Organ-Specific Metabolic Shifts of Flavonoids in Scutellaria baicalensis at Different Growth and Development Stages


Abstract
:1. Introduction
2. Results and Discussion
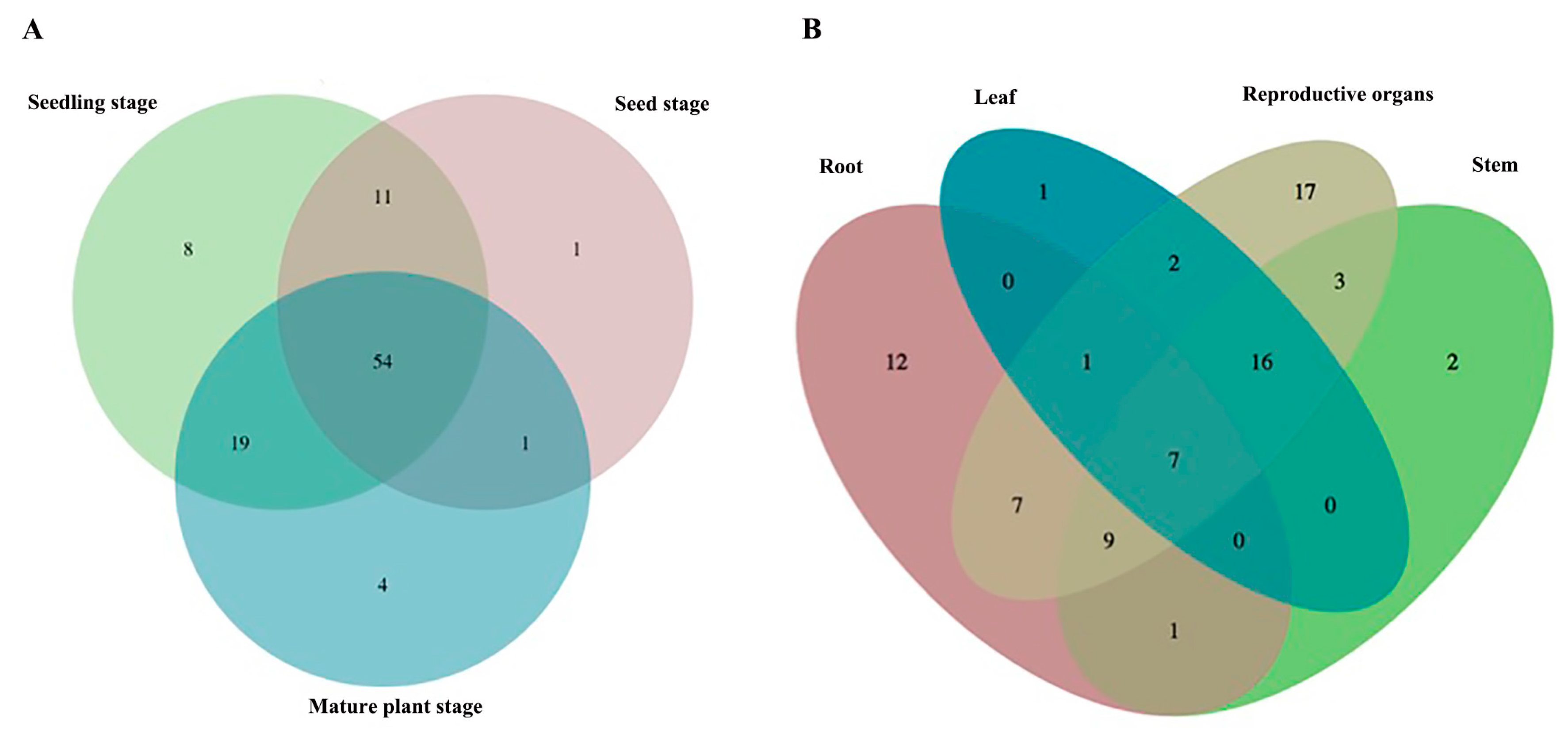
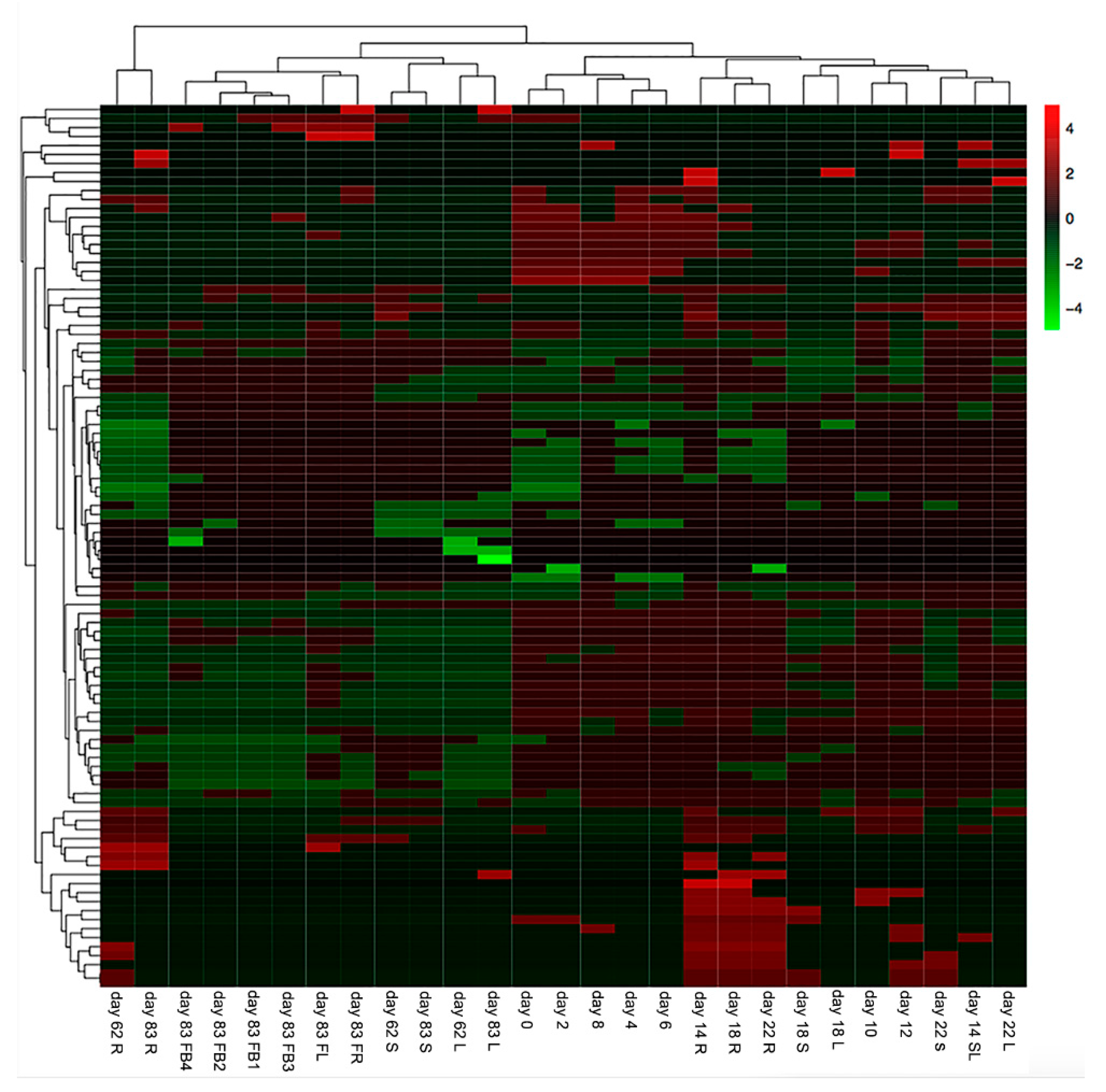
2.1. The Metabolic Profiles of S. baicalensis Were Organ-Specific and Changed at Different Growth and Development Stages
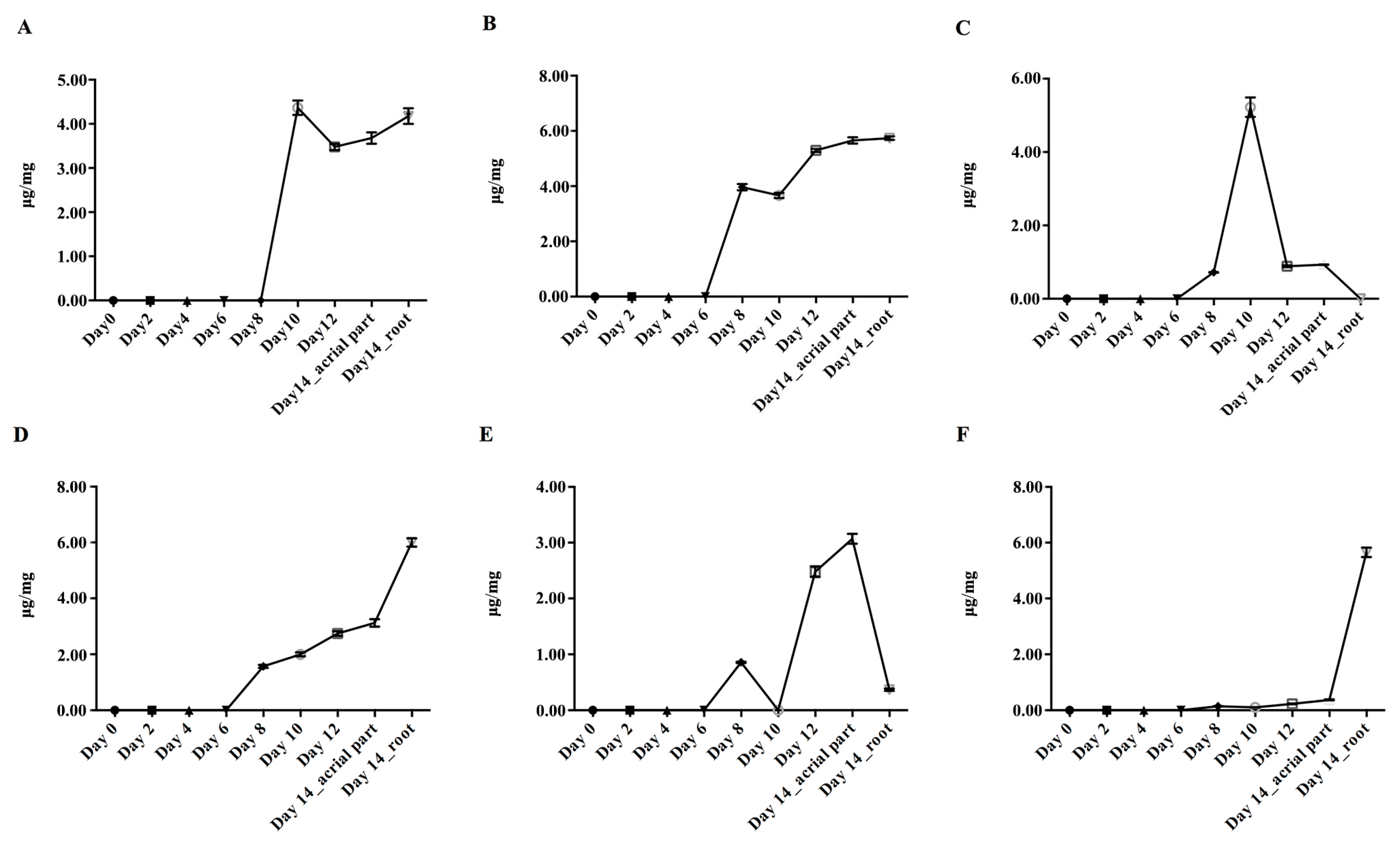
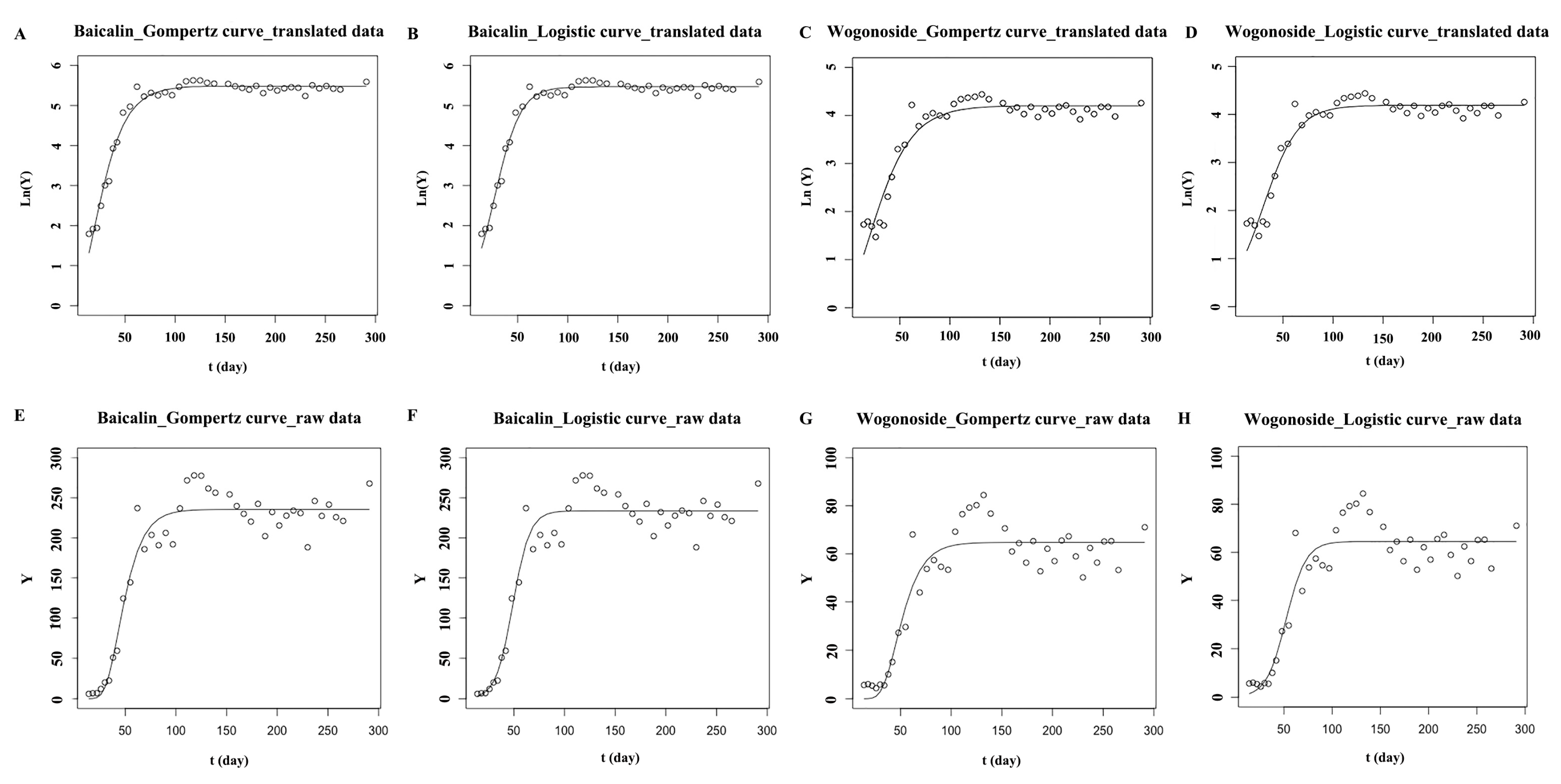
2.2. Dynamic Accumulation of Main Flavonoids in S. baicalensis from Seed Sowing to the True Leaf Coming
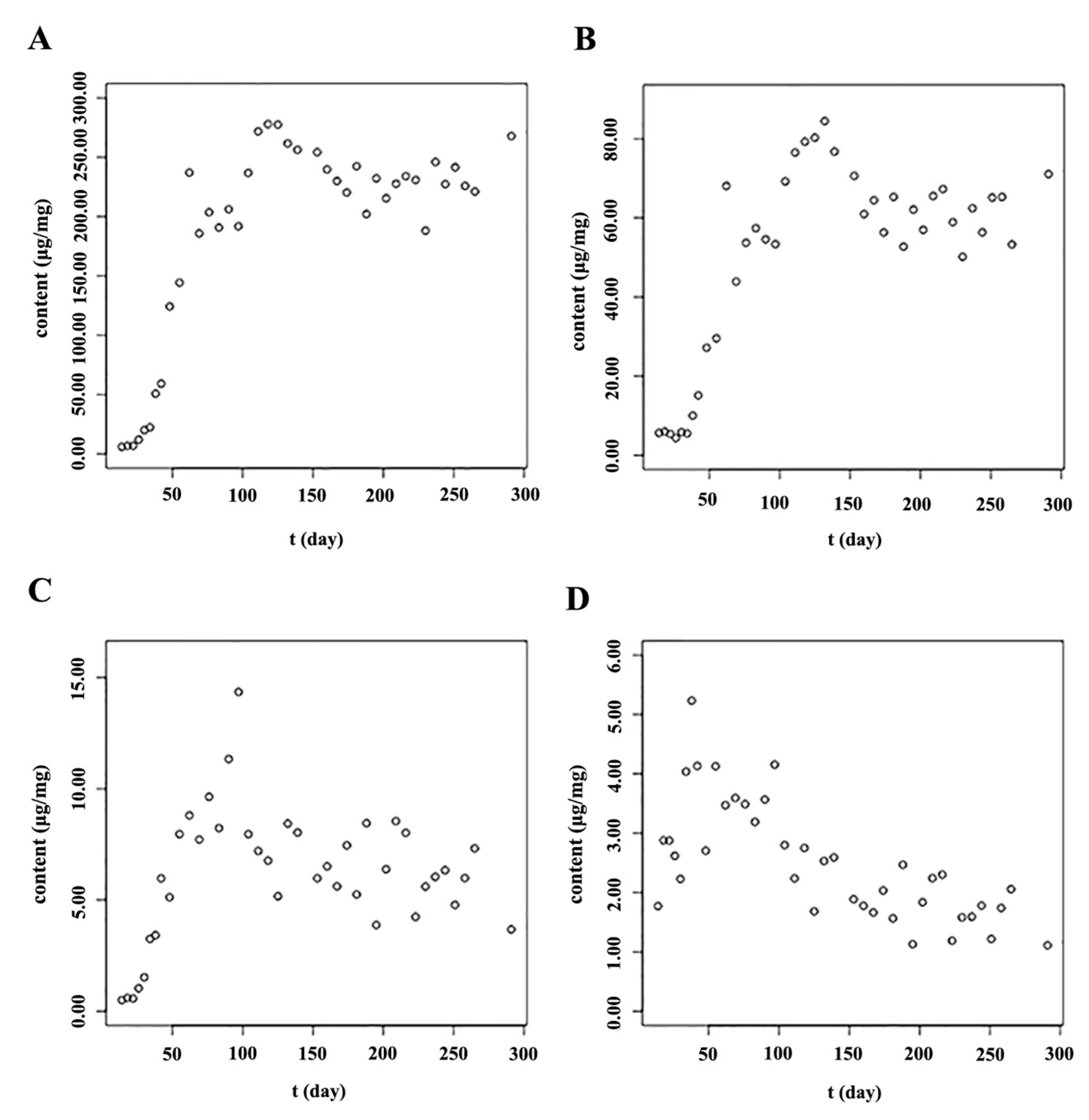
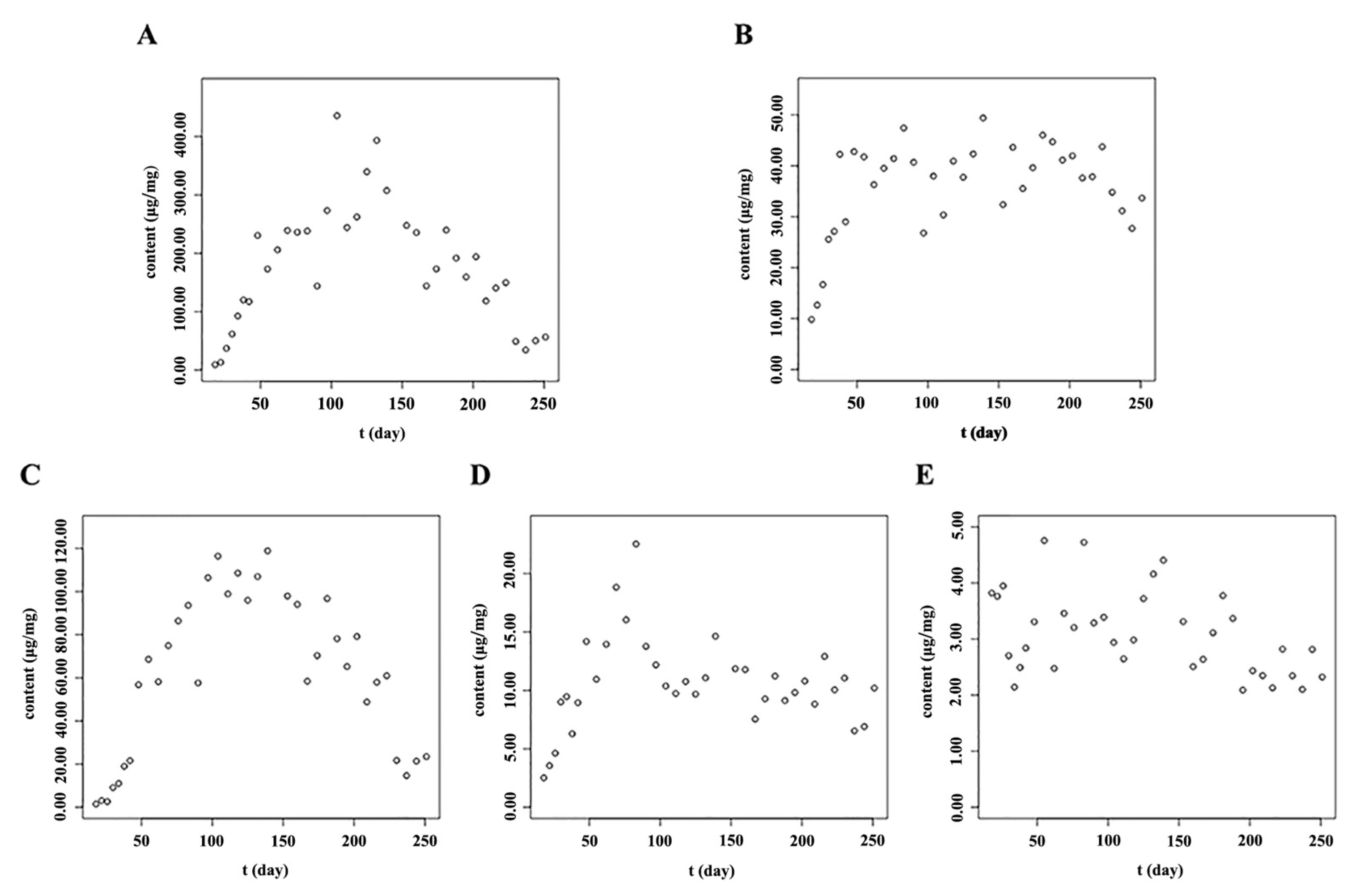
2.3. The Accumulation Patterns of Main Flavonoids in Roots of S. baicalensis
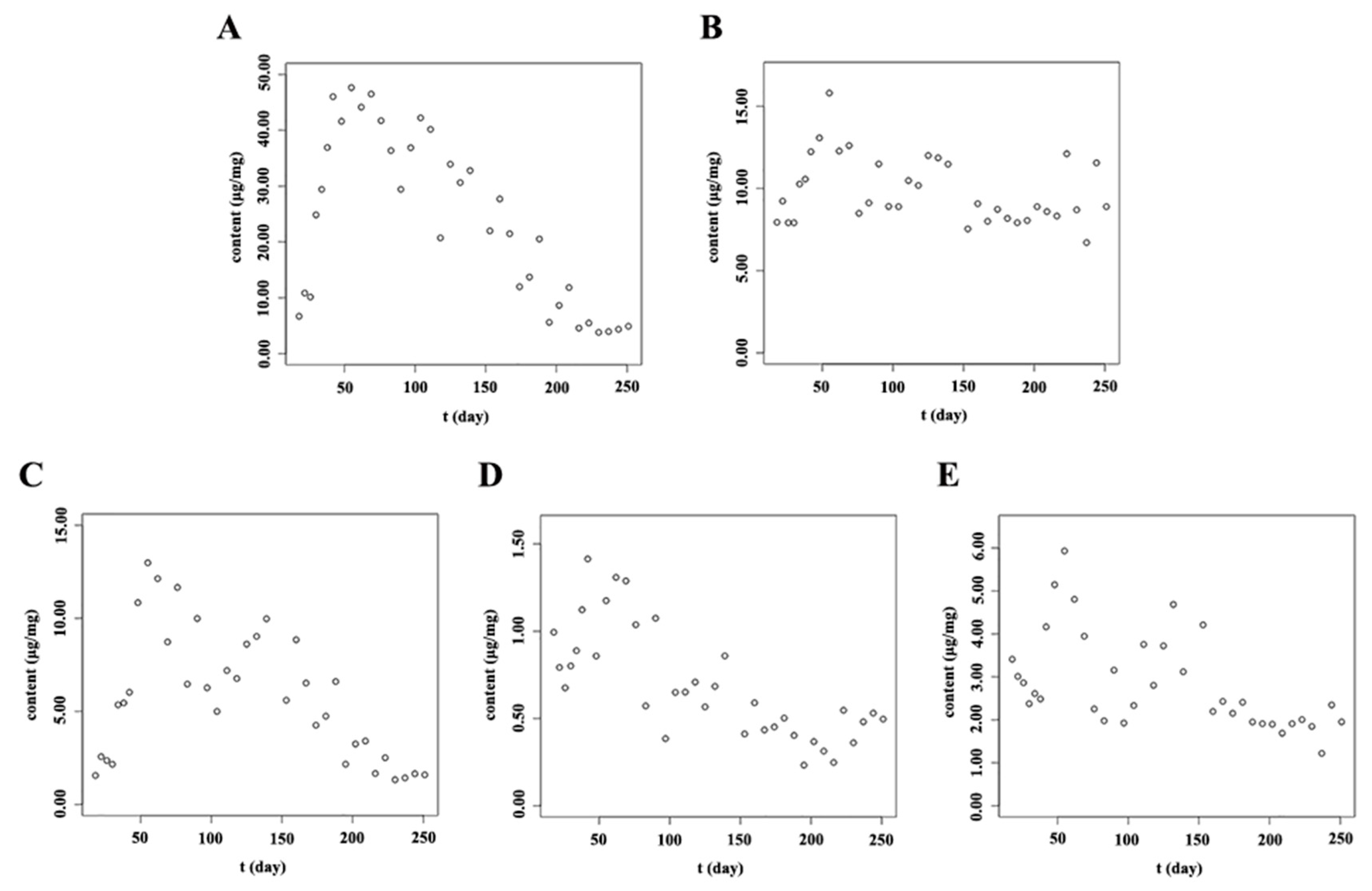
2.4. Dynamic Accumulation of Flavonoids in Stems and Leaves of S. baicalensis at Different Growth Stages
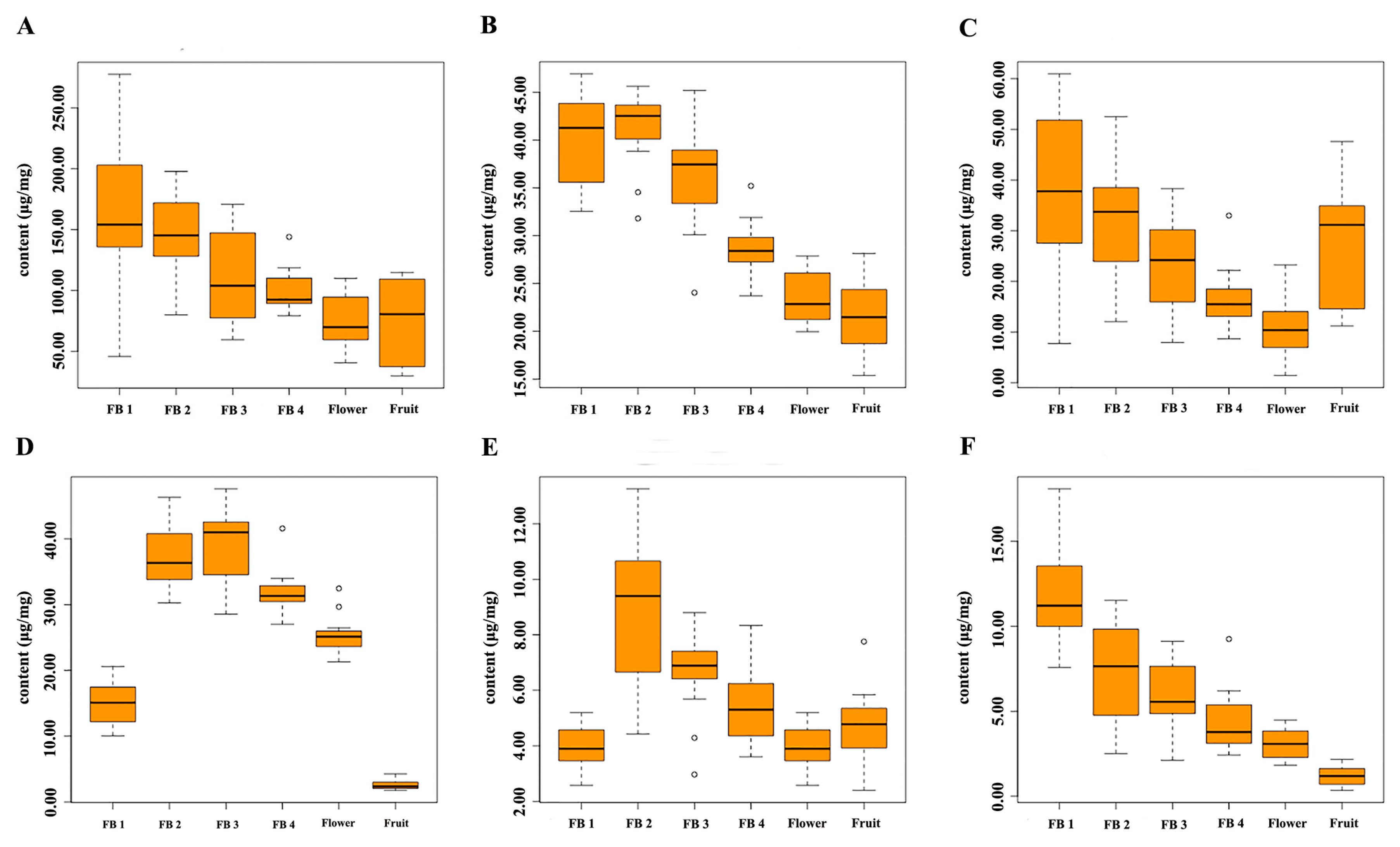
2.5. Dynamic Accumulation of Flavonoids in Reproductive Organs of S. baicalensis at Different Growth Stages
3. Materials and Methods
3.1. Plant Materials and Sample Preparation
3.2. Chemicals and Reagents
3.3. UHPLC-QTOF-MS/MS Based Qualitative Analysis and HPLC Quantification
3.4. Method Validation
