Estrogenic Effects of the Extracts from the Chinese Yam (Dioscorea opposite Thunb.) and Its Effective Compounds in Vitro and in Vivo

Abstract
:
1. Introduction
2. Results
2.1. The Influences of Chinese Yam on MCF-7 Cells
2.2. The Influences of Chinese Yam on Immature Female Swiss Mice
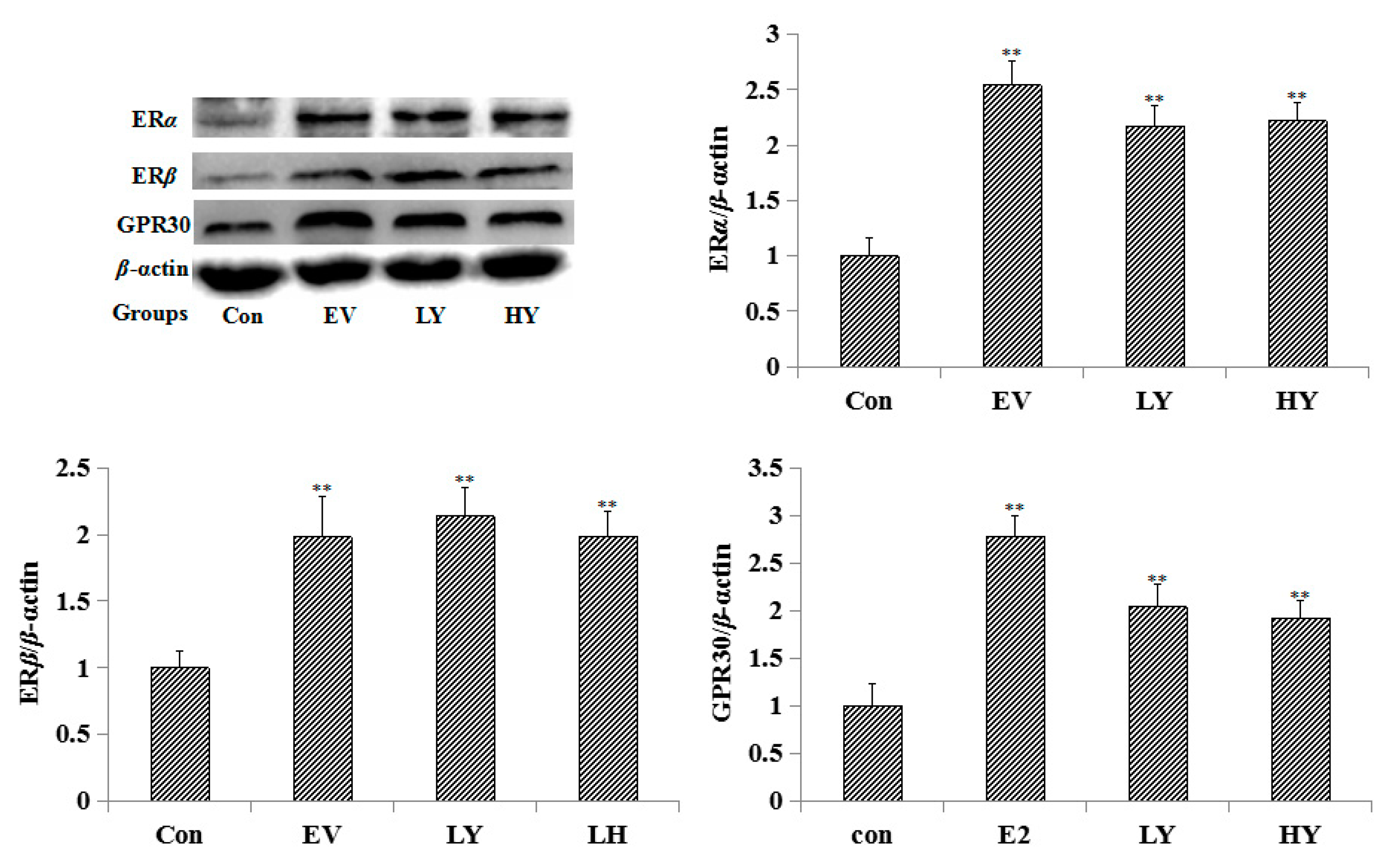
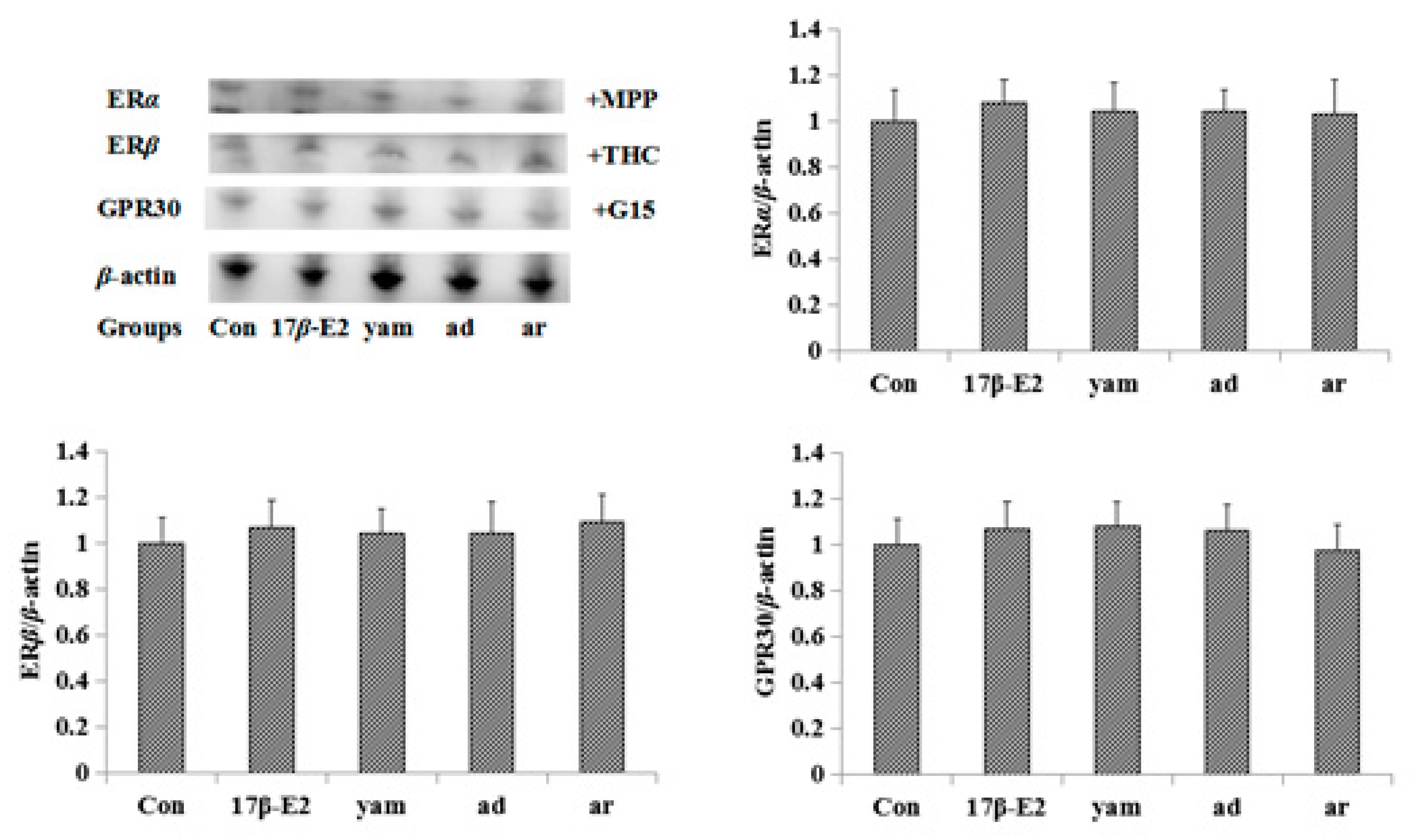
2.3. Effect of Chinese Yam on the Expression of ERα, ERβ and GPR30 in the Uterus
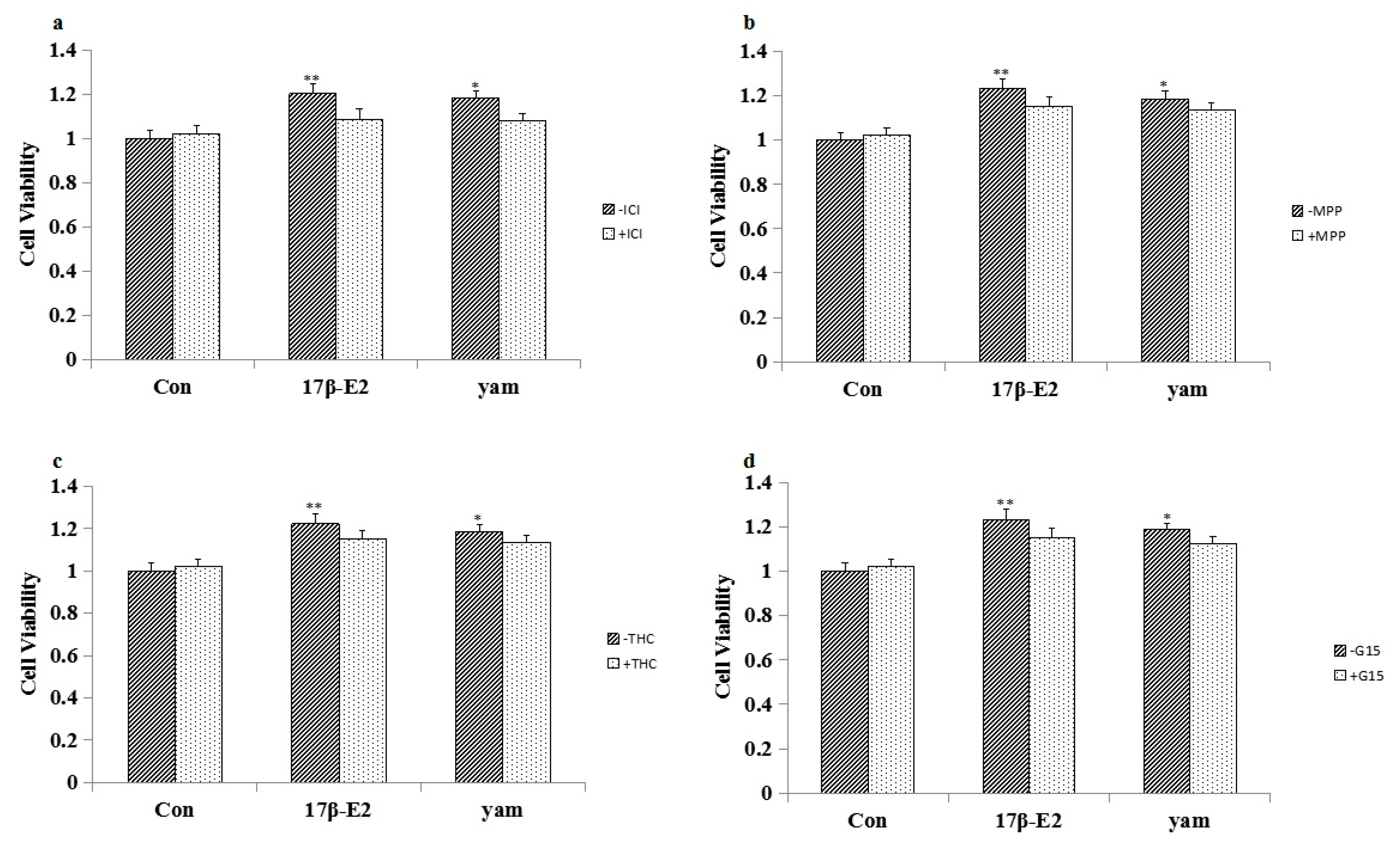
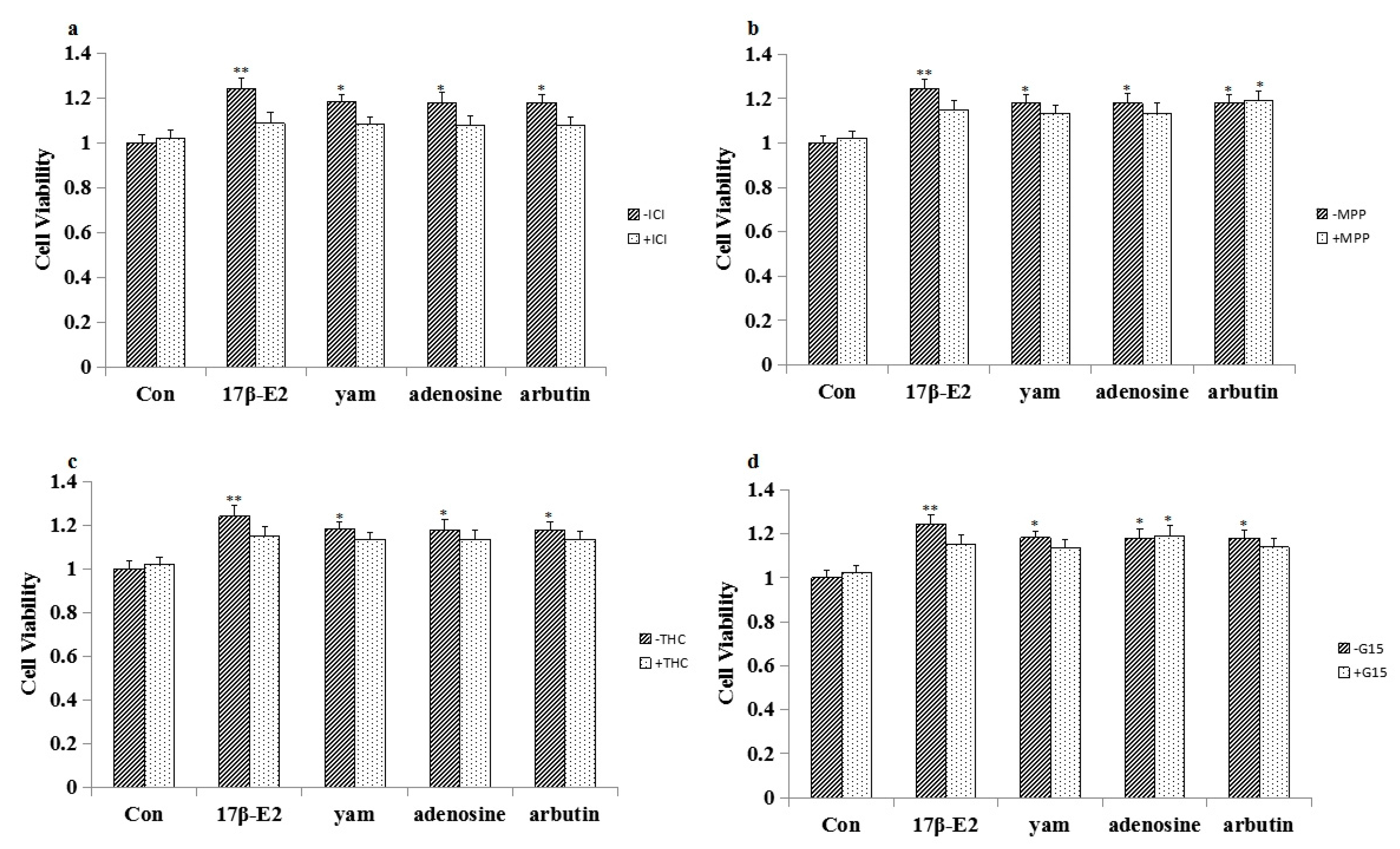
2.4. Effect of ICI182,780, MPP, THC and G15 on the Proliferation of Yam-Stimulated MCF-7 Cells
2.5. The Influences of Adenosine and Arbutin on MCF-7 Cells
2.6. The Influences of Adenosine and Arbutin on Immature Female Swiss Mice
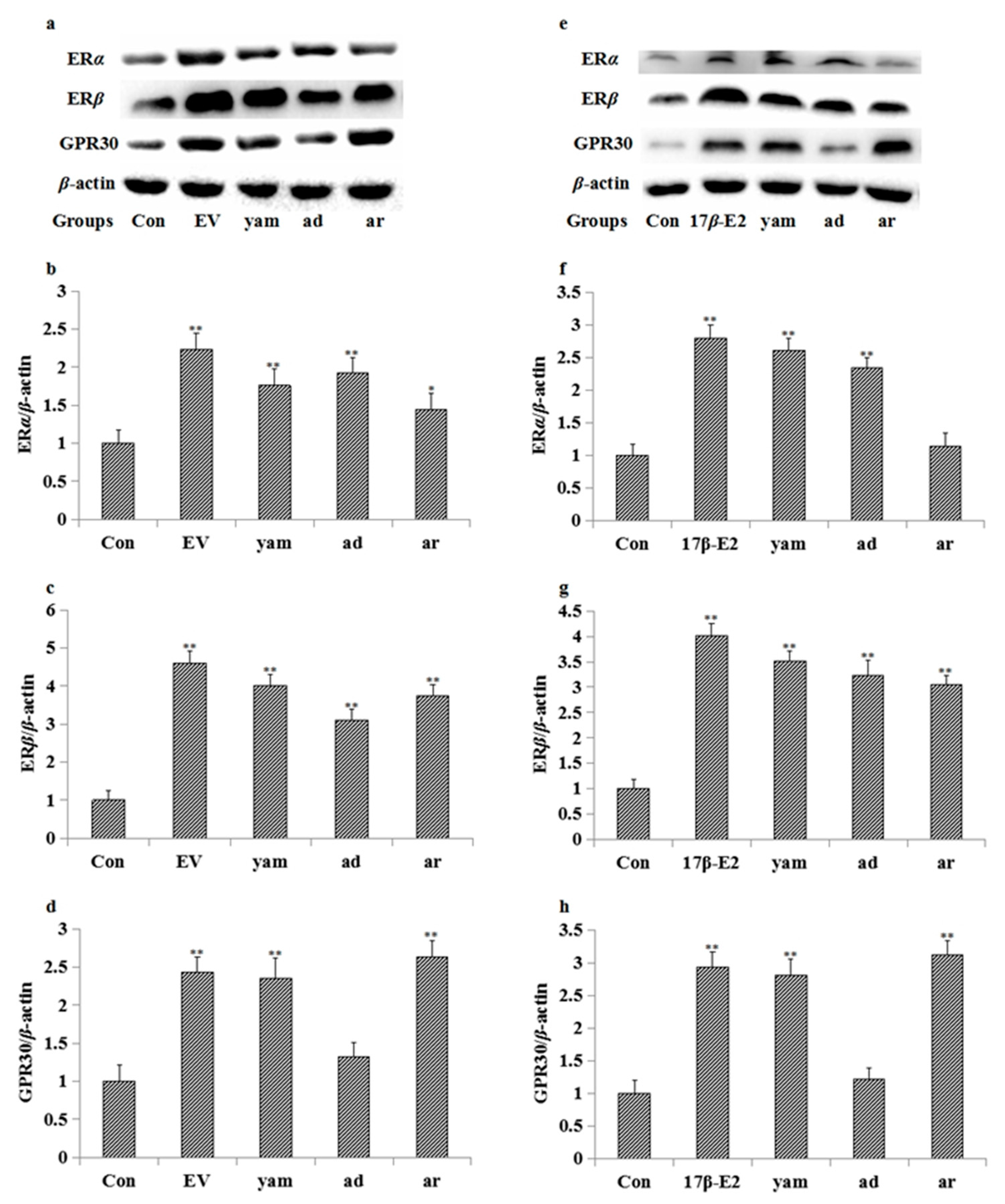
2.7. Effect on the Expression of ERα, ERβ and GPR30 in MCF-7 Cells and the Uterus
2.8. Effect of ICI182,780, MPP, THC and G-15 on Adenosine- and Arbutin-Stimulated MCF-7 Cell Proliferation and the Expression of ERα, ERβ and GPR30 in MCF-7 Cells
2.9. Pharmacological Experiment with Serum
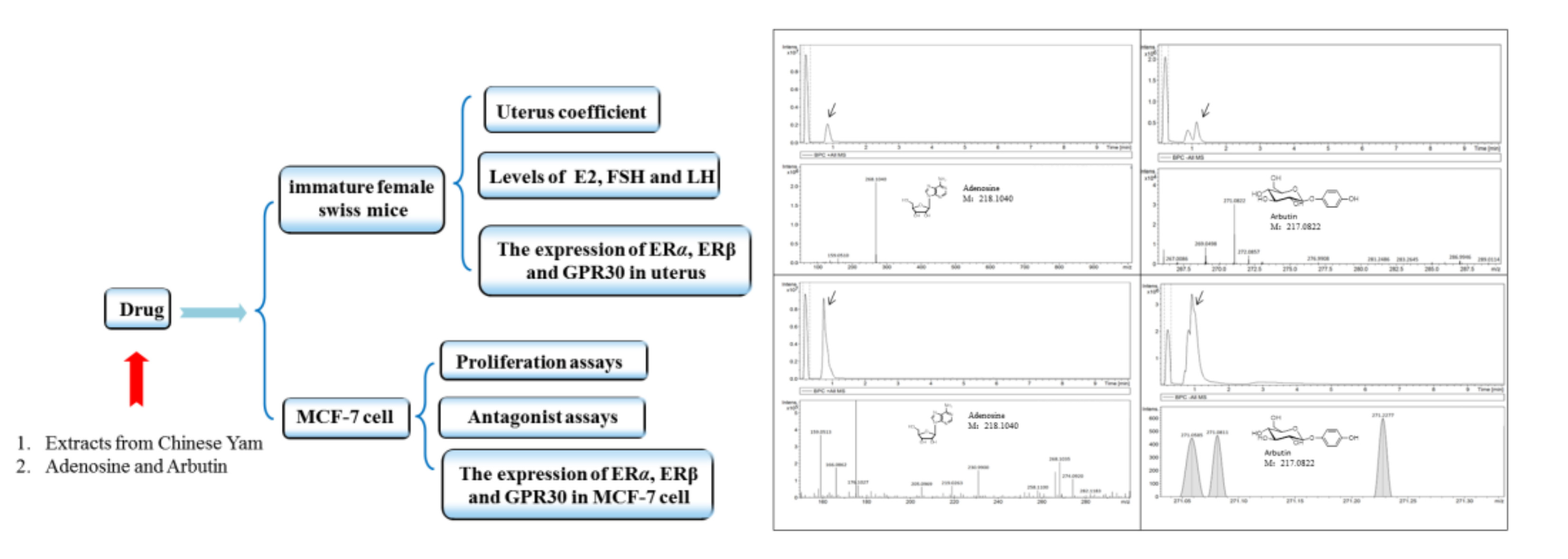
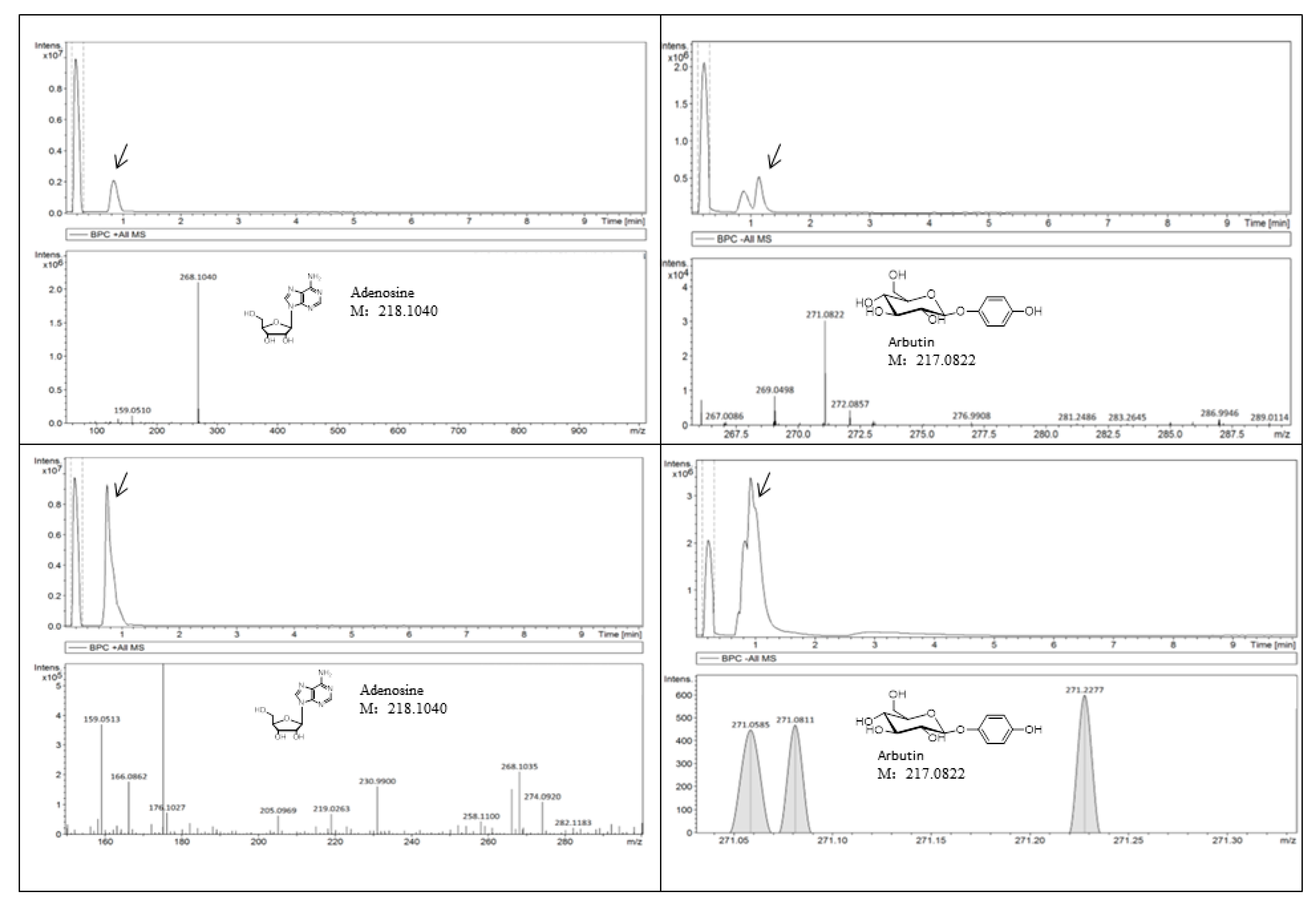
2.10. Quantitative Analysis of Adenosine and Arbutin
3. Discussion
4. Materials and Methods
4.1. Plant Material and Reagents
4.2. Cell Culture and Treatment
4.3. MTT Assay
4.4. Animals
4.5. Western Blot
4.6. ELISA
4.7. Serum Pharmacology Experiment
