Anti-Inflammatory Activities of Pentaherbs formula and Its Influence on Gut Microbiota in Allergic Asthma

 ,
, 
Abstract
:
1. Introduction
2. Results
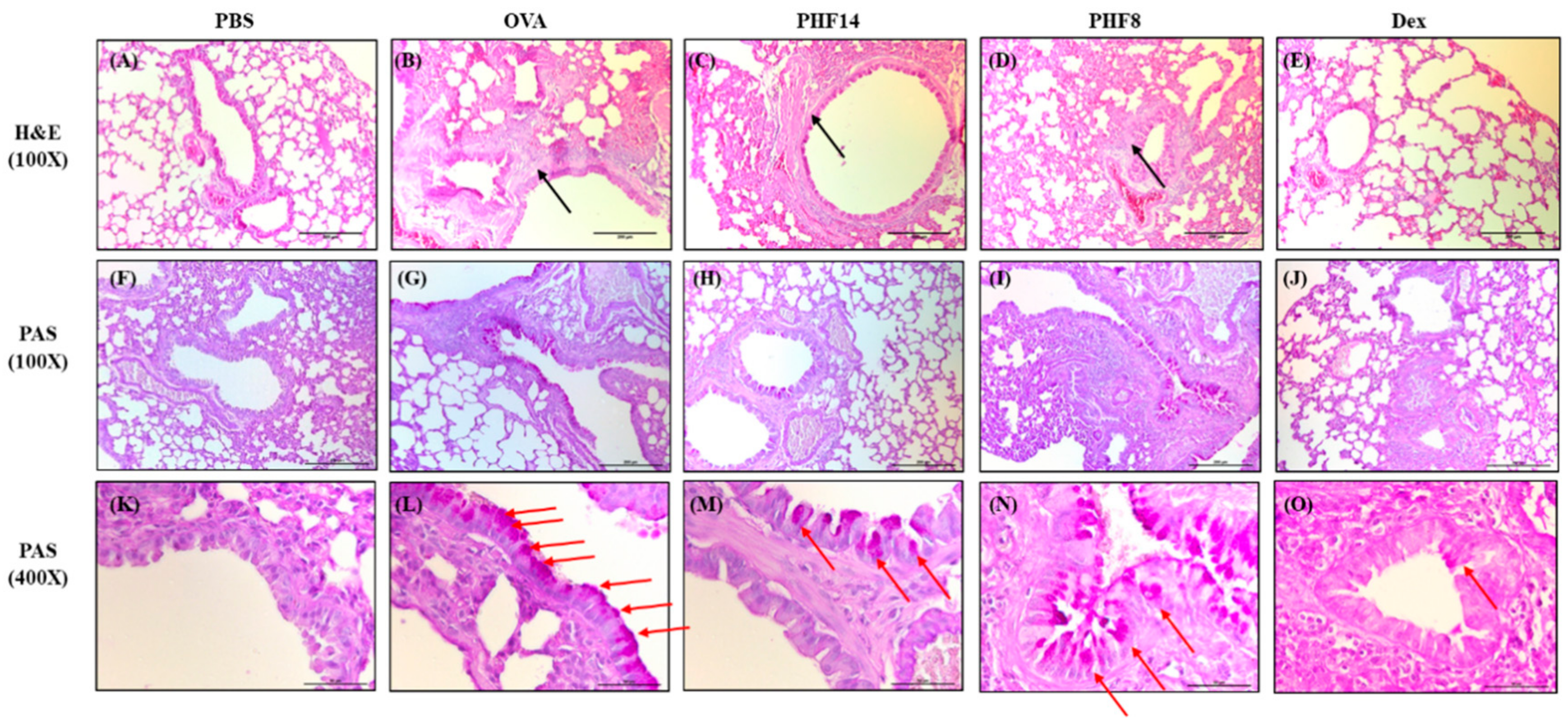
2.1. Pentaherbs Formula Reduced Serum OVA-Specific IgE, Airway Hyperresponsiveness (AHR) and Airway Wall Remodeling of OVA-Induced Allergic Asthmatic Mice
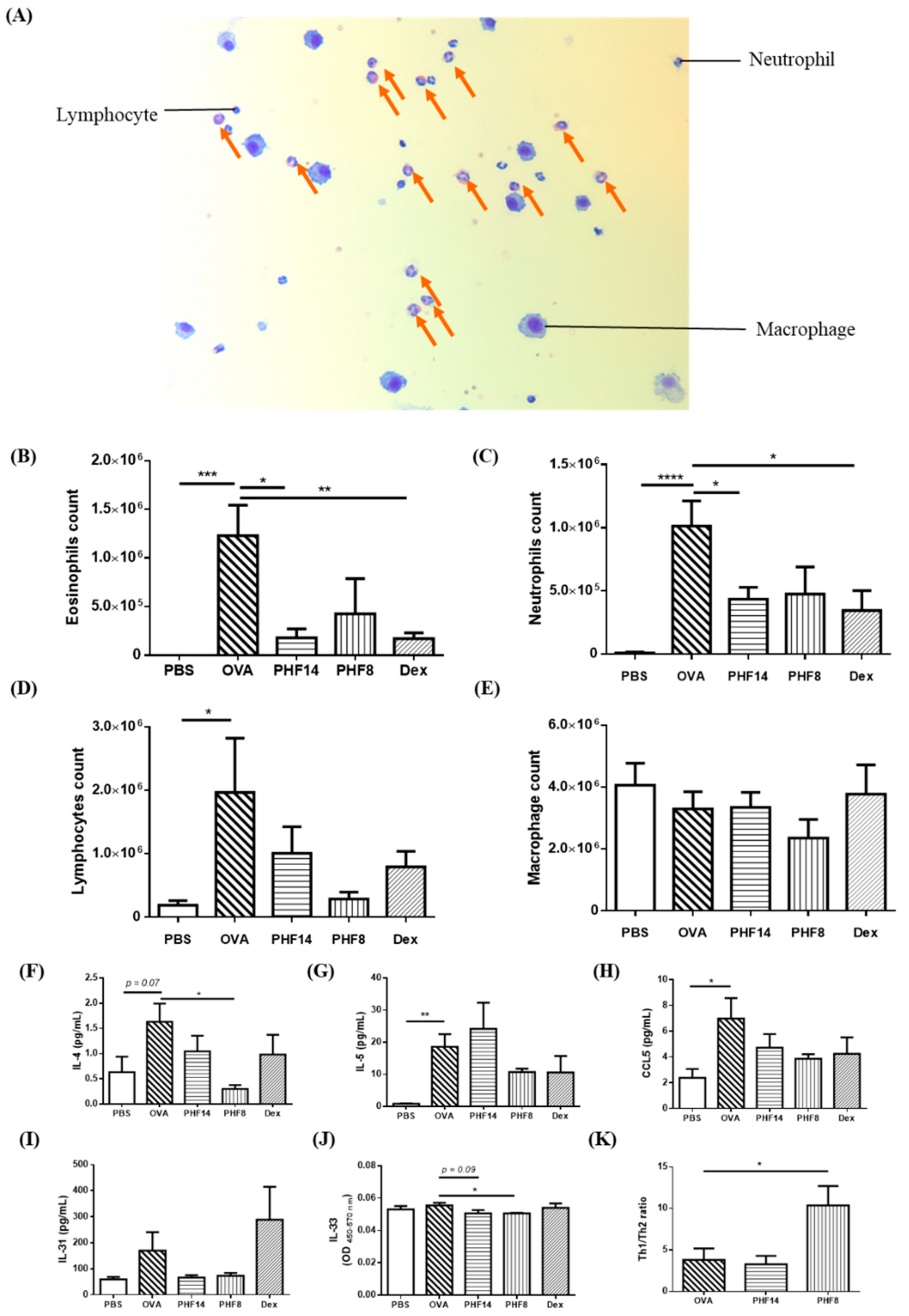
2.2. Pentaherbs Formula Significantly Suppressed the Infiltration of Eosinophils and the Release of Allergy-Related Cytokines in BAL of OVA-Induced Allergic Asthmatic Mice
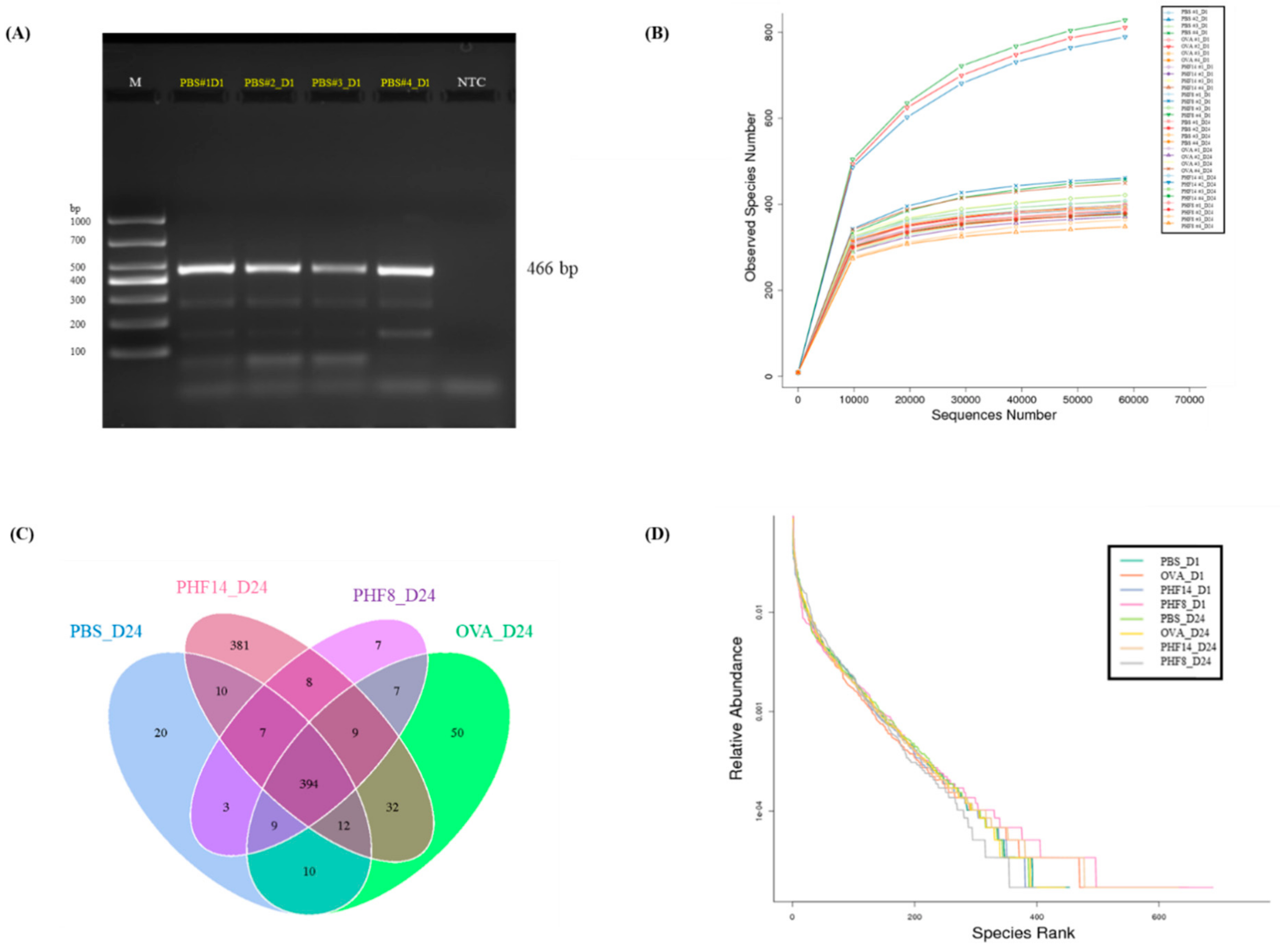
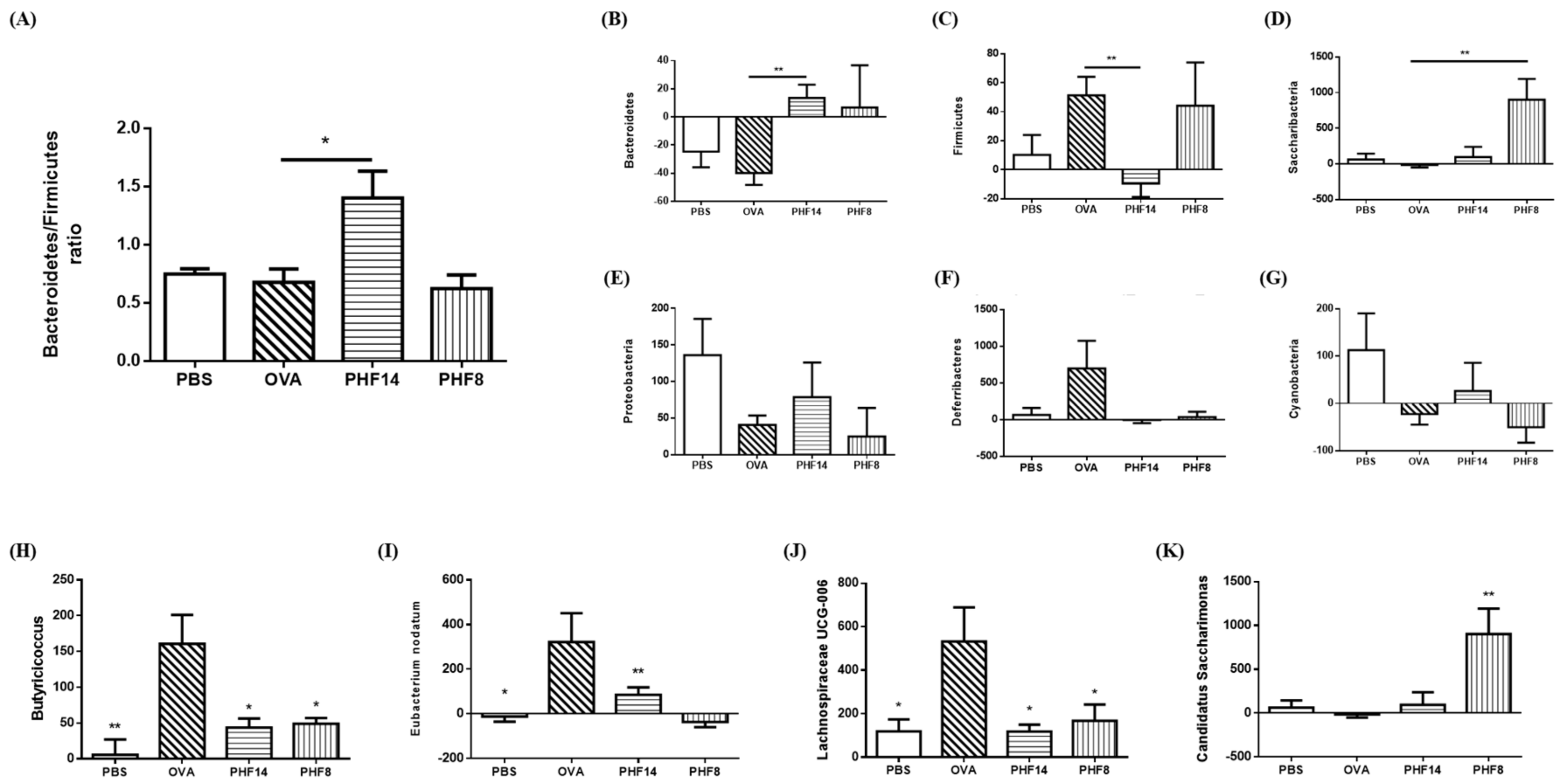
2.3. Pentaherbs Formula Altered the Gut Microbial Diversity
2.4. Effects of Pentaherbs Formula on the Change in Abundance at Phylum and Genus Levels.
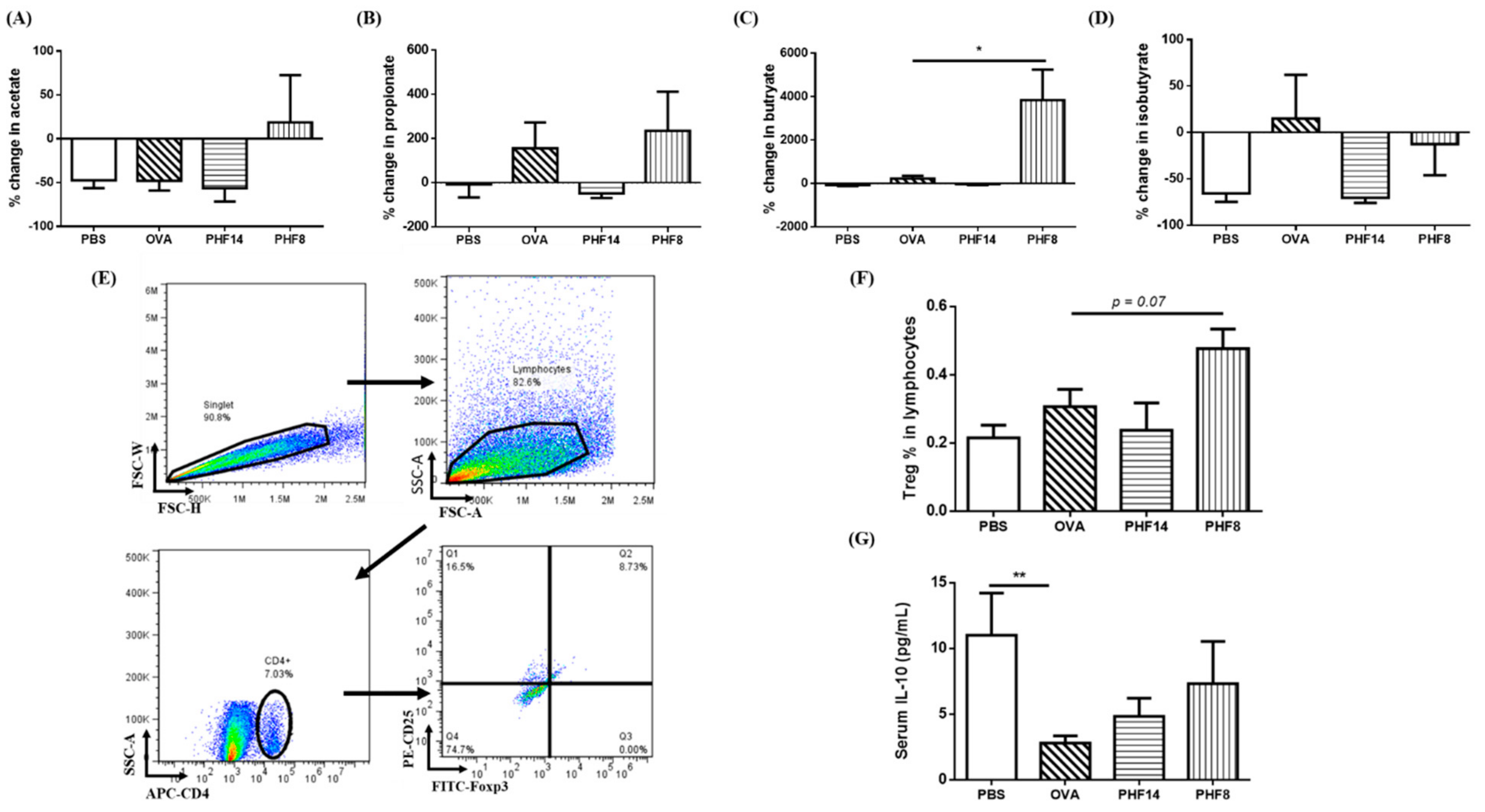
2.5. Changes in Short-Chain Fatty Acids in the Gut, Modulation on Splenic Treg Cells Population and Serum IL-10 Level
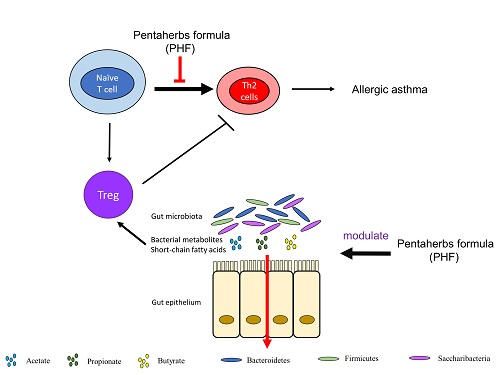
3. Discussion
4. Materials and Methods
4.1. Preparation of Pentaherbs Formula (PHF)
4.2. Animal Experiment
4.3. Measurement of Lung Function by Bronchoconstriction Challenge
4.4. Histological Examination
4.5. Bronchoalveolar Lavage (BAL) Fluid Collection and Immune Cell Determination
4.6. Measurement of OVA-Specific IgE, Total IgE, Cytokines and Chemokines in Serum and BAL Fluid
4.7. Determination of Splenic Treg Cells
4.8. Determination of Gut Microbiota
4.9. Determination of SCFAs in Stools
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Masoli, M.; Fabian, D.; Holt, S.; Holt, S.; Beasley, R. The global burden of asthma: Executive summary of the gina dissemination committee report. Allergy 2004, 59, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Global Initiative for Asthma (GINA). Global Strategy for Asthma Management and Prevention 2018. Available online: http://www.ginasthma.org (accessed on 12 April 2018).
