Valorization of Brewers’ Spent Grain for the Production of Lipids by Oleaginous Yeast




Abstract
:
1. Introduction
2. Results and Discussion
2.1. Characterization and Enzymatic Hydrolysis of BSG
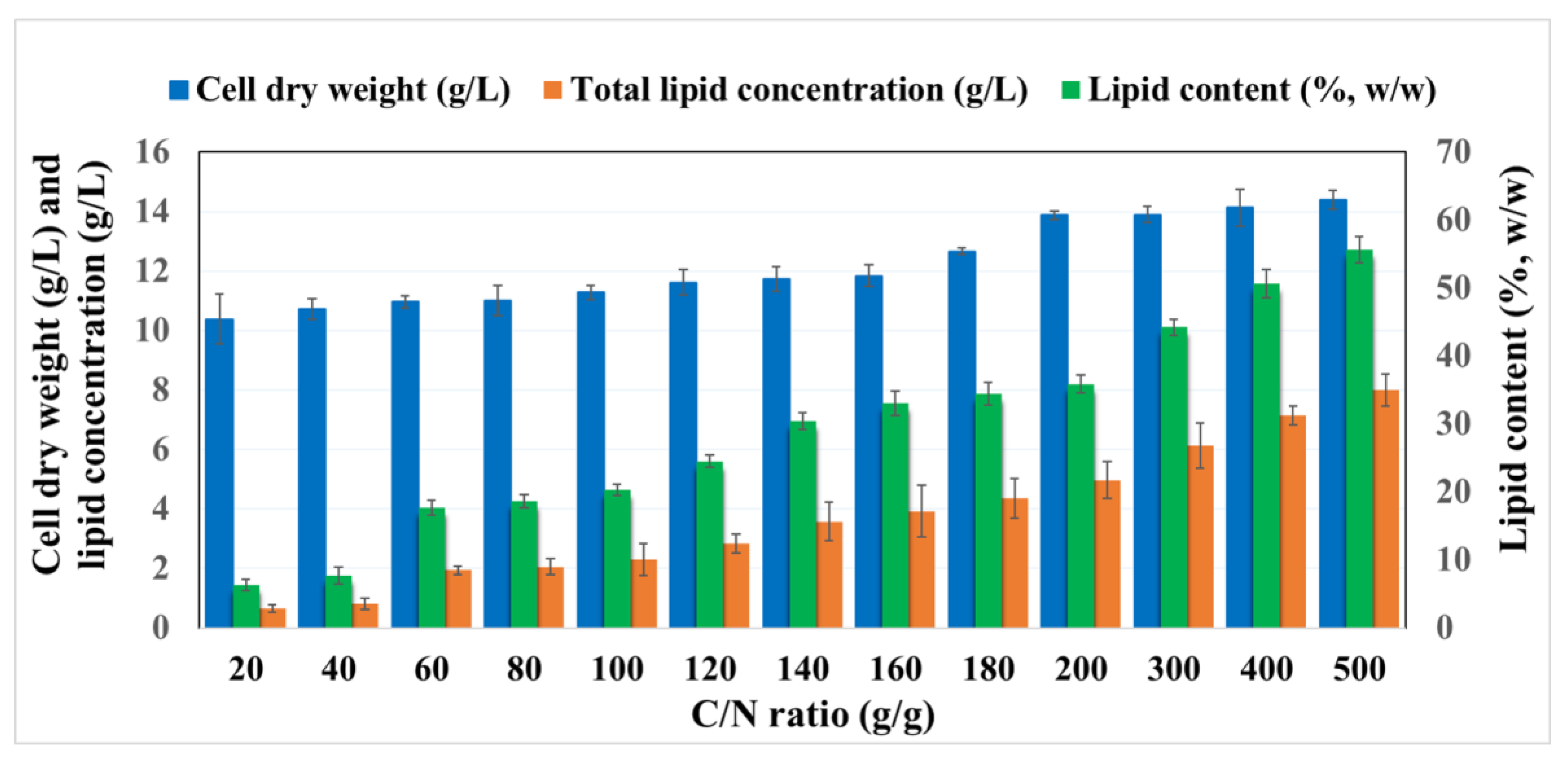
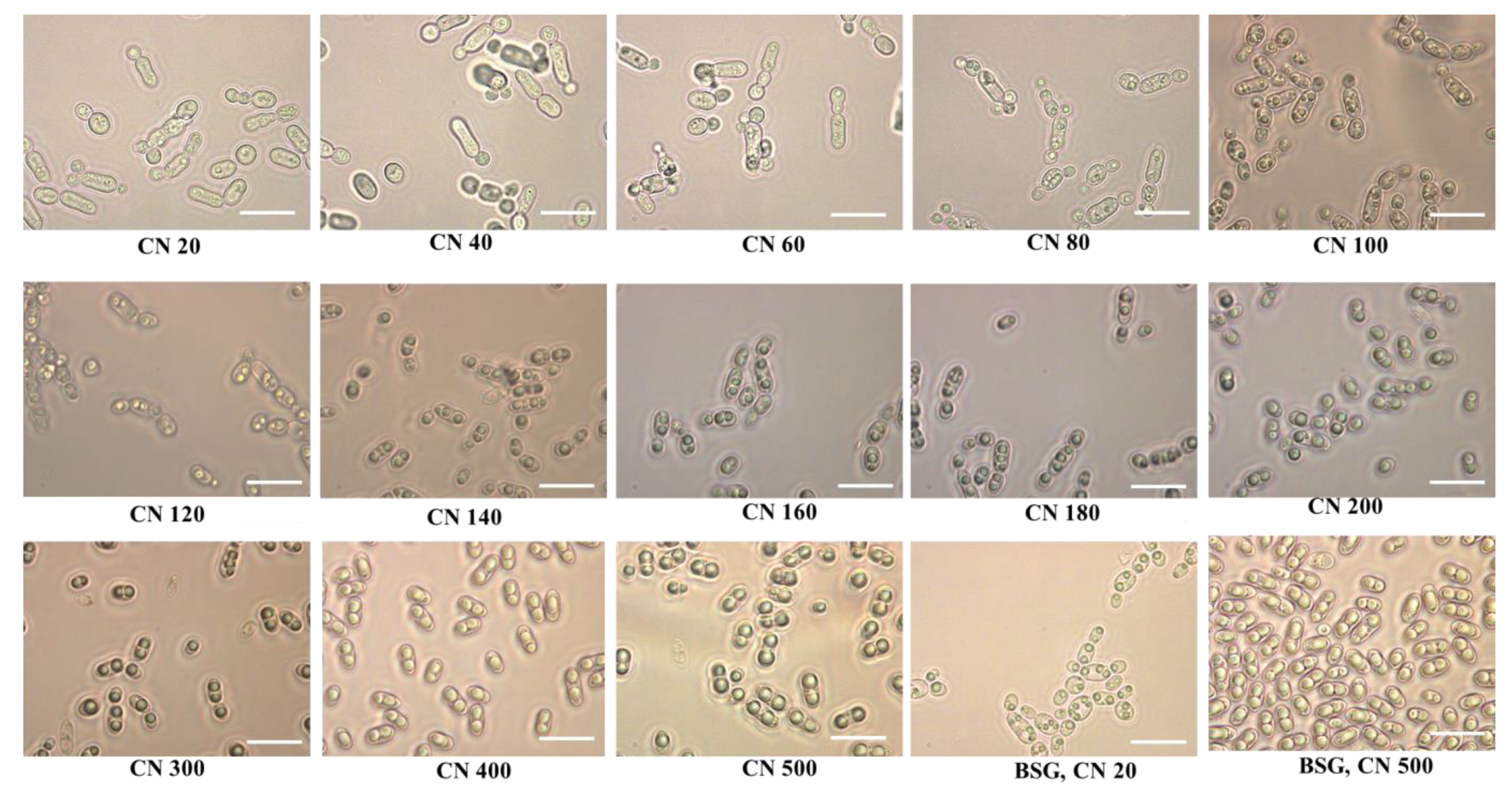
2.2. Optimization of the C/N (g/g) Ratio for Lipid Accumulation of R. Toruloides
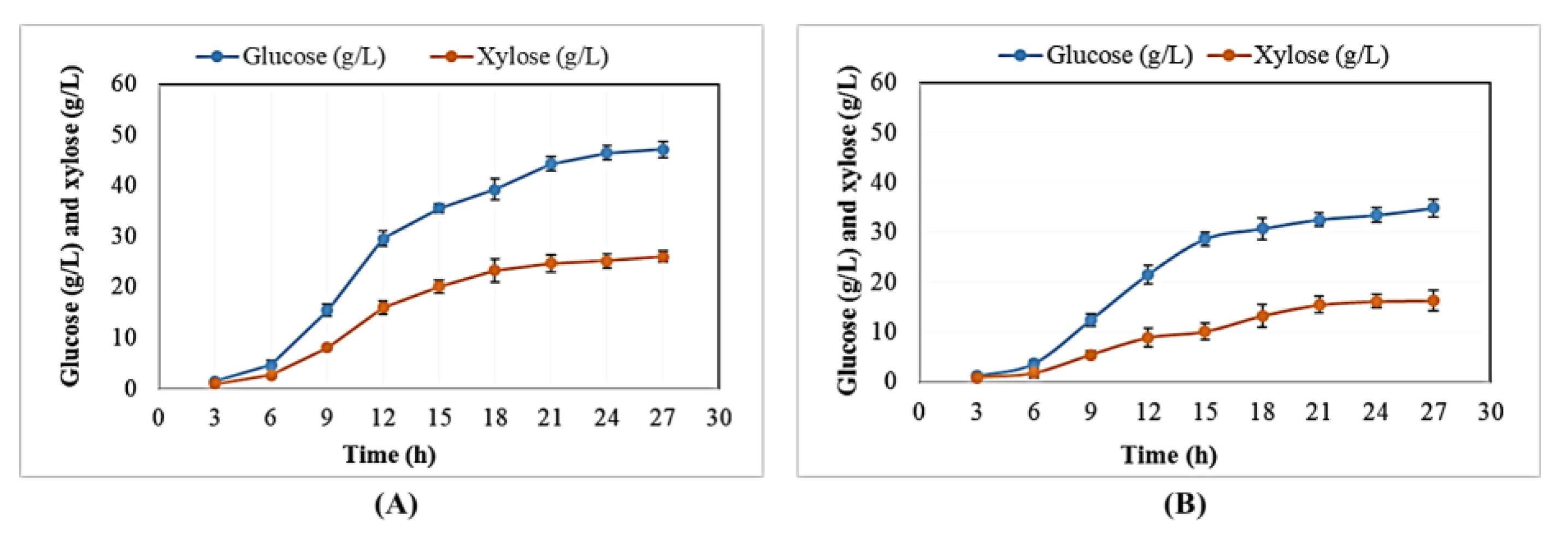
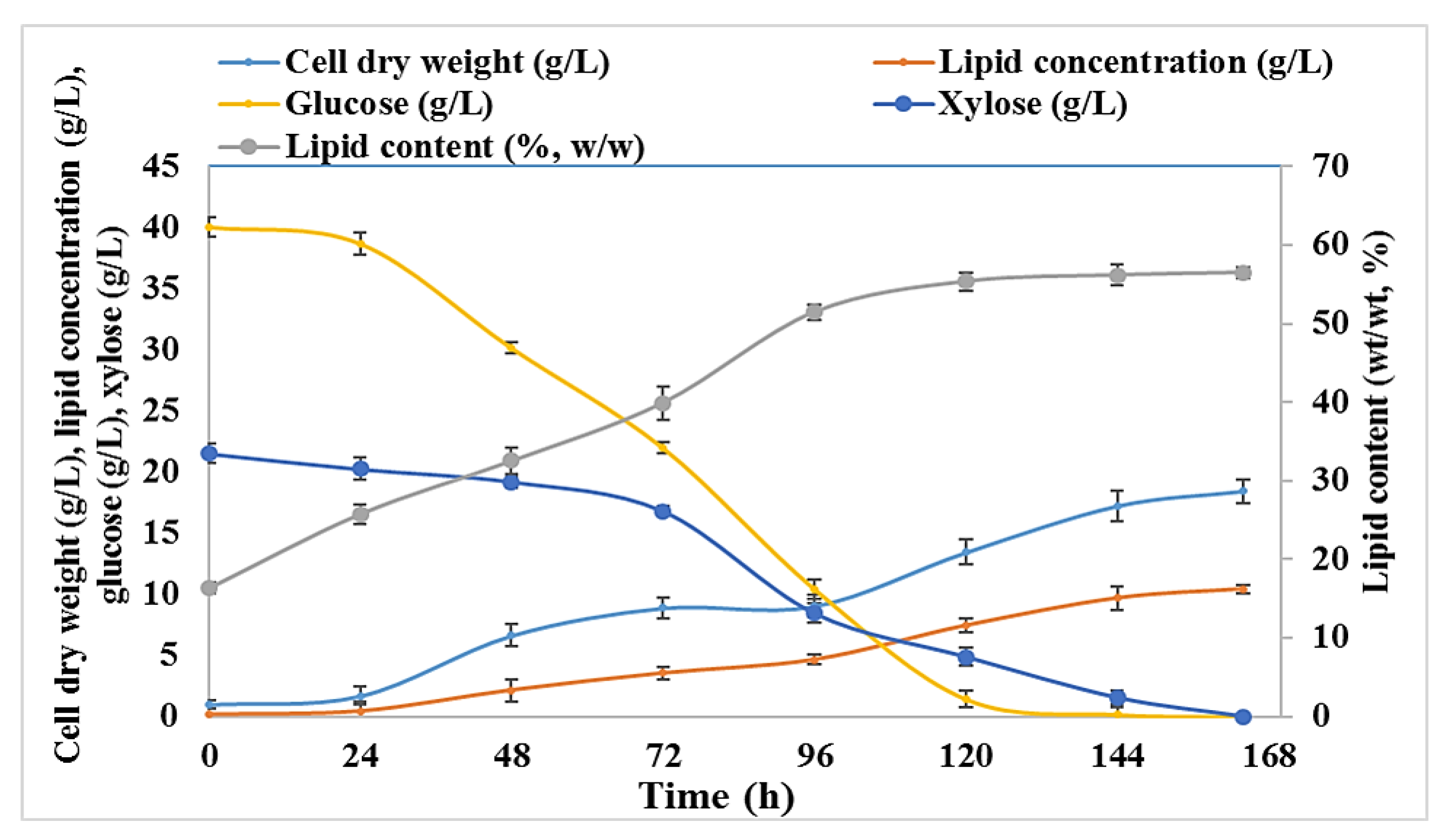
2.3. Time Course Experiments of R. toruloides Grown in BSG Hydrolysates
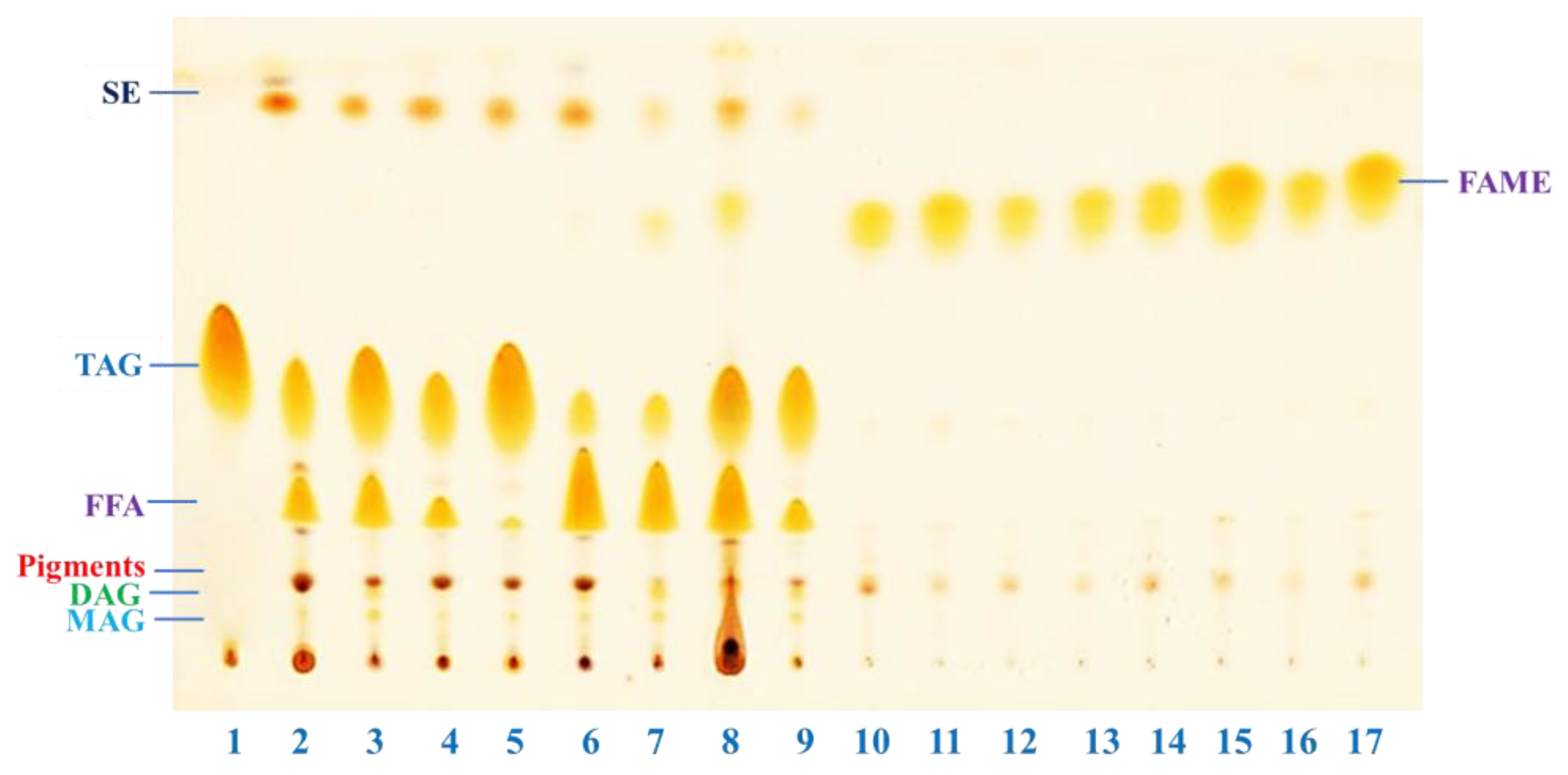
2.4. Estimation of Extracted Fatty Acids and Transesterified Products by Thin Layer Chromatography
2.5. Fatty Acid Analysis of the Lipids Obtained from R. toruloides
3. Materials and Methods
3.1. Collection of BSG and Characterization
3.2. Pretreatment of BSG and Enzymatic Hydrolysis
3.3. Yeast Strain and Growth Condition
3.4. Optimization of the C/N Ratio (g/g) for the Lipid Fermentation by Oleaginous Yeast
3.5. Batch Cultivation Experiments
3.6. Analytical Methods
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMP | Adenosine monophosphate |
| ASTM | American Society for Testing and Materials |
| BSG | Brewers’ spent grain |
| CCR | Carbon catabolite repression |
| CDW | Cell dry weight |
| CFPP | Cold filter plugging point |
| C/N | Carbon/nitrogen |
| CN | Cetane number |
| DAG | Diacylglycerols |
| FAMEs | Fatty acid methyl esters |
| FFA | Free fatty acids |
| h | Hour(s) |
| LCSF | Long chain saturation factor |
| LD | Lipid droplets |
| MAG | Monoacylglycerols |
| MUFAs | Mono-unsaturated fatty acids |
| NREL | National Renewable Energy Laboratory |
| OD | Optical density |
| PUFAs | Poly-unsaturated fatty acids |
| SFAs | Saturated fatty acids |
| TAGs | Triacylglycerols |
| TLC | Thin layer chromatography |
| YPD | Yeast extract peptone dextrose |
| YNB | Yeast nitrogen Base |
References
- Amorim, W.S.; Valduga, I.B.; Ribeiro, J.M.P.; Williamson, V.G.; Krauser, G.E.; Magtoto, M K.; de Andrade Guerra, J.B.S.O. The nexus between water, energy, and food in the context of the global risks: An analysis of the interactions between food, water, and energy security. Environ. Impact Assess. Rev. 2018, 72, 1–11. [Google Scholar] [CrossRef]
