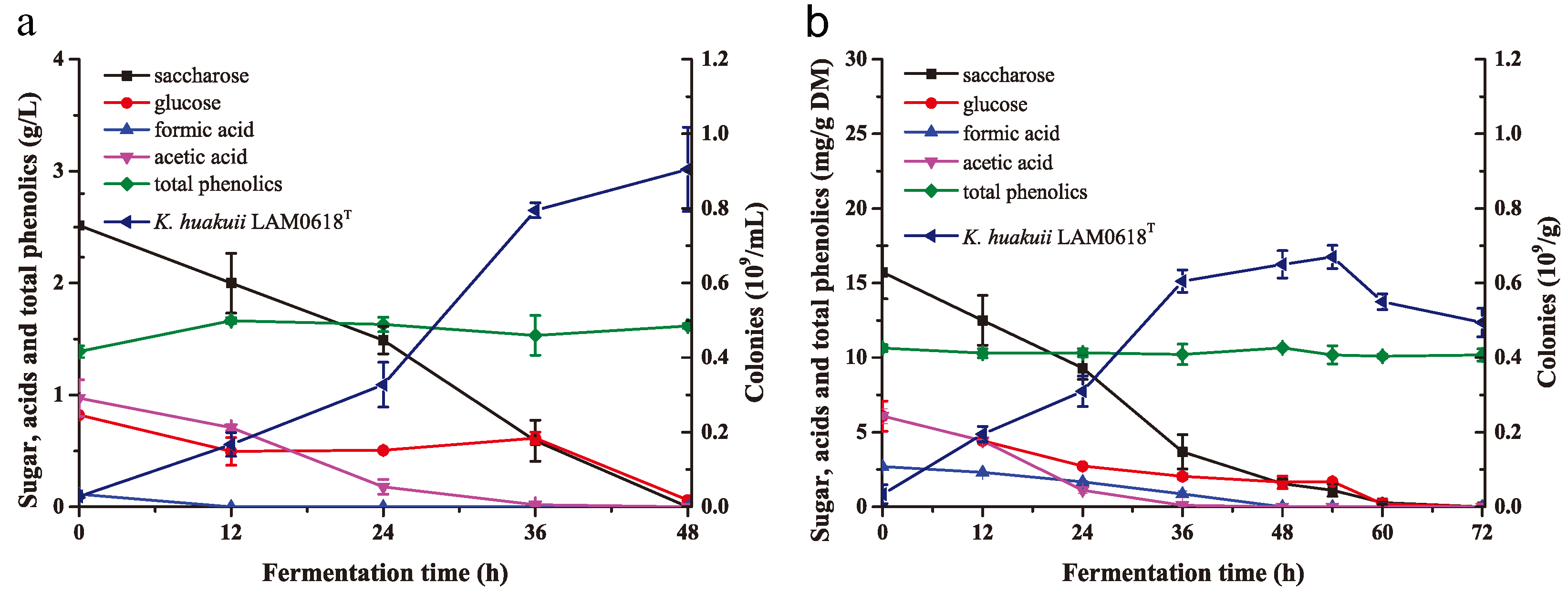
2.4. Biodetoxification of NH3/H2O2-Pretreated Corn Stover by K. huakuii LAM0618T
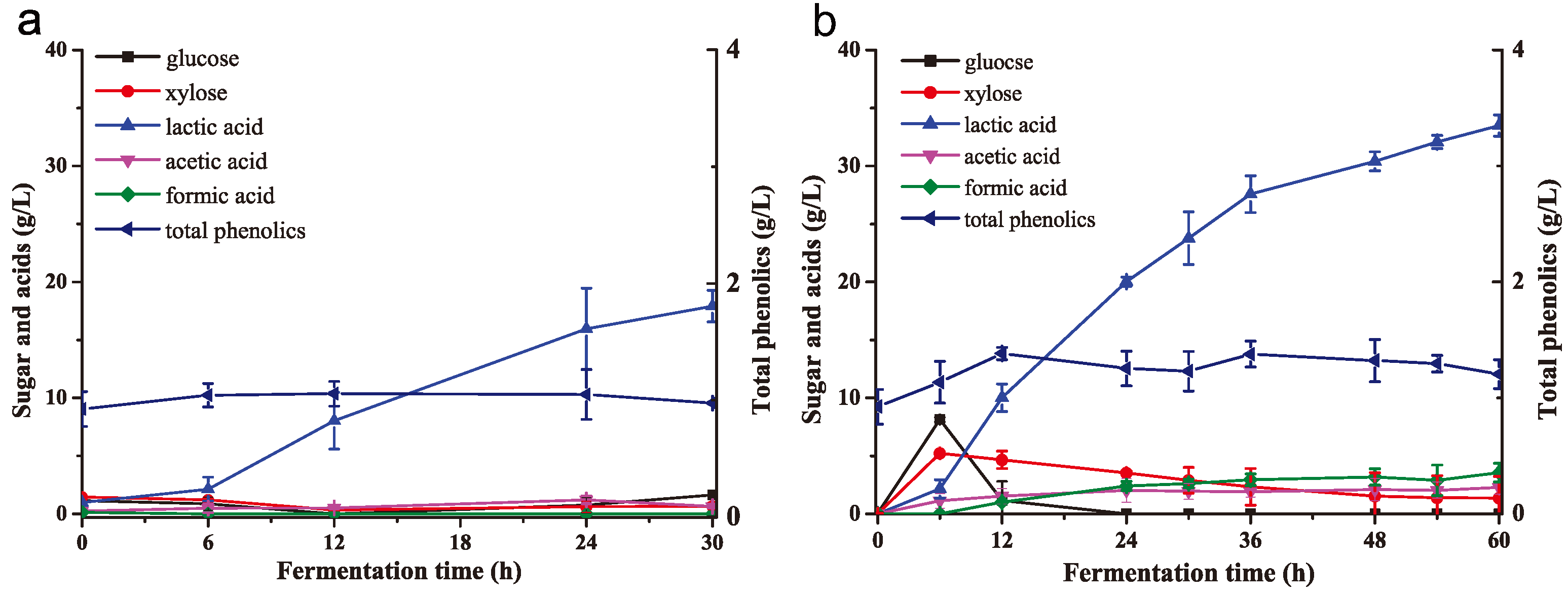
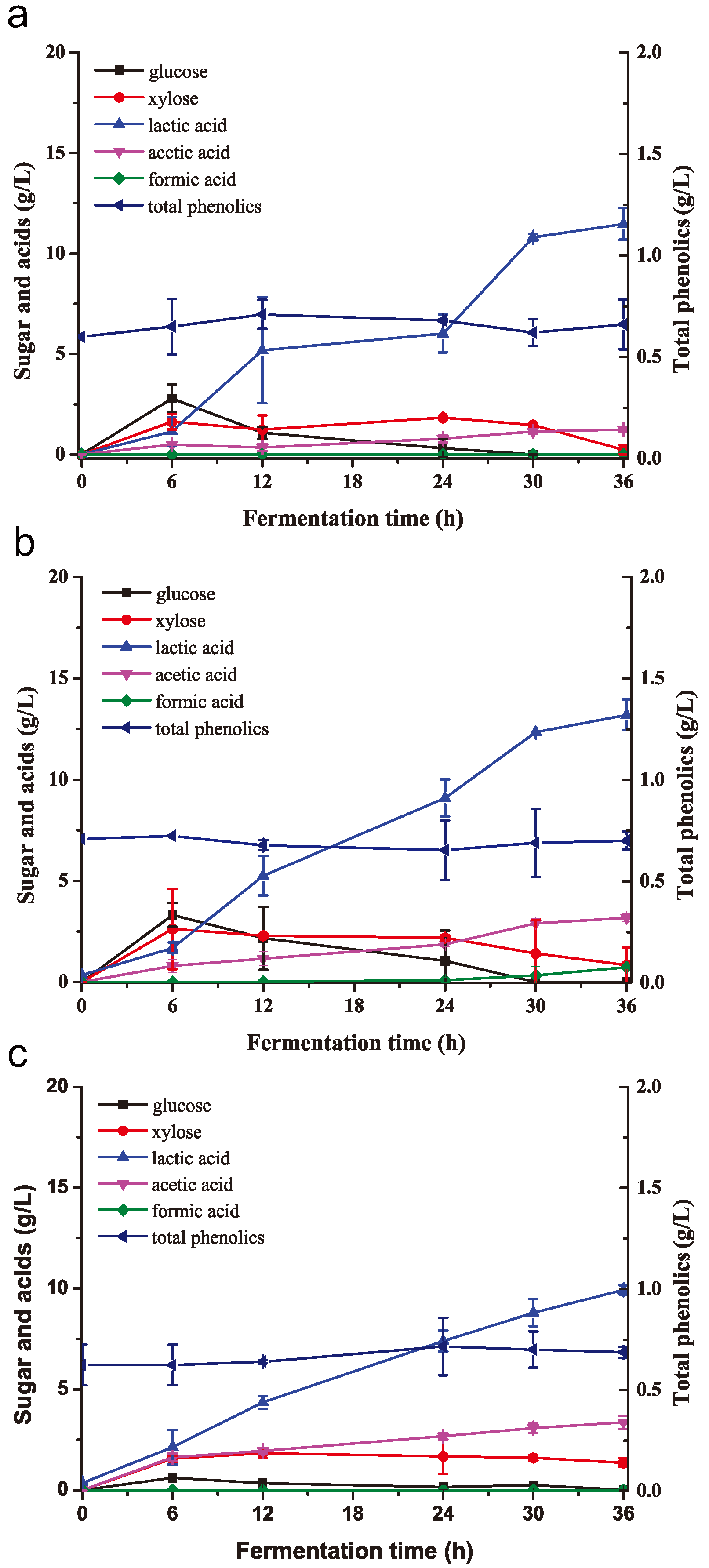
2.5. Lactic Acid Fermentation from Biodetoxified and Non-Detoxified Corn Stover in SSF
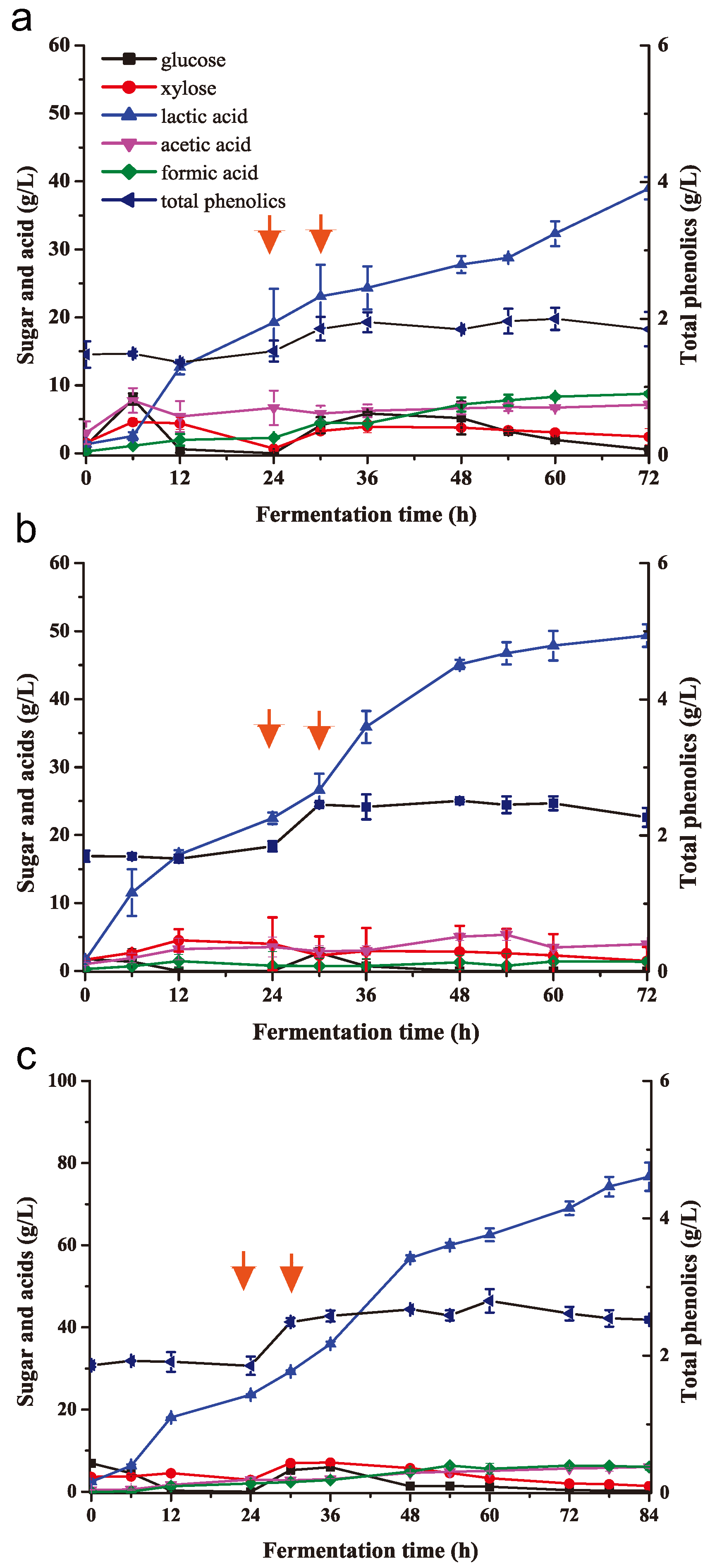
2.6. Lactic Acid Fermentation at High-Solid Loading of Biodetoxified Corn Stover in Fed-Batch SSF
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Raw Material and Pretreatments
4.3. Enzymes and Reagents
4.4. Analysis of K. huakuii LAM0618T Growth on Phenolic Compounds and formate/Actetae in the Presence of Glucose
4.5. Analysis of K. huakuii LAM0618T Growth with Phenolic Compounds and Formate/Acetate as the Carbon Sources
4.6. Biodetoxification of Alkaline-Pretreated Corn Stover
4.7. Co-Culture of K. huakuii LAM0618T and B. coagulans LA204 for Lactic Acid Production
4.8. Lactic Acid Fermentation of Biodetoxified and Non-Detoxified Corn Stover by SSF
4.9. Analysis of Sugars, Acids, Inhibitors, and Laccase Activity
4.10. Statistical Tests
Author Contributions
Funding
Conflicts of Interest
References
- Hu, J.; Lin, Y.; Zhang, Z.; Xiang, T.; Mei, Y.; Zhao, S.; Liang, Y.; Peng, N. High-titer lactic acid production by Lactobacillus pentosus FL0421 from corn stover using fed-batch simultaneous saccharification and fermentation. Bioresour. Technol. 2016, 214, 74–80. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, J.; Bao, J. Acceleration of biodetoxification on dilute acid pretreated lignocellulose feedstock by aeration and the consequent ethanol fermentation evaluation. Biotechnol. Biofuels 2016, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, Q.; Zhang, H.; Bao, J. Inhibitor degradation and lipid accumulation potentials of oleaginous yeast Trichosporon cutaneum using lignocellulose feedstock. Bioresour. Technol. 