- Holgate, S.T.; Wenzel, S.; Postma, D.S.; Weiss, S.T.; Renz, H.; Sly, P.D. Asthma. Nat. Rev. Dis. Primers 2015, 1, 15025. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, S.B. Safety of beta2-agonists in asthma: Linking mechanisms, meta-analyses and regulatory practice. AAPS J. 2015, 17, 754–757. [Google Scholar] [CrossRef] [PubMed]
- Asamoah, F.; Kakourou, A.; Dhami, S.; Lau, S.; Agache, I.; Muraro, A.; Roberts, G.; Akdis, C.; Bonini, M.; Cavkaytar, O.; et al. Allergen immunotherapy for allergic asthma: A systematic overview of systematic reviews. Clin. Transl. Allergy 2017, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Yu, J.; Oh, M.H.; Zhu, Z. The atopic march: Progression from atopic dermatitis to allergic rhinitis and asthma. Allergy Asthma Immunol. Res. 2011, 3, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Bantz, S.K.; Zhu, Z.; Zheng, T. The atopic march: Progression from atopic dermatitis to allergic rhinitis and asthma. J. Clin. Cell. Immunol. 2014, 5, 202. [Google Scholar] [PubMed]
- Hon, K.L.; Leung, T.F.; Ng, P.C.; Lam, M.C.; Kam, W.Y.; Wong, K.Y.; Lee, K.C.; Sung, Y.T.; Cheng, K.F.; Fok, T.F.; et al. Efficacy and tolerability of a chinese herbal medicine concoction for treatment of atopic dermatitis: A randomized, double-blind, placebo-controlled study. Br. J. Dermatol. 2007, 157, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Hon, K.L.; Lee, V.W.; Leung, T.F.; Lee, K.K.; Chan, A.K.; Fok, T.F.; Leung, P.C. Corticosteroids are not present in a traditional chinese medicine formulation for atopic dermatitis in children. Ann. Acad. Med. Singap. 2006, 35, 759–763. [Google Scholar] [PubMed]
- Leung, T.F.; Wong, K.Y.; Wong, C.K.; Fung, K.P.; Lam, C.W.; Fok, T.F.; Leung, P.C.; Hon, K.L. In vitro and clinical immunomodulatory effects of a novel pentaherbs concoction for atopic dermatitis. Br. J. Dermatol. 