- Lin, L.; Gettelman, A.; Fu, Q.; Xu, Y. Simulated differences in 21st century aridity due to different scenarios of greenhouse gases and aerosols. Clim. Chang. 2018, 146, 407–422. [Google Scholar] [CrossRef]
- Jenniches, S. Assessing the regional economic impacts of renewable energy sources—A literature review. Renew. Sustain. Energy Rev. 2018, 93, 35–51. [Google Scholar] [CrossRef]
- Saidur, R.; Boroumandjazi, G.; Mekhilef, S.; Mohammed, H.A. A review on exergy analysis of biomass based fuels. Renew. Sustain. Energy Rev. 2012, 16, 1217–1222. [Google Scholar] [CrossRef]
- Tenenbaum, D.J. Food vs. fuel diversion of crops could cause more hunger. Environ. Health Perspect. 2008, 116, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Mandal, B.K. A comprehensive review of biodiesel as an alternative fuel for compression ignition engine. Renew. Sustain. Energy Rev. 2016, 57, 799–821. [Google Scholar] [CrossRef]
- Knothe, G. Biodiesel and renewable diesel: A comparison. Prog. Energy Combust. Sci. 2010, 36, 364–373. [Google Scholar] [CrossRef]
- Li, Q.; Du, W.; Liu, D. Perspectives of microbial oils for biodiesel production. Appl. Microbiol. Biotechnol. 2008, 80, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Leiva-Candia, D.E.; Pinzi, S.; Redel-Macías, M.D.; Koutinas, A.; Webb, C.; Dorado, M.P. The potential for agro-industrial waste utilization using oleaginous yeast for the production of biodiesel. Fuel 2014, 123, 33–42. [Google Scholar] [CrossRef]
- Gancedo, J.M. Yeast Carbon Catabolite Repression. Microbiol. Mol. Biol. Rev. 1998, 62, 334–361. [Google Scholar] [PubMed]
- Stülke, J.; Hillen, W. Carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 1999, 2, 195–201. [Google Scholar] [CrossRef]
- Kim, J.-H.; Block, D.E.; Mills, D.A. Simultaneous consumption of pentose and hexose sugars: An optimal microbial phenotype for efficient fermentation of lignocellulosic biomass. Appl. Microbiol. Biotechnol. 2010, 88, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, D. Exploitation of genus Rhodosporidium for microbial lipid production. World J. Microbiol. Biotechnol. 2017, 33, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Dragone, G.; Roberto, I.C. Brewers’ spent grain: Generation, characteristics and potential applications. J. Cereal Sci. 2006, 43, 1–14. [Google Scholar] [CrossRef]