2016, 218, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, K.; Laser, J.; Sinskey, A.J. Tolerance and adaptive evolution of triacylglycerol-producing Rhodococcus opacus to lignocellulose-derived inhibitors. Biotechnol. Biofuels 2015, 8, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talebnia, F.; Karakashev, D.; Angelidaki, I. Production of bioethanol from wheat straw: An overview on pretreatment, hydrolysis and fermentation. Bioresour. Technol. 2010, 101, 4744–4753. [Google Scholar] [CrossRef] [PubMed]
- Uppugundla, N.; da Costa Sousa, L.; Chundawat, S.P.; Yu, X.; Simmons, B.; Singh, S.; Gao, X.; Kumar, R.; Wyman, C.E.; Dale, B.E.; et al. A comparative study of ethanol production using dilute acid, ionic liquid and AFEX pretreated corn stover. Biotechnol. Biofuels 2014, 7, 72. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Mochidzuki, K.; Cheng, W.; Zhou, M.; Gao, H.; Zheng, D.; Wang, X.; Cui, Z. Effects of different pretreatment strategies on corn stalk acidogenic fermentation using a microbial consortium. Bioresour. Technol. 2011, 102, 7526–7531. [Google Scholar] [CrossRef] [PubMed]
- Toquero, C.; Bolado, S. Effect of four pretreatments on enzymatic hydrolysis and ethanol fermentation of wheat straw. Influence of inhibitors and washing. Bioresour. Technol. 2014, 157, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xie, Y.; He, X.; Li, X.; Hu, J.; Ruan, Z.; Zhao, S.; Peng, N.; Liang, Y. Comparison of high-titer lactic acid fermentation from NaOH- and NH3-H2O2-pretreated corncob by Bacillus coagulans using simultaneous saccharification and fermentation. Sci. Rep. 2016, 6, 37245. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, L.J.; Alriksson, B.; Nilvebrant, N.O. Bioconversion of lignocellulose: Inhibitors and detoxification. Biotechnol Biofuels 2013, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.K.; Um, Y.; Park, Y.C.; Seo, J.H.; Kim, K.H. Compounds inhibiting the bioconversion of hydrothermally pretreated lignocellulose. Appl. Microbiol. Biotechnol. 