2008, 158, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Hon, K.L.; Leung, T.F.; Wong, Y.; Lam, W.K.; Guan, D.Q.; Ma, K.C.; Sung, Y.T.; Fok, T.F.; Leung, P.C. A pentaherbs capsule as a treatment option for atopic dermatitis in children: An open-labeled case series. Am. J. Chin. Med. 2004, 32, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Tsang, M.S.; Jiao, D.; Chan, B.C.; Hon, K.L.; Leung, P.C.; Lau, C.B.; Wong, E.C.; Cheng, L.; Chan, C.K.; Lam, C.W.; et al. Anti-inflammatory activities of pentaherbs formula, berberine, gallic acid and chlorogenic acid in atopic dermatitis-like skin inflammation. Molecules 2016, 21, 519. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.Y.; Park, H.R.; Jeong, H.G.; Kim, S.W. Berberine reduce allergic inflammation in a house dust mite allergic rhinitis mouse model. Rhinology 2015, 53, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.R.; Lee, D.M.; Lee, S.H.; Seong, A.R.; Gin, D.W.; Hwang, J.A.; Park, J.H. Chlorogenic acid suppresses pulmonary eosinophilia, IgE production, and th2-type cytokine production in an ovalbumin-induced allergic asthma: Activation of stat-6 and JNK is inhibited by chlorogenic acid. Int. Immunopharmacol. 2010, 10, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.C.; Hon, K.L.; Leung, P.C.; Sam, S.W.; Fung, K.P.; Lee, M.Y.; Lau, H.Y. Traditional chinese medicine for atopic eczema: Pentaherbs formula suppresses inflammatory mediators release from mast cells. J. Ethnopharmacol. 2008, 120, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.Y.; Hu, S.; Chan, B.C.; Wat, E.C.; Lau, C.B.; Hon, K.L.; Fung, K.P.; Leung, P.C.; Hui, P.C.; Lam, C.W.; et al. Anti-inflammatory and anti-allergic activities of pentaherb formula, moutan cortex (Danpi) and gallic acid. Molecules 2013, 18, 2483–2500. [Google Scholar] [CrossRef] [PubMed]
- West, C.E.; Renz, H.; Jenmalm, M.C.; Kozyrskyj, A.L.; Allen, K.J.; Vuillermin, P.; Prescott, S.L. The gut microbiota and inflammatory noncommunicable diseases: Associations and potentials for gut microbiota therapies. J. Allergy Clin. Immunol. 