- Xiros, C.; Christakopoulos, P. Biotechnological potential of brewers spent grain and its recent applications. Waste Biomass Valori. 2012, 3, 213–232. [Google Scholar] [CrossRef]
- Mussatto, S.I. Brewer’s spent grain: A valuable feedstock for industrial applications. J. Sci. Food Agric. 2014, 94, 1264–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliyu, S.; Bala, M. Brewer’s spent grain: A review of its potentials and applications. Afr. J. Biotechnol. 2011, 10, 324–331. [Google Scholar]
- Fărcaş, A.; Tofană, M.; Socaci, S.; Mudura, E.; Scrob, S.; Salanţă, L.; Mureşan, V. Brewers ’ spent grain—A new potential ingredient for functional foods. J. Agroaliment. Process. Technol. 2014, 20, 137–141. [Google Scholar]
- Santos, M.; Jiménez, J.; Bartolomé, B.; Gómez-Cordovés, C.; del Nozal, M.J. Variability of brewer’s spent grain within a brewery. Food Chem. 2003, 80, 17–21. [Google Scholar] [CrossRef]
- Xiros, C.; Christakopoulos, P. Enhanced ethanol production from brewer’s spent grain by a Fusarium oxysporum consolidated system. Biotechnol. Biofuels 2009, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Cooray, S.T.; Lee, J.J.L.; Chen, W.N. Evaluation of brewers’ spent grain as a novel media for yeast growth. AMB Express 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meneses, N.G.T.; Martins, S.; Teixeira, J.A.; Mussatto, S.I. Influence of extraction solvents on the recovery of antioxidant phenolic compounds from brewer’s spent grains. Sep. Purif. Technol. 2013, 108, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Celus, I.; Brijs, K.; Delcour, J.A. The effects of malting and mashing on barley protein extractability. J. Cereal Sci. 2006, 44, 203–211. [Google Scholar] [CrossRef]
- Robertson, J.A.; I’Anson, K.J.A.; Treimo, J.; Faulds, C.B.; Brocklehurst, T.F.; Eijsink, V.G.H.; Waldron, K.W. Profiling brewers’ spent grain for composition and microbial ecology at the site of production. LWT—Food Sci. Technol. 2010, 43, 890–896. [Google Scholar] [CrossRef]