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; Chinese Medical Science and Technology Press: Beijing, China, 2015; Volume 1, pp. 301–302. [Google Scholar]
- Shang, X.F.; He, X.R.; He, X.Y.; Li, M.X.; Zhang, R.X.; Fan, P.C.; Zhang, Q.L.; Jia, Z.P. The genus Scutellaria an ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2010, 128, 279–313. [Google Scholar] [CrossRef] [PubMed]
- Saralamma, V.V.G.; Lee, H.J.; Hong, G.E.; Park, H.S.; Yumnam, S.; Raha, S.; Lee, W.S.; Kim, E.H.; Sung, N.J.; Lee, S.J.; et al. Korean Scutellaria baicalensis Georgi flavonoid extract induces mitochondrially mediated apoptosis in human gastric cancer AGS cells. Oncol. Lett. 2017, 14, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.L.; Wang, P.W.; Leu, Y.L.; Wu, T.H.; Wu, T.S. Inhibitory effects of Scutellaria baicalensis extract on hepatic stellate cells through inducing G2/M cell cycle arrest and activating ERK-dependent apoptosis via Bax and caspase pathway. J. Ethnopharmacol. 2012, 139, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, Y.Z.; Brooks, J.D.; Corke, H. The in vitro antibacterial activity of dietary spice and medicinal herb extracts. Int. J. Food Microbiol. 