2015, 99, 4201–4212. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Ximenes, E.; Mosier, N.S.; Ladisch, M.R. Soluble inhibitors/deactivators of cellulase enzymes from lignocellulosic biomass. Enzyme Microb. Technol. 2011, 48, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Li, W.C.; Liu, L.; Zhu, J.Q.; Li, X.; Li, B.Z.; Yuan, Y.J. Inhibition of lignin-derived phenolic compounds to cellulase. Biotechnol. Biofuels 2016, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yong, Q.; Xu, Y.; Yu, S. Detoxification of corn stover prehydrolyzate by trialkylamine extraction to improve the ethanol production with Pichia stipitis CBS 5776. Bioresour. Technol. 2011, 102, 1663–1668. [Google Scholar] [CrossRef] [PubMed]
- Florencio, C.; Badino, A.C.; Farinas, C.S. Soybean protein as a cost-effective lignin-blocking additive for the saccharification of sugarcane bagasse. Bioresour. Technol. 2016, 221, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Ximenes, E.; Nichols, N.N.; Zhang, L.; Ladisch, M. Biological abatement of cellulase inhibitors. Bioresour. Technol. 2013, 146, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Kunasundari, B.; Arai, T.; Sudesh, K.; Hashim, R.; Sulaiman, O.; Stalin, N.J.; Kosugi, A. Detoxification of sap from felled oil palm trunks for the efficient production of lactic acid. Appl. Biochem. Biotechnol. 2017, 183, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.; Carneiro, L.M.; Roberto, I.C. Treatment of rice straw hemicellulosic hydrolysates with advanced oxidative processes: A new and promising detoxification method to improve the bioconversion process. Biotechnol. Biofuels 2013, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhu, Z.; Wang, X.; Wang, N.; Wang, W.; Bao, J. Biodetoxification of toxins generated from lignocellulose pretreatment using a newly isolated fungus, Amorphotheca resinae ZN1, and the consequent ethanol fermentation. Biotechnol. Biofuels 2010, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Gong, G.; Park, H.S.; Um, Y.; Sim, S.J.; Woo, H.M. Extreme furfural tolerance of a soil bacterium Enterobacter cloacae GGT036. J. Biotechnol. 