- Gong, Z.; Wang, Q.; Shen, H.; Hu, C.; Jin, G.; Zhao, Z.K. Co-fermentation of cellobiose and xylose by Lipomyces starkeyi for lipid production. Bioresour. Technol. 2012, 117, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.; Pruthi, V.; Pruthi, P.A. Synchronized nutrient stress conditions trigger the diversion of CDP-DG pathway of phospholipids synthesis towards de novo TAG synthesis in oleaginous yeast escalating biodiesel production. Energy 2017, 139, 962–974. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Wu, S.; Zhao, X.; Shen, H.; Wang, Q.; Zhao, Z.K. Microbial lipid production by Rhodosporidium toruloides under sulfate-limited conditions. Bioresour. Technol. 2011, 102, 1803–1807. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hu, C.; Jin, G.; Zhao, X.; Zhao, Z.K. Phosphate-limitation mediated lipid production by Rhodosporidium toruloides. Bioresour. Technol. 2010, 101, 6124–6129. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Aggelis, G.; Marc, I. Kinetic profile of the cellular lipid composition in an oleaginous Yarrowia lipolytica capable of producing a cocoa-butter substitute from industrial fats. Antonie Van Leeuwenhoek 2001, 80, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C.; Wynn, J.P. The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv. Appl. Microbiol. 2002, 51, 1–51. [Google Scholar] [PubMed]
- Ratledge, C. Regulation of lipid accumulation in oleaginous micro-organisms. Biochem. Soc. Trans. 2002, 30, 1047–1050. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Seraphim, P.; Papanikolaou, S.; Seraphim, P.; Papanikolaou, S.; Seraphim, P.; Papanikolaou, S. Oleaginous Yeasts: Biochemical Events Related with Lipid Synthesis and Potential Biotechnological Applications. Ferment. Technol. 2012, 1, e103. [Google Scholar] [CrossRef]
- Matsakas, L.; Bonturi, N.; Miranda, E.A.; Rova, U.; Christakopoulos, P. High concentrations of dried sorghum stalks as a biomass feedstock for single cell oil production by Rhodosporidium toruloides. Biotechnol. Biofuels 2015, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, H.Y.; Chiang, L.C.; Ueng, P.P.; Tsao, G.T. Sequential utilization of mixed monosaccharides by yeasts. Appl. Environ. Microbiol. 1982, 43, 840–845. [Google Scholar] [PubMed]