2007, 117, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Li, R.; Song, W.; Miao, W.J.; Liu, J.; Chen, H.B.; Guo, D.A.; Ye, M. A targeted strategy to analyze untargeted mass spectral data: Rapid chemical profiling of Scutellaria baicalensis using ultra-high performance liquid chromatography coupled with hybrid quadrupole orbitrap mass spectrometry and key ion filtering. J. Chromatogr. A 2016, 1441, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Seo, O.N.; Kim, G.S.; Kim, Y.H.; Park, S.; Jeong, S.W.; Lee, S.J.; Jin, J.S.; Sung, C.S. Determination of polyphenol components of Korean Scutellaria baicalensis Georgi using liquid chromatography–tandem mass spectrometry: Contribution to overall antioxidant activity. J. Funct. Foods 2013, 5, 1741–1750. [Google Scholar] [CrossRef]
- Zhao, T.H.; Chen, S.P.; Yang, H.S.; Deng, S.H. Antiviral effects of active fraction from stems and leaves of Scutellaria baicalensis. J. China Pharm. Univ. 2006, 37, 544–547. [Google Scholar]
- Li, S.T.; Yang, H.M.; Shi, Y.H.; Gong, M.Y. Protective effects of Scutellaria baicalensis stem-leaf total flavonoid on carbon tetrachloride acute hepatic injury in mice. J. Chengde Med. Coll. 2003, 20, 292–294. [Google Scholar]