2015, 193, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, B.G.; Moutta Rde, O.; Ferraz Fde, O.; Vieira, E.R.; Nogueira, A.S.; Baratella, B.F.; Rodrigues, L.C.; Hou-Rui, Z.; da Silva, S.S. Biological detoxification of different hemicellulosic hydrolysates using Issatchenkia occidentalis CCTCC M 206097 yeast. J. Ind. Microbiol. Biotechnol. 2011, 38, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wu, H.; Smith, T.J.; Liu, Z.J.; Lou, W.Y.; Zong, M.H. In vivo detoxification of furfural during lipid production by the oleaginous yeast Trichosporon fermentans. Biotechnol. Lett. 2012, 34, 1637–1642. [Google Scholar] [CrossRef] [PubMed]
- Ran, H.; Zhang, J.; Gao, Q.; Lin, Z.; Bao, J. Analysis of biodegradation performance of furfural and 5-hydroxymethylfurfural by Amorphotheca resinae ZN1. Biotechnol. Biofuels 2014, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Sun, J.; Zhang, J.; Tu, Y.; Bao, J. High titer l-lactic acid production from corn stover with minimum wastewater generation and techno-economic evaluation based on Aspen plus modeling. Bioresour. Technol. 2015, 198, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Zhang, P.; Sun, J.; Tu, Y.; Gao, Q.; Zhang, J.; Bao, J. Engineering wild-type robust Pediococcus acidilactici strain for high titer l- and d-lactic acid production from corn stover feedstock. J. Biotechnol. 2016, 217, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.; Wang, Y.; Song, J.; Jiang, S.; Wang, H.; Li, Y.; Zhao, B.; Jiang, R.; Zhao, B. Kurthia huakuii sp. nov., isolated from biogas slurry, and emended description of the genus Kurthia. Int. J. Syst. Evol. Microbiol. 2014, 64, 18–521. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhou, S.; Wang, Y.; Song, J.; Wang, H.; Kong, D.; Zhu, J.; Dong, W.; He, M.; Hu, G.; et al. Characterization of a highly thermostable and organic solvent-tolerant copper-containing polyphenol oxidase with dye-decolorizing ability from Kurthia huakuii LAM0618T. PLoS ONE 2016, 11, e0164810. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, Z.; Lin, Y.; Zhao, S.; Mei, Y.; Liang, Y.; Peng, N. High-titer lactic acid production from NaOH-pretreated corn stover by Bacillus coagulans LA204 using fed-batch simultaneous saccharification and fermentation under non-sterile condition. Bioresour. Technol. 2015, 182, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Li, Y.; Wan, C. Lactic acid production from corn stover using mixed cultures of Lactobacillus rhamnosus and Lactobacillus brevis. Bioresour. Technol. 2011, 102, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Vadlani, P.V.; Kumar, A.; Hardwidge, P.R.; Govind, R.; Tanaka, T.; Kondo, A. Enhanced d-lactic acid production from renewable resources using engineered Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 2016, 100, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Qiao, Q.; Chu, D.; Gu, H.; Dao, T.H.; Zhang, J.; Bao, J. Simultaneous saccharification and high titer lactic acid fermentation of corn stover using a newly isolated lactic acid bacterium Pediococcus acidilactici DQ2. Bioresour. Technol. 2013, 135, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Van der Pol, E.C.; Eggink, G.; Weusthuis, R.A. Production of l(+)-lactic acid from acid pretreated sugarcane bagasse using Bacillus coagulans DSM2314 in a simultaneous saccharification and fermentation strategy. Biotechnol. Biofuels 2016, 9, 248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, X.; Luo, J.; Qi, B.; Wan, Y. An efficient process for lactic acid production from wheat straw by a newly isolated Bacillus coagulans strain IPE22. Bioresour. Technol. 2014, 158, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