- Sànchez i Nogué, V.; Black, B.A.; Kruger, J.S.; Singer, C.A.; Ramirez, K.J.; Reed, M.; Cleveland, N.S.; Singer, E.R.; Yi, X.; Yeap, R.Y.; et al. Integrated diesel production from lignocellulosic sugars via oleaginous yeast. Green Chem. 2018, 20, 4349–4365. [Google Scholar] [CrossRef]
- Zhao, X.; Kong, X.; Hua, Y.; Feng, B.; Zhao, Z.K. Medium optimization for lipid production through co-fermentation of glucose and xylose by the oleaginous yeastLipomyces starkeyi. Eur. J. Lipid Sci. Technol. 2008, 110, 405–412. [Google Scholar] [CrossRef]
- Huang, C.; Zong, M.H.; Wu, H.; Liu, Q.P. Microbial oil production from rice straw hydrolysate by Trichosporon fermentans. Bioresour. Technol. 2009, 100, 4535–4538. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, W.; Li, Y.; Zhang, Y. Lipid production by Rhodosporidium toruloides Y2 in bioethanol wastewater and evaluation of biomass energetic yield. Bioresour. Technol. 2013, 127, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Canakci, M.; Van Gerpen, J. Biodiesel production from oils and fats with high free fatty acids. Trans. ASAE 2001, 44, 1429–1436. [Google Scholar] [CrossRef]
- Moser, B.R. Biodiesel production, properties, and feedstocks. Vitr. Cell. Dev. Biol.-Plant 2009, 45, 229–266. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Sartaj, K.; Pruthi, V.; Pruthi, P.A. Sustainable biodiesel production from oleaginous yeasts utilizing hydrolysates of various non-edible lignocellulosic biomasses. Renew. Sustain. Energy Rev. 2016, 62, 836–855. [Google Scholar] [CrossRef]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process. Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Knothe, G. Analyzing biodiesel: Standards and other methods. J. Am. Oil Chem. Soc. 2006, 83, 823–833. [Google Scholar] [CrossRef]
- Verma, P.; Sharma, M.P.; Dwivedi, G. Evaluation and enhancement of cold flow properties of palm oil and its biodiesel. Energy Rep. 2016, 2, 8–13. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernández, C.M.; Casas, A.; Rodríguez, L.; Pérez, Á. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Knothe, G.; Razon, L.F. Biodiesel fuels. Prog. Energy Combust. Sci. 2017, 58, 36–59. [Google Scholar] [CrossRef]