4.8. Quantitative Analysis of Adenosine and Arbutin
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MCF-7 cell | Breast adenocarcinoma cell line |
| LH | Luteinizing hormone |
| FSH | Follicle stimulating hormone |
| ICI182,780 | Faslodex |
| MPP | Specific ERα antagonist methylpiperidino-pyrazole |
| THC | Specific ERβ antagonist Delta (9) –tetrahydrocannabinol |
| G-15 | Specific GPR30 antagonist |
| TBST | Tris Buffered Saline with Tween-20 |
| MTT | Methyl thiazolyl tetrazolium |
| PBS | Phosphate Buffer Saline |
| DMSO | Dimethyl sulfoxide |
| LC-MS | Liquid chromatography-mass spectrometry |
| ERT | Estrogen replacement therapy |
| TCM | Traditional Chinese Medicine |
| 17β-E2 | 17-beta-estradiol |
| PVDF | Poiy vinylidene fluoride |
| AD | Alzheimer’s disease |
| VEGF | vascular endothelial growth factor |
| SERMs | Selective estrogen receptor modulators |
References
- Webster, R.W. Aboriginal Women and Menopause. J. Obstet. Gynaecol. Can. 2002, 24, 591–596. [Google Scholar] [CrossRef]
- Anna, M.; Sawka, A.; Huh, L.; Dolovich, A.; Papaioannou, K.E.; Lehana, T.; Amiram, G.; Amie, C.; Mary, M.; Meir, S.; et al. Attitudes of Women Who are Currently Using or Recently Stopped Estrogen Replacement Therapy with or without Progestins: Results of the Aware Survey. J. Obstet. Gynaecol. Can. 2004, 26, 237–242. [Google Scholar]