Sample Availability: Samples of the compounds phenolics, acids, sugars and corn stover are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Detoxified or Non-Detoxified Corn Stover | Non-Detoxified | Detoxified | Initial: Detoxified; Fed: Non-Detoxified | Initial: Detoxified; Fed: Detoxified | Initial: Detoxified; Fed: Detoxified |
|---|---|---|---|---|---|
| Corn stover concentration (w/w) | 8% | 8% | 8–12% | 8–12% | 8–15% |
| Fermentation time (h) | 30 c | 60 | 72 | 72 | 96 |
| Lactic acid titer (g/L) | 17.91 ± 1.11 | 33.47 ± 1.33 | 38.99 ± 1.64 | 49.35 ± 1.67 | 84.49 ± 0.95 |
| Lactic acid yield (g/g) a | 0.22 | 0.42 | 0.32 | 0.41 | 0.56 |
| Lactic acid productivity (g/L/h) b | 0.60 | 0.56 | 0.54 | 0.69 | 0.88 |
| Acetic acid titer (g/L) | 0.65 ± 0.12 | 2.28 ± 0.56 | 7.14 ± 0.48 | 3.97 ± 0.17 | 5.39 ± 0.17 |
| Acetic acid yield (g/g) a | 0.01 | 0.03 | 0.06 | 0.03 | 0.04 |
| Acetic acid productivity (g/L/h) b | 0.02 | 0.04 | 0.10 | 0.06 | 0.06 |
| α (A) | 0.01 | 0.05 | - | ||
| p-value | 0.005 | 0.024 | - | ||
| Fermentation Strains | Substrate | Pretreatment | Fermentation Mode | Lactic Acid | Detoxification Mode | Detoxification Strains | Ref. | ||
|---|---|---|---|---|---|---|---|---|---|
| Titer (g/L) | Yield (g/g) | Productivity (g/L/h) | |||||||
| Lb. pentosus FL0421 | Corn stover | NaOH | Fed-batch SSF | 92.3 | 0.66 a | 1.92 | Water rinsing | [1] | |
| Lb. rhamnosus and Lb. brevis | Corn stover | NaOH | Fed-batch SSF | 60.3 | 0.70 a | 0.58 | Water rinsing | [29] | |
| B. coagulans LA204 | Corn stover | NaOH | Fed-batch SSF | 97.59 | 0.68 a | 1.63 | Water rinsing | [28] | |
| Lb. plantarum NCIMB 8826 | Corn stover | NaOH | Fed-batch SSF | 61.4 | 0.77 b | 0.32 | None | [30] | |
| P. acidilactici DQ2 | Corn stover | Dilute acid | SSF | 101.9 | 0.77 c | 1.06 | Bio-detoxification | A. resinae ZN1 | [31] |