2015, 135, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, M.A.; Bird, A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2014, 7, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Berthon, B.S.; Macdonald-Wicks, L.K.; Gibson, P.G.; Wood, L.G. Investigation of the association between dietary intake, disease severity and airway inflammation in asthma. Respirology 2013, 18, 447–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor gpr43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory t-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Guo, J.; Tao, W.; Liu, P.; Shang, E.; Zhu, Z.; Fan, X.; Shen, J.; Hua, Y.; Zhu, K.Y.; et al. Gancao-gansui combination impacts gut microbiota diversity and related metabolic functions. J. Ethnopharmacol. 2018, 214, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.H.; Shi, Q.; Han, N.; Zhang, L.; Zhang, Y.Y.; Gao, T.X.; Chen, C.; Li, Y.L. Structural modulation of gut microbiota in rats with allergic bronchial asthma treated with recuperating lung decoction. Biomed. Environ. Sci. 2016, 29, 574–583. [Google Scholar] [PubMed]
- Stott, B.; Lavender, P.; Lehmann, S.; Pennino, D.; Durham, S.; Schmidt-Weber, C.B. Human il-31 is induced by il-4 and promotes th2-driven inflammation. J. Allergy Clin. Immunol. 2013, 132, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Leung, K.M.; Qiu, H.N.; Chow, J.Y.; Choi, A.O.; Lam, C.W. Activation of eosinophils interacting with dermal fibroblasts by pruritogenic cytokine il-31 and alarmin il-33: Implications in atopic dermatitis. PLoS ONE 2012, 7, e29815. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhu, L.; Tao, Y.; Sun, H.X.; Li, Y.; Wang, P.; Hou, Y.; Zhao, Y.; Zhang, X.; Zhang, L.; et al. Characterization and allergic role of il-33-induced neutrophil polarization. Cell. Mol. Immunol. 2018, 15, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Berger, A. Th1 and th2 responses: What are they? BMJ 2000, 321, 424. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Muller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the german biodiversity exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Kong, H.; Zeng, X.; Guo, L.; Sun, X.; He, S. Subsets of regulatory t cells and their roles in allergy. J. Transl. Med. 2014, 12, 125. [Google Scholar] [CrossRef] [PubMed]