- Seki, K.; Minami, Y.; Nishikawa, M.; Kawata, S.; Miyoshi, S.; Imai, Y.; Tarui, S. “Nonalcoholic steatohepatitis” induced by massive doses of synthetic estrogen. Gastroenterol. Jpn. 1983, 18, 197–203. [Google Scholar]
- Marks, K.J.; Hartman, T.J.; Taylor, E.V.; Rybak, M.E.; Northstone, K. Exposure to phytoestrogens in utero and age at menarche in a contemporary British cohort. Environ. Res. 2017, 19, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wang, Y.; Li, X. Purification and structural characterization of Chinese yam polysaccharide and its activities. Carbohydr. Polym. 2015, 117, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Lin, K.W. Effects of heating temperature on the total phenolic compound, antioxidative ability and the stability of dioscorin of various yam cultivars. Food Chem. 2007, 101, 955–963. [Google Scholar] [CrossRef]
- Choi, E.M.; Koo, S.J.; Hwang, J.K. Immune cell stimulating activity of mucopolysaccharide isolated from yam (Dioscorea batatas). Ethnopharmacology 2004, 91, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; Mazur, W. Phyto-oestrogens and Western diseases. Ann. Med. 1997, 29, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Glazier, M.G.; Bowman, M.A. A review of the evidence for the use of phytoestrogens as a replacement for traditional estrogen replacement therapy. Arch. Intern. Med. 2001, 161, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Tham, D.M.; Gardner, C.D.; Haskell, W.L. Clinical review 97: Potential health benefits of dietary phytoestrogens: A review of the clinical, epidemiological, and mechanistic evidence. J. Clin. Endocrinol. Metab. 