
Characterization of Arginine Catabolism by Lactic Acid Bacteria Isolated from Kimchi

Abstract
:1. Introduction
2. Results
2.1. Arginine Catabolic Activity of Lactic Acid Bacteria
2.2. The ADI System of Lactic Acid Bacteria
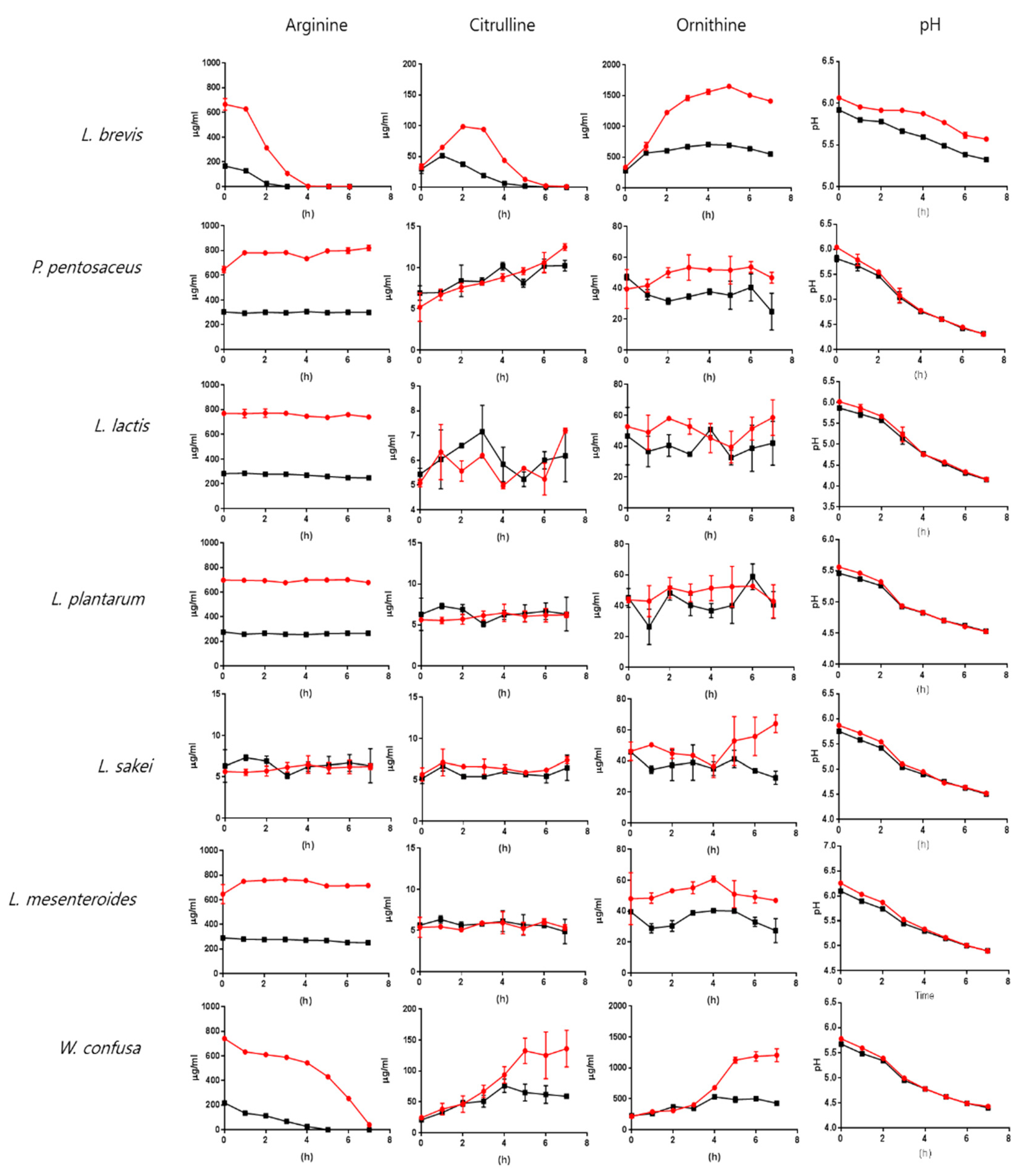
2.3. Arginine, Citrulline, and Ornithine Production by Lactic Acid Bacteria
2.4. Effect of Citrulline and Ornithine on the Growth of Lactic Acid Bacteria
2.5. Arginine Catabolism in Kimchi
3. Discussion
4. Materials and Methods
4.1. Medium and Bacterial Culture Conditions
4.2. Sample Preparation and Analysis
4.3. Protein Sequence Analysis
4.4. Statistical Analysis
4.5. Comparison and Analysis of Metagenomics Data
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| aa | Amino acid |
| ADI | Arginine deiminase |
| CK | Carbamate kinase |
| HICA | 2-Hydroxyisocaproic acid |
| MRM | Multiple-reaction monitoring |
| MRS | De Man, Rogosa, and Sharpe |
| OTC | Ornithine transcarbamylase |
References
- Patra, J.K.; Das, G.; Paramithiotis, S.; Shin, H.S. Kimchi and Other Widely Consumed Traditional Fermented Foods of Korea: A Review. Front. Microbiol. 2016, 7, 1493. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Bae, J.H. Fermented food intake is associated with a reduced likelihood of atopic dermatitis in an adult population (Korean National Health and Nutrition Examination Survey 2012-2013). Nutr. Res. 2016, 36, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Lee, D.; Yang, C.; Jeon, J.; Kim, J.; Han, H. Microbial population dynamics of kimchi, a fermented cabbage product. FEMS Microbiol. Lett. 2006, 257, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Lee, S.H.; Jung, J.Y.; Choi, E.J.; Jeon, C.O. Microbial succession and metabolite changes during long-term storage of Kimchi. J. Food Sci. 2013, 78, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Park, B.; Hwang, H.; Chang, J.Y.; Hong, S.W.; Lee, S.H.; Jung, M.Y.; Sohn, S.O.; Park, H.W.; Lee, J.H. Identification of 2-hydroxyisocaproic acid production in lactic acid bacteria and evaluation of microbial dynamics during kimchi ripening. Sci. Rep. 2017, 7, 10904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Jung, J.Y.; Jeon, C.O. Source Tracking and Succession of Kimchi Lactic Acid Bacteria during Fermentation. J. Food Sci. 2015, 80, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.K.; Cho, M.S.; Park, D.S. Red pepper powder is a crucial factor that influences the ontogeny of Weissella cibaria during kimchi fermentation. Sci. Rep. 2016, 6, 28232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadsah, L.S.; Min, S.G.; Han, S.K.; Hong, Y.; Kim, H.Y. Effect of Low Salt Concentrations on Microbial Changes During Kimchi Fermentation Monitored by PCR-DGGE and Their Sensory Acceptance. J. Microbiol. Biotechnol. 2015, 25, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Seo, H.Y.; Park, W.S.; Jeon, C.O. Effects of Leuconostoc mesenteroides starter cultures on microbial communities and metabolites during kimchi fermentation. Int. J. Food Microbiol. 2012, 153, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.-Y.; Kim, G.-H. Changes in vitamin U and amino acid levels of Korean Chinese cabbages during Kimchi fermentation. Korean J. Food Preserv. 2005, 12, 411–416. [Google Scholar]