- Kumar, N. Oxidative stability of biodiesel: Causes, effects and prevention. Fuel 2017, 190, 328–350. [Google Scholar] [CrossRef]
- Hoekman, S.K.; Broch, A.; Robbins, C.; Ceniceros, E.; Natarajan, M. Review of biodiesel composition, properties, and specifications. Renew. Sustain. Energy Rev. 2012, 16, 143–169. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Mehtani, J.; Pruthi, V.; Pruthi, P.A. Assessment of fuel properties on the basis of fatty acid profiles of oleaginous yeast for potential biodiesel production. Renew. Sustain. Energy Rev. 2017, 77, 604–616. [Google Scholar] [CrossRef]
- Fassinou, W.F.; Sako, A.; Fofana, A.; Koua, K.B.; Toure, S. Fatty acids composition as a means to estimate the high heating value (HHV) of vegetable oils and biodiesel fuels. Energy 2010, 35, 4949–4954. [Google Scholar] [CrossRef]
- Sluiter, A.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Extractives in Biomass: Laboratory Analytical Procedure (LAP); NREL/TP-510-42619; National Renewable Energy Laboratory: Golden, CO, USA, 2005; pp. 1–9. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Ash in Biomass Laboratory Analytical Procedure (LAP); NREL/TP-510-42622; National Renewable Energy Laboratory: Golden, CO, USA, 2005; pp. 1–5. [Google Scholar]
- Ravindran, R.; Jaiswal, S.; Abu-Ghannam, N.; Jaiswal, A.K. A comparative analysis of pretreatment strategies on the properties and hydrolysis of brewers’ spent grain. Bioresour. Technol. 2018, 248, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Kalogiannis, K.G.; Matsakas, L.; Aspden, J.; Lappas, A.A.; Rova, U.; Christakopoulos, P. Acid assisted organosolv delignification of beechwood and pulp conversion towards high concentrated cellulosic ethanol via high gravity enzymatic hydrolysis and fermentation. Molecules 2018, 23, 1647. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. Heterotrophic cultivation of Auxenochlorella protothecoides using forest biomass as a feedstock for sustainable biodiesel production. Biotechnol. Biofuels 2018, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Laurens, L.M.L.; Quinn, M.; Van Wychen, S.; Templeton, D.W.; Wolfrum, E.J. Accurate and reliable quantification of total microalgal fuel potential as fatty acid methyl esters by in situ transesterification. Anal. Bioanal. Chem. 2012, 403, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Talebi, A.F.; Tabatabaei, M.; Chisti, Y. BiodieselAnalyzer: A User-Friendly Software for Predicting the Properties of Prospective Biodiesel. Biofuel Res. J. 2014, 2, 55–57. [Google Scholar] [CrossRef]