- Gao, Z.; Huang, K.; Xu, H. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in HS-SY5Y cells. Pharmacol. Res. 2001, 43, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.Y.; Zhao, S.M. Protective effects of preconditioning of total fiavonoid from the stem and leaf of Scutellaria baicalensis on cardiocyte ultrastructure of rats with ischemic reperfusion. Chin. J. Clin. Rehabil. 2006, 10, 49–51. [Google Scholar]
- Liu, G.; Rajesh, N.; Wang, X.; Zhang, M.; Wu, Q.; Li, S.; Chen, B.; Yao, S. Identification of flavonoids in the stems and leaves of Scutellaria baicalensis Georgi. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, J.; Wu, J.; Qi, F.; Wang, H.; Wang, Z.; Xu, Z. Scutellarin protects cardiomyocyte ischemia-reperfusion injury by reducing apoptosis and oxidative stress. Life Sci. 2016, 157, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Zhang, X.-M.; Jiang, J.-G.; Zhu, W. Apigenin-7-O-β-d-glucuronide inhibits modified low-density lipoprotein uptake and foam cell formation in macrophages. J. Funct. Foods 2017, 36, 615–621. [Google Scholar] [CrossRef]
- Wang, H.; Cao, J.; Xu, S.; Gu, D.; Wang, Y.; Xiao, S. Depletion of high-abundance flavonoids by metal complexation and identification of low-abundance flavonoids in Scutellaria baicalensis Georgi. J. Chromatogr. A 