| P. acidilactici TY112 | Corn stover | Dilute acid | SSF | 77.66 | 0.65 c | 1.06 | Bio-detoxification | A. resinae ZN1 | [25] |
| B. coagulans DSM2314 | Sugarcane bagasse | Dilute acid | Fed-batch SSF | 58.7 | 0.73 a | 1.81 | Bio-detoxification | B. coagulans DSM2314 | [32] |
| B. coagulans IPE22 | Wheat straw | Dilute acid | SSF | 46.12 | 0.46 a | 0.42 | Bio-detoxification | B. coagulans IPE22 | [33] |
| B. coagulans LA204 | Corn stover | NH3/H2O2 | Fed-batch SSF | 84.49 | 0.56 a | 0.99 | Bio-detoxification | K. huakuii LAM0618T | This study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Y.; Hu, Q.; Feng, G.; Jiang, X.; Hu, J.; He, M.; Hu, G.; Zhao, S.; Liang, Y.; Ruan, Z.; et al. Biodetoxification of Phenolic Inhibitors from Lignocellulose Pretreatment using Kurthia huakuii LAM0618T and Subsequent Lactic Acid Fermentation. Molecules 2018, 23, 2626. https://doi.org/10.3390/molecules23102626
Xie Y, Hu Q, Feng G, Jiang X, Hu J, He M, Hu G, Zhao S, Liang Y, Ruan Z, et al. Biodetoxification of Phenolic Inhibitors from Lignocellulose Pretreatment using Kurthia huakuii LAM0618T and Subsequent Lactic Acid Fermentation. Molecules. 2018; 23(10):2626. https://doi.org/10.3390/molecules23102626
Chicago/Turabian StyleXie, Yuejiao, Qing Hu, Guodong Feng, Xu Jiang, Jinlong Hu, Mingxiong He, Guoquan Hu, Shumiao Zhao, Yunxiang Liang, Zhiyong Ruan, and et al. 2018. "Biodetoxification of Phenolic Inhibitors from Lignocellulose Pretreatment using Kurthia huakuii LAM0618T and Subsequent Lactic Acid Fermentation" Molecules 23, no. 10: 2626. https://doi.org/10.3390/molecules23102626
APA StyleXie, Y., Hu, Q., Feng, G., Jiang, X., Hu, J., He, M., Hu, G., Zhao, S., Liang, Y., Ruan, Z., & Peng, N. (2018). Biodetoxification of Phenolic Inhibitors from Lignocellulose Pretreatment using Kurthia huakuii LAM0618T and Subsequent Lactic Acid Fermentation. Molecules, 23(10), 2626. https://doi.org/10.3390/molecules23102626