Sample Availability: Samples of the lipids produced during this work are available from the authors at a reasonable request. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellulose (% w/w) | Xylan (% w/w) | Protein (% w/w) | Lignin (% w/w) | References |
|---|---|---|---|---|
| 21.73 ± 1.36 | 13.63 ± 0.82 | 24.69 ± 1.04 | 19.40 ± 0.34 | [23] |
| 0.3 (1→3; 1→4)-β-Glucan | 22.5 (Arabinoxylan) | 26.7 | n.d. | [24] |
| 13–21 | 21–27 | 10–18 | 12–16 | [25] |
| 38.88 ± 0.87 | 18.23 ± 1.17 | 12.54 ± 0.65 | 13.7 ± 0.98 | This study |
| Fatty Acids (%) | Glucose C/N 20 | Glucose C/N 500 | Xylose C/N 20 | Xylose C/N 500 | Glucose + Xylose C/N 20 | Glucose + Xylose C/N 500 | BSG C/N 20 | BSG C/N 500 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Saturated Fatty Acid (SFA) | (C14:0) | ND | 21.67 | 0.98 | 30.60 | ND | 24.49 | 0.89 | 30.46 | 0.91 | 26.87 | 0.86 | 31.4 | ND | 20.07 | 1.00 | 33.10 |
| (C16:0) | 13.33 | 21.52 | 15.36 | 21.52 | 16.37 | 20.96 | 14.25 | 21.50 | |||||||||
| (C18:0) | 6.69 | 8.10 | 7.56 | 6.32 | 7.76 | 8.34 | 3.96 | 7.04 | |||||||||
| (C20:0) | ND | ND | ND | ND | ND | 0.47 | ND | 2.39 | |||||||||
| (C24:0) | 1.65 | 1.18 | 1.57 | 1.73 | 1.83 | 0.77 | 1.86 | 1.17 | |||||||||
| Mono Unsaturated Fatty Acid (MUFA) | (C18:1n9t) | 46.25 | 46.25 | 51.14 | 51.14 | 47.91 | 47.91 | 42.37 | 42.37 | 43.45 | 43.45 | 52.37 | 52.37 | 54.84 | 54.84 | 50.85 | 50.85 |
| Poly Unsaturated Fatty Acid (PUFA) | (C18:2n6c) | 17.97 | 20.52 | 11.60 | 14.44 | 15.80 | 18.48 | 12.30 | 15.3 | 23.95 | 7.19 | 10.24 | 14.31 | 20.63 | 24.84 | 12.14 | 13.97 |
| (C18:3n3) | 1.43 | 1.60 | 1.70 | 1.77 | 1.96 | 2.31 | 3.45 | 0.29 | |||||||||
| (C22:2) | 1.12 | 1.24 | 0.98 | 1.23 | 1.28 | 1.76 | 0.76 | 1.54 | |||||||||
| Total Fatty Acids (%) | 88.44 | 96.18 | 90.88 | 88.13 | 97.51 | 98.08 | 99.75 | 97.92 | |||||||||