1998, 83, 2223–2235. [Google Scholar] [PubMed]
- Montilla, P.; Túnez, I.; Muñoz, M.C.; Delgado, M.J.; Salcedo, M. Hyperlipidemic nephropathy induced by adriamycin in ovariectomized rats: role of free radicals and effect of 17-beta-estradiol administration. Nephron 2000, 85, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Gouva, L.; Tsatsoulis, A. The role of estrogens in cardiovascular disease in the aftermath of clinical trials. Hormones 2006, 3, 237–246. [Google Scholar] [CrossRef]
- Henderson, V.W. Estrogens, episodic memory, and Alzheimer's disease: a critical update. Seminars in Reproductive Medicine 2009, 27, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.T.; Lefkowitz, R. J. Regulation of rabbit myometrial alpha adrenergic receptors by estrogen and progesterone. J. Clin. Investig. 1977, 60, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H. Coexistence of cytoplasmic and nuclear estrogen receptors. A histochemical study on human mammary cancer and rabbit uterus. Cancer 1989, 64, 1461–1466. [Google Scholar] [CrossRef]
- Carlson, R. A.; Gorski, J. Characterization of a unique population of unfilled estrogen-binding sites associated with the nuclear fraction of immature rat uteri. Endocrinology 1980, 106, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Degani, H.; Victor, T.A.; Kaye, A.M. Effects of 17 beta-estradiol on high energy phosphate concentrations and the flux catalyzed by creatine kinase in immature rat uteri: 31P nuclear magnetic resonance studies. Endocrinology 1988, 122, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Wang, D.; He, R.; Zhu, H.; Wang, Y.; He, S. Medicinal herb research: serum pharmacological method and plasma pharmacological method. Biol. Pharm. Bull. 2010, 33, 217–224. [Google Scholar]
- Kewen, Z.; Peng, S.; Yaxing, Z.; Xinchao, Y.; Ping, L.; Tinghuai, W. Estrogen stimulated migration and invasion of estrogen receptor-negative breast cancer cells involves an ezrin-dependent crosstalk between G protein-coupled receptor 30 and estrogen receptor beta signaling. Steroids 2016, 14, 147–153. [Google Scholar]
- Dowsett, M.; Johnston, S.R.; Iveson, T.; Smith, I.E. Response to specific anti-oestrogen (ICI182780) in tamoxifen-resistant breast cancer. Lancet 1998, 345, 525–527. [Google Scholar] [CrossRef]
- David, S.; Jean, T. Estrogen regulation and ion dependence of taurine uptake by MCF-7 human breast cancer cells. Cell Mol. Boil. Lett. 2007, 12, 2177–2183. [Google Scholar]