2013, 1315, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Moco, S.; Capanoglu, E.; Tikunov, Y.; Bino, R.J.; Boyacioglu, D.; Hall, R.D.; Vervoort, J.; Devos, R.C. Tissue specialization at the metabolite level is perceived during the development of tomato fruit. J. Exp. Bot. 2007, 58, 4131–4146. [Google Scholar] [CrossRef] [PubMed]
- Abu-Reidah, I.M.; Del, M.C.M.; Arráez-Román, D.; Fernández-Gutiérrez, A.; Segura-Carretero, A. UHPLC-ESI-QTOF-MS-based metabolic profiling of Vicia faba L. (Fabaceae) seeds as a key strategy for characterization in foodomics. Electrophoresis 2014, 35, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Álvarezfernández, M.A.; Hornedoortega, R.; Cerezo, A.B.; Troncoso, A.M.; García-Parrilla, M.C. Determination of nonanthocyanin phenolic compounds using High-Resolution Mass Spectrometry (UHPLC-Orbitrap-MS/MS) and impact of storage conditions in a beverage made from Strawberry by fermentation. J. Agric. Food Chem. 2016, 64, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Erşan, S.; Üstündağ, Ö.G.; Carle, R.; Schweiggert, R.M. Determination of pistachio (Pistacia vera L.) hull (exo- and mesocarp) phenolics by HPLC-DAD-ESI/MS n and UHPLC-DAD-ELSD after ultrasound-assisted extraction. J. Food Compos. Anal. 2017, 62, 103–114. [Google Scholar] [CrossRef]