| Biodiesel Properties | Units | Glucose C/N 20 | Glucose C/N 500 | Xylose C/N 20 | Xylose C/N 500 | Glucose + Xylose C/N 20 | Glucose + Xylose C/N 500 | BSG C/N 20 | BSG C/N 500 | Biodiesel Standards | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ASTM D6751 Limits | EN 14214 Limits | ||||||||||
| Long chain saturation factor | - | 7.978 | 8.562 | 8.456 | 8.772 | 5.517 | 6.736 | 3.405 | 8.060 | - | - |
| Oxidative stability, 110 °C | h | 8.7 | 11.5 | 9.3 | 11.0 | 7.1 | 12.0 | 7.5 | 12.1 | 3 min | 6 min |
| Density | g/cm3 | 0.77 | 0.85 | 0.80 | 0.77 | 0.84 | 0.85 | 0.86 | 0.85 | - | 0.86–0.90 |
| Cold filter plugging point | °C | 8.6 | 10.4 | 10.1 | 11.1 | 0.9 | 4.7 | 0.6 | 8.8 | - | - |
| Cloud Point | °C | 2.0 | 6.3 | 3.1 | 6.3 | 3.6 | 6.0 | 2.5 | 6.3 | ||
| Pour Point | °C | −4.6 | 0.0 | −3.5 | 0.0 | −2.9 | −0.3 | −4.1 | 0.0 | ||
| Cetane number | - | 59.2 | 57.5 | 58.7 | 61.8 | 54.3 | 57.2 | 52.1 | 58.2 | 47 min | 51 min |
| Kinematic Viscosity | mm2/s | 3.4 | 3.9 | 3.5 | 3.4 | 3.7 | 3.8 | 3.8 | 3.8 | 1.9–6.0 | 3.5–5 |
| Saponification value | mg KOH/g-oil | 177.1 | 197.0 | 182.4 | 178.4 | 193.3 | 197.0 | 197.0 | 195.8 | 0.50 min | 0.50 min |
| Iodine value | mgI2/100g | 79.8 | 73.3 | 77.8 | 67.1 | 89.8 | 74.6 | 97.3 | 70.8 | - | 120 max |
| High heating value | MJ/kg | 35.0 | 38.5 | 36.0 | 34.8 | 37.8 | 38.5 | 38.7 | 38.2 | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, A.; Mikes, F.; Bühler, S.; Matsakas, L. Valorization of Brewers’ Spent Grain for the Production of Lipids by Oleaginous Yeast. Molecules 2018, 23, 3052. https://doi.org/10.3390/molecules23123052
Patel A, Mikes F, Bühler S, Matsakas L. Valorization of Brewers’ Spent Grain for the Production of Lipids by Oleaginous Yeast. Molecules. 2018; 23(12):3052. https://doi.org/10.3390/molecules23123052
Chicago/Turabian StylePatel, Alok, Fabio Mikes, Saskja Bühler, and Leonidas Matsakas. 2018. "Valorization of Brewers’ Spent Grain for the Production of Lipids by Oleaginous Yeast" Molecules 23, no. 12: 3052. https://doi.org/10.3390/molecules23123052
APA StylePatel, A., Mikes, F., Bühler, S., & Matsakas, L. (2018). Valorization of Brewers’ Spent Grain for the Production of Lipids by Oleaginous Yeast. Molecules, 23(12), 3052. https://doi.org/10.3390/molecules23123052