- Watanabe, K.; Motoya, E.; Matsuzawa, N.; Funahashi, T.; Kimura, T.; Matsunaga, T.; Arizono, K.; Yamamoto, I. Marijuana extracts possess the effects like the endocrine disrupting chemicals. Toxicology 2004, 206, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Duan, Y.H.; Jing, W. Baicalein, unlike 4-hydroxytamoxifen but similar to G15, suppresses17β-estradiol-induced cell invasion, and matrix metalloproteinase-9 expressionand activation in MCF-7 human breast cancer cells. Oncol. Lett. 2017, 14, 1823–1830. [Google Scholar]
- Hastings, J.M.; Licence, D.R.; Burton, G.J. Soluble vascular endothelial growth factor receptor 1 inhibits edema and epithelial proliferation induced by 17beta-estradiol in the mouse uterus. Endocrinology 2002, 144, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; Ijzerman, A.P.; Jacobson, K.A.; lotz, K.N.; Linden, J. Nomenclature and classification of adenosine receptors. Pharmacol. Rev. 2001, 52, 527–552. [Google Scholar]
- Fredholm, B.B. Adensine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ. 2007, 14, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Mishima, Y.; Hatta, Y. Induction of melanogenesis suppression: Cellular pharmacology and mode of differential action. Pigment Melanoma Cell Res. 1988, 367–374. [Google Scholar] [CrossRef]
- Migas, P.; Krauze-Baranowska, M. The significance of arbutin and its derivatives in therapy and cosmetics. Phytochem. Lett. 2015, 13, 35–40. [Google Scholar] [CrossRef]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Dose | Cell Viability |
|---|---|---|
| Con | — | 1.00 ± 0.021 |