- Mi-Soon, J.; Hyeon Gyu, N.; Kyoung-Chun, S.; Ki-Ho, N.; Hee-Yeon, P. Changes in the taste compounds of Kimchi with seafood added during its fermentation. Korean J. Food Preserv. 2013, 20, 404–418. [Google Scholar]
- Choi, Y.-J.; Lee, H.-W.; Yang, J.-H.; Hong, S.-W.; Park, S.-H.; Lee, M.-A. Changes in quality properties of kimchi based on the nitrogen content of fermented anchovy sauce, Myeolchi Aekjeot, during fermentation. Food Sci. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Zhang, J.; Zhou, J.; Zhou, Z.; Li, T.; Lu, L.; Zeng, W.; Du, G.; Chen, J. Accumulation of Citrulline by Microbial Arginine Metabolism during Alcoholic Fermentation of Soy Sauce. J. Agric. Food Chem. 2018, 66, 2108–2113. [Google Scholar] [CrossRef] [PubMed]
- Araque, I.; Bordons, A.; Reguant, C. Effect of ethanol and low pH on citrulline and ornithine excretion and arc gene expression by strains of Lactobacillus brevis and Pediococcus pentosaceus. Food Microbiol. 2013, 33, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Araque, I.; Reguant, C.; Rozes, N.; Bordons, A. Influence of wine-like conditions on arginine utilization by lactic acid bacteria. Int. Microbiol. 2011, 14, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Novák, L.; Zubáčová, Z.; Karnkowska, A.; Kolisko, M.; Hroudová, M.; Stairs, C.W.; Simpson, A.G.B.; Keeling, P.J.; Roger, A.J.; Čepička, I.; et al. Arginine deiminase pathway enzymes: Evolutionary history in metamonads and other eukaryotes. BMC Evol. Biol. 2016, 16, 197. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, A.; Revsbech, N.P. An NH4+ biosensor based on ammonia-oxidizing bacteria for use under anoxic conditions. Sens. Actuators B 2005, 105, 412–418. [Google Scholar] [CrossRef]
- Palková, Z.; Janderová, B.; Gabriel, J.í.; Zikánová, B.; Pospíŝek, M.; Forstová, J. Ammonia mediates communication between yeast colonies. Nature 1997, 390, 532. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Kim, B.M.; Lee, H.J.; Jang, G.J.; Song, S.H.; Lee, J.I.; Lee, S.B.; Shim, J.M.; Lee, K.W.; Kim, J.H.; et al. Effects of Different Salt Treatments on the Fermentation Metabolites and Bacterial Profiles of Kimchi. J. Food Sci. 2017, 82, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Hye-Hyun, L.; Gun-Hee, K. Changes in the Levels of γ-Aminobutyric Acid and Free Amino Acids during Kimchi Fermentation. Korean J. Food Cook. Sci. 2013, 29, 671–677. [Google Scholar] [Green Version]
- Champomier Verges, M.C.; Zuniga, M.; Morel-Deville, F.; Perez-Martinez, G.; Zagorec, M.; Ehrlich, S.D. Relationships between arginine degradation, pH and survival in Lactobacillus sakei. FEMS Microbiol. Lett. 1999, 180, 297–304. [Google Scholar] [CrossRef]