- Zhang, L.; Tu, Z.C.; Xie, X.; Wang, H.; Wang, H.; Wang, Z.X.; Sha, X.M.; Lu, Y. Jackfruit (Artocarpus heterophyllus Lam.) peel: A better source of antioxidants and α-glucosidase inhibitors than pulp, flake and seed, and phytochemical profile by HPLC-QTOF-MS/MS. Food Chem. 2017, 234, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Johnson-Cicalese, J.; Singh, A.P.; Vorsa, N. Characterization and quantification of flavonoids and organic acids over fruit development in American cranberry (Vaccinium macrocarpon) cultivars using HPLC and APCI-MS/MS. Plant Sci. 2017, 262, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.L.; Shen, C.C.; Shen, Y.C.; Chiou, W.F.; Chen, C.C. Anti-inflammatory and antiosteoporosis flavonoids from the rhizomes of Helminthostachys zeylanica. J. Nat. Prod. 2017, 80, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, X.; Qi, J.; Yu, B. Simultaneous qualitative and quantitative analysis of flavonoids and alkaloids from the leaves of Nelumbo nucifera Gaertn. using high-performanceliquid chromatography with quadrupole time-of-flight mass spectrometry. J. Sep. Sci. 2016, 39, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhang, Z.; Lu, L.; Liu, Y.; Li, S.; Wang, J.; Song, Z.; Yan, Z.; Miao, J. Rapid identification and quantitative analysis of the chemical constituents in Scutellaria indica L. by UHPLC-QTOF-MS and UHPLC-MS/MS. J. Pharm. Biomed. Anal. 2016, 117, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; He, L.; Lu, L.; Liu, Y.; Dong, G.; Miao, J.; Luo, P. Characterization and quantification of the chemical compositions of Scutellariae barbatae herba and differentiation from its substitute by combining UHPLC-PDA-QTOF-MS/MS with UHPLC-MS/MS. J. Pharm. Biomed. Anal. 2015, 109, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Liu, S.; Wu, B. Structural identification of chemical constituents from Scutellaria baicalensis by HPLC-ESI-MS/MS and NMR spectroscopy. Asian J. Chem. 2013, 25, 3799–3805. [Google Scholar] [CrossRef]
- Liu, G.; Ma, J.; Chen, Y.; Tian, Q.; Shen, Y.; Wang, X.; Chen, B.; Yao, S. Investigation of flavonoid profile of Scutellaria bacalensis Georgi by high performance liquid chromatography with diode array detection and electrospray ion trap mass spectrometry. J. Chromatogr. A 2009, 1216, 4809–4814. [Google Scholar] [CrossRef] [PubMed]