| 17β-E2(μM) | 1 | 1.25 ± 0.015 ** |
| Yam (mg/mL) | 1 | 1.31 ± 0.023 ** |
| 10−1 | 1.26 ± 0.028 ** | |
| 10−2 | 1.19 ± 0.022 * | |
| 10−3 | 1.17 ± 0.030 * | |
| 10−4 | 1.09 ± 0.027 |
| Groups | Dose (mg/kg) | Uterus Coefficient (%) | E2 (pmol/L) | FSH (mIU/mL) | LH (mIU/mL) |
|---|---|---|---|---|---|
| Con | — | 0.1002 ± 0.009 | 31.12 ± 3.01 | 45.32 ± 5.90 | 3.69 ± 0.69 |
| EV | 0.33 | 0.2515 ± 0.021 ** | 38.94 ± 2.98 ** | 53.17 ± 4.97 ** | 4.77 ± 0.74 ** |
| LY | 1630 | 0.1227 ± 0.027 * | 39.21 ± 3.97 ** | 55.13 ± 4.26 ** | 3.54 ± 0.29 |
| HY | 3260 | 0.1269 ± 0.023 * | 42.09 ± 5.04 ** | 56.13 ± 3.26 ** | 3.61 ± 0.48 |
| Groups | Dose (μM) | Cell Viability |
|---|---|---|
| Con | — | 1.00 ± 0.021 |
| 17β-E2 | 1 | 1.27 ± 0.016 ** |
| Yam (mg/ml) | 10−2 | 1.21 ± 0.024 * |
| adenosine | 5 | 1.20 ± 0.028 * |
| 10 | 1.25 ± 0.022 ** | |
| arbutin | 5 | 1.19 ± 0.030 * |
| 10 | 1.25 ± 0.027 ** |
| Groups | Dose (mg/kg) | Uterus Coefficient (%) | E2 (p mol/L) | FSH (mIU/mL) | LH (mIU/mL) |
|---|---|---|---|---|---|
| Con | — | 0.0994 ± 0.013 | 29.05 ± 5.11 | 45.32 ± 5.90 | 4.55 ± 0.54 |
| EV | 0.33 | 0.2254 ± 0.015 ** | 40.16 ± 8.09 ** | 56.30 ± 5.12 ** | 5.14 ± 0.58 ** |
| yam | 1630 | 0.1323 ± 0.036 * | 38.23 ± 1.67 ** | 62.43 ± 7.72 ** | 4.16 ± 0.42 |
| adenosine | 50 | 0.1354 ± 0.023 * | 42.96 ± 5.36 ** | 60.14 ± 6.17 ** | 4.07 ± 0.41 |
| arbutin | 50 | 0.1271 ± 0.018 * | 38.39 ± 3.81 ** | 58.26 ± 6.32 ** | 4.19 ± 0.39 |