- Majsnerowska, M.; Noens, E.E.E.; Lolkema, J.S. Arginine and Citrulline Catabolic Pathways encoded by the arc gene cluster of Lactobacillus brevis ATCC367. J. Bacteriol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Noens, E.E.E.; Lolkema, J.S. Convergent evolution of the arginine deiminase pathway: The ArcD and ArcE arginine/ornithine exchangers. Microbiol. Open 2017, 6, e00412. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.Y.; Lee, J.; Park, B.; Hwang, H.; Sohn, S.O.; Lee, S.H.; Lim, H.I.; Park, H.W.; Lee, J.H. Applicability of a colorimetric method for evaluation of lactic acid bacteria with probiotic properties. Food Microbiol. 2017, 64, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.D.; Gardner, C.D. Composition, Stability, and Bioavailability of Garlic Products Being Used in a Clinical Trial. J. Agric. Food Chem. 2005, 53, 6254–6261. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | ADI | OTC | CK | |||
|---|---|---|---|---|---|---|
| Protein | Gene | Protein | Gene | Protein | Gene | |
| L. brevis ATCC 367/JCM 1170 | Q03NY8 | arcA, (LVIS_20270 | Q03NY9 | arcB, (LVIS_2026) | Q03NG5, Q03NZ2 | LVIS_2207, LVIS_2023 |
| P. pentosaceus ATCC 25754 | Q03DS2 | arcA, (PEPE_1629) | Q03DS0 | arcB, (PEPE_1631) | Q03DS1, Q03HM6 | PEPE_1630, PEPE_0192 |
| L. plantarum WCFS1 | Q6U3A0 | arcA | O08322 | argF, (lp_0532) | Q6U398 | arcC |
| L. lactis | - | - | A0A1B2A277 | BCR17_06740 | - | - |
| L. sakei 23K | Q38YQ6 | arcA, (LCA_0370) | Q38YQ5 | arcB, LCA_0371 | Q38YQ4 | arcC, (LCA_0372) |
| L. mesenteroides ATCC 8293 | Q03XZ3 | arcA, (LEUM_0821) | Q03W72 | arcB, (LEUM_1457) | - | - |
| W. confuse LBAE C39-2 | H1X7H9 | arcA, (WEISS39_05260) | H1X7H8 | arcB, (WEISS39_05255) | H1X7H6 | arcC3, (WEISSC39_05245) |
| Bacterial Strain | ADI | OTC | CK |
|---|---|---|---|
| Lactobacillus brevis | ● | ● | ● |
| Lactobacillus curvatus | |||
| Lactobacillus graminis | |||
| Lactobacillus mali | ● | ||
| Lactobacillus parabrevis | ● | ● | |
| Lactobacillus parabuchneri | ● | ● | ● |
| Lactobacillus paracasei | |||
| Lactobacillus paraplantarum | ● | ||
| Lactobacillus pentosus | ● | ||
| Lactobacillus plantarum | ● | ● | ● |
| Lactobacillus sakei | ● | ● | ● |
| Lactococcus lactis | ● | ● | ● |
| Leuconostoc carnosum | |||
| Leuconostoc citreum | ● | ||
| Leuconostoc gelidum subsp. gasicomitatum | ● | ||
| Leuconostoc kimchii | ● | ||
| Leuconostoc lactis | ● | ||
| Leuconostoc mesenteroides | ● | ● | |
| Leuconostoc pseudomesenteroides | ● | ||
| Pediococcus pentosaceus | ● | ● | ● |
| Streptococcus gallolyticus | ● | ● | |
| Weissella cibaria | ● | ● | ● |
| Weissella confusa | ● | ● | ● |
| Weissella hellenica | |||
| Weissella paramesenteroides | |||
| Weissella soli | |||
| Weissella viridescens | ● | ||
| Weissella koreensis | ● | ● | ● |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, H.; Lee, J.-H. Characterization of Arginine Catabolism by Lactic Acid Bacteria Isolated from Kimchi. Molecules 2018, 23, 3049. https://doi.org/10.3390/molecules23113049
Hwang H, Lee J-H. Characterization of Arginine Catabolism by Lactic Acid Bacteria Isolated from Kimchi. Molecules. 2018; 23(11):3049. https://doi.org/10.3390/molecules23113049
Chicago/Turabian StyleHwang, Hyelyeon, and Jong-Hee Lee. 2018. "Characterization of Arginine Catabolism by Lactic Acid Bacteria Isolated from Kimchi" Molecules 23, no. 11: 3049. https://doi.org/10.3390/molecules23113049
APA StyleHwang, H., & Lee, J.-H. (2018). Characterization of Arginine Catabolism by Lactic Acid Bacteria Isolated from Kimchi. Molecules, 23(11), 3049. https://doi.org/10.3390/molecules23113049