- Hawrylowicz, C.M.; O'Garra, A. Potential role of interleukin-10-secreting regulatory t cells in allergy and asthma. Nat. Rev. Immunol. 2005, 5, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Riiser, A. The human microbiome, asthma, and allergy. Allergy Asthma Clin. Immunol. 2015, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosnjak, B.; Stelzmueller, B.; Erb, K.J.; Epstein, M.M. Treatment of allergic asthma: Modulation of th2 cells and their responses. Respir. Res. 2011, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wong, C.K.; Cai, Z.; Jiao, D.; Chu, M.; Lam, C.W. Amelioration of allergic airway inflammation in mice by regulatory il-35 through dampening inflammatory dendritic cells. Allergy 2015, 70, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Bjorksten, B.; Engstrand, L.; Jenmalm, M.C. Low gut microbiota diversity in early infancy precedes asthma at school age. Clin. Exp. Allergy 2014, 44, 842–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Bjorksten, B.; Engstrand, L.; Jenmalm, M.C. Low diversity of the gut microbiota in infants with atopic eczema. J. Allergy Clin. Immunol. 2012, 129, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.; Raison, C.L.; Lowry, C.A. Microbial 'old friends', immunoregulation and socioeconomic status. Clin. Exp. Immunol. 2014, 177, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide utilization by gut bacteria: Potential for new insights from genomic analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gao, X. Role of dendritic cells: A step forward for the hygiene hypothesis. Cell. Mol. Immunol. 2011, 8, 12–18. [Google Scholar] [CrossRef] [PubMed]