| Groups | Cell Viability |
|---|---|
| Con | 1.00 ± 0.021 |
| E2 | 1.23 ± 0.022 ** |
| Yam | 1.25 ± 0.019 ** |
| Adenosine | 1.24 ± 0.027 ** |
| Arbutin | 1.21 ± 0.030 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, M.; Zhang, L.; Li, M.; Zhang, B.; Zhou, N.; Ke, Y.; Feng, W.; Zheng, X. Estrogenic Effects of the Extracts from the Chinese Yam (Dioscorea opposite Thunb.) and Its Effective Compounds in Vitro and in Vivo. Molecules 2018, 23, 11. https://doi.org/10.3390/molecules23020011
Zeng M, Zhang L, Li M, Zhang B, Zhou N, Ke Y, Feng W, Zheng X. Estrogenic Effects of the Extracts from the Chinese Yam (Dioscorea opposite Thunb.) and Its Effective Compounds in Vitro and in Vivo. Molecules. 2018; 23(2):11. https://doi.org/10.3390/molecules23020011
Chicago/Turabian StyleZeng, Mengnan, Li Zhang, Miao Li, Beibei Zhang, Ning Zhou, Yingying Ke, Weisheng Feng, and Xiaoke Zheng. 2018. "Estrogenic Effects of the Extracts from the Chinese Yam (Dioscorea opposite Thunb.) and Its Effective Compounds in Vitro and in Vivo" Molecules 23, no. 2: 11. https://doi.org/10.3390/molecules23020011
APA StyleZeng, M., Zhang, L., Li, M., Zhang, B., Zhou, N., Ke, Y., Feng, W., & Zheng, X. (2018). Estrogenic Effects of the Extracts from the Chinese Yam (Dioscorea opposite Thunb.) and Its Effective Compounds in Vitro and in Vivo. Molecules, 23(2), 11. https://doi.org/10.3390/molecules23020011