- He, X.; McLean, J.S.; Edlund, A.; Yooseph, S.; Hall, A.P.; Liu, S.Y.; Dorrestein, P.C.; Esquenazi, E.; Hunter, R.C.; Cheng, G.; et al. Cultivation of a human-associated tm7 phylotype reveals a reduced genome and epibiotic parasitic lifestyle. Proc. Natl. Acad. Sci. USA 2015, 112, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Paez, A.; Del Pulgar, E.M.G.; Kjolbaek, L.; Brahe, L.K.; Astrup, A.; Larsen, L.; Sanz, Y. Impact of dietary fiber and fat on gut microbiota re-modeling and metabolic health. Trends Food Sci. Technol. 2016, 57, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Gong, F.; Zhu, H.Y.; Zhu, J.; Dong, Q.J.; Huang, X.; Jiang, D.J. Circulating CXCR5(+)CD4(+) T cells participate in the IgE accumulation in allergic asthma. Immunol. Lett. 2018, 197, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Winstead, C.J. Follicular helper T cell-mediated mucosal barrier maintenance. Immunol. Lett. 2014, 162, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Kato, L.M.; Kawamoto, S.; Maruya, M.; Fagarasan, S.; Gut, T.F. IgA: Key players for regulation of bacterial communities and immune homeostasis. Immunol. Cell Biol. 2014, 92, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M. Tfr cells and IgA join forces to diversify the microbiota. Immunity 2014, 41, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Chen, R.; Cai, Z.; Yu, L.; Fei, Y.; Weng, L.; Wang, J.; Ge, X.; Zhu, T.; Wang, J.; et al. Salmeterol attenuates the inflammatory response in asthma and decreases the pro-inflammatory cytokine secretion of dendritic cells. Cell. Mol. Immunol. 2012, 9, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16s rrna diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Kim, K.H. Parallel analysis of volatile fatty acids, indole, skatole, phenol, and trimethylamine from waste-related source environments. J. Chromatogr. A 2013, 1314, 241–248. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of Pentaherbs formula and the compound gallic acid, berberine and chlorogenic acid are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PBS | OVA | PHF14 | PHF8 | Dex | |
|---|---|---|---|---|---|
| Smooth muscle hypertrophy | - | ++ | - | ++ | + |
| Goblet cell hyperplasia | - | +++ | + | - | + |
| Eosinophil infiltration | - | +++ | + | +++ | - |
| Day 1 | Day 24 | |||||||
|---|---|---|---|---|---|---|---|---|
| PBS | OVA | PHF14 | PHF8 | PBS | OVA | PHF14 | PHF8 | |
| Mean of total OTU observed | 413 | 490 | 398 | 525 | 394 | 400 | 485 | 370 |
| Mean of Shannon diversity index | 6.49 | 6.29 | 6.30 | 6.21 | 6.36 | 6.32 | 6.44 | 6.48 |
| Mean of Simpson index | 0.975 | 0.968 | 0.965 | 0.943 | 0.970 | 0.964 | 0.972 | 0.979 |
| Taxonomy | Mean of Relative Abundance (%) | p-Value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Day 1 | Day 24 | ||||||||||||
| Phylum | Genus | PBS | OVA | PHF 14 | PHF8 | PBS | OVA | PHF 14 | PHF8 | PBS | OVA | PHF 14 | PHF 8 |
| Bacteroidetes | Alistipes | 9.26 | 8.24 | 7.66 | 11.2 | 8.50 | 8.10 | 7.59 | 5.75 | 0.17 | 0.95 | 0.99 | 0.19 |
| Bacteroides | 4.48 | 1.46 | 1.26 | 1.54 | 3.29 | 4.57 | 3.85 | 2.26 | 0.78 | 0.09 | 0.13 | 0.64 | |
| Odoribacter | 1.47 | 1.96 | 0.86 | 1.53 | 1.24 | 0.85 | 0.49 | 0.80 | 0.38 | 0.22 | 0.13 | 0.03 | |
| Rikenella | 1.62 | 2.17 | 1.54 | 1.11 | 3.23 | 0.96 | 0.59 | 0.39 | 0.29 | 0.09 | 0.02 | 0.061 | |
| Rikenellaceae_RC9_gut_group | 1.10 | 0.86 | 0.60 | 0.88 | 0.68 | 1.48 | 0.46 | 0.18 | 0.31 | 0.39 | 0.42 | 0.08 | |
| Deferribacteres | Mucispirillum | 0.13 | 0.07 | 0.80 | 0.34 | 0.19 | 0.30 | 0.11 | 0.24 | 0.58 | 0.04 | 0.39 | 0.59 |
| Firmicutes | [Eubacterium]_xylanophilum_group | 1.21 | 0.50 | 0.67 | 0.37 | 0.62 | 1.09 | 0.52 | 0.76 | 0.54 | 0.01 | 0.73 | 0.25 |
| Lachnospiraceae_NK4A136_group | 14.9 | 12.5 | 13.2 | 11.2 | 20.0 | 19.8 | 10.1 | 20.5 | 0.16 | 0.32 | 0.56 | 0.17 | |
| Lactobacillus | 2.16 | 2.40 | 8.22 | 12.3 | 3.16 | 4.30 | 2.13 | 1.87 | 0.31 | 0.35 | 0.17 | 0.30 | |
| Ruminiclostridium | 1.83 | 1.14 | 1.13 | 1.30 | 0.65 | 2.01 | 0.82 | 1.00 | 0.04 | 0.26 | 0.29 | 0.65 | |
| Ruminiclostridium_9 | 1.84 | 1.36 | 1.47 | 1.70 | 1.21 | 1.64 | 1.34 | 2.03 | 0.08 | 0.58 | 0.57 | 0.51 | |
| Ruminococcaceae_UCG-014 | 5.49 | 2.51 | 5.08 | 5.55 | 7.63 | 3.99 | 6.80 | 4.89 | 0.65 | 0.40 | 0.68 | 0.85 | |
| unidentified_Lachnospiraceae | 1.83 | 1.03 | 1.76 | 1.78 | 1.89 | 2.73 | 2.20 | 3.43 | 0.79 | 0.07 | 0.52 | 0.13 | |
| unidentified_Ruminococcaceae | 0.98 | 0.89 | 0.81 | 0.70 | 0.45 | 0.84 | 0.60 | 1.70 | 0.21 | 0.86 | 0.13 | 0.14 | |
| Proteobacteria | Desulfovibrio | 0.59 | 0.81 | 0.76 | 0.51 | 2.74 | 1.58 | 2.06 | 2.02 | 0.11 | 0.32 | 0.11 | 0.06 |
| Saccharibacteria | Candidatus_Saccharimonas | 1.00 | 2.60 | 1.96 | 0.26 | 0.98 | 1.55 | 1.66 | 1.64 | 0.94 | 0.35 | 0.70 | 0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsang, M.S.-M.; Cheng, S.-W.; Zhu, J.; Atli, K.; Chan, B.C.-L.; Liu, D.; Chan, H.Y.-T.; Sun, X.; Chu, I.M.-T.; Hon, K.-L.; et al. Anti-Inflammatory Activities of Pentaherbs formula and Its Influence on Gut Microbiota in Allergic Asthma. Molecules 2018, 23, 2776. https://doi.org/10.3390/molecules23112776
Tsang MS-M, Cheng S-W, Zhu J, Atli K, Chan BC-L, Liu D, Chan HY-T, Sun X, Chu IM-T, Hon K-L, et al. Anti-Inflammatory Activities of Pentaherbs formula and Its Influence on Gut Microbiota in Allergic Asthma. Molecules. 2018; 23(11):2776. https://doi.org/10.3390/molecules23112776
Chicago/Turabian StyleTsang, Miranda Sin-Man, Sau-Wan Cheng, Jing Zhu, Karam Atli, Ben Chung-Lap Chan, Dehua Liu, Helen Yau-Tsz Chan, Xiaoyu Sun, Ida Miu-Ting Chu, Kam-Lun Hon, and et al. 2018. "Anti-Inflammatory Activities of Pentaherbs formula and Its Influence on Gut Microbiota in Allergic Asthma" Molecules 23, no. 11: 2776. https://doi.org/10.3390/molecules23112776
APA StyleTsang, M. S. -M., Cheng, S. -W., Zhu, J., Atli, K., Chan, B. C. -L., Liu, D., Chan, H. Y. -T., Sun, X., Chu, I. M. -T., Hon, K. -L., Lam, C. W. -K., Shaw, P. -C., Leung, P. -C., & Wong, C. -K. (2018). Anti-Inflammatory Activities of Pentaherbs formula and Its Influence on Gut Microbiota in Allergic Asthma. Molecules, 23(11), 2776. https://doi.org/10.3390/molecules23112776