- Yukinori, M.; Yoshitaka, I.; Hiroyuki, S.; Tsuyoshi, T. Studies on the constituents of Seutellaria species (X) On the flavonoid constituents of the leaves of Scutellaria baicalensis Gergi. Shoyakugaku Zasshi 1988, 42, 216–219. [Google Scholar]
- Wang, L.; Zhu, X.; Lou, X.; Zheng, F.; Feng, Y.; Liu, W.; Feng, F.; Xie, N. Systematic characterization and simultaneous quantification of the multiple components of Rhododendron dauricum based on high-performance liquid chromatography with quadrupole time-of-flight tandem mass spectrometry. J. Sep. Sci. 2015, 38, 3161–3169. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Downey, F.; Ng, K.Y. Comprehensive profiling of flavonoids in Scutellaria incana L. using LC-Q-TOF-MS. Acta Chromatogr. 2013, 25, 555–569. [Google Scholar] [CrossRef]
- Zheng, L.; Cong, H.J.; Wu, B.; Xue, M.; Xiang, T.; Yao, Z.Q.; Lin, W.H. HPLC-Q-TOF-MS/MS Analysis of the Constituents in the Rat Biological Fluids After Oral Administration of Qing Ru Xiao granules. J. Chromatogr. Sci. 2015, 53, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.P.; Wei, H.Q. An overview of the chemical constituents of Scutellaria. Drugs Clin. 1994, 9, 147–156. [Google Scholar]
- Xi, Y.L.; Shang, Y.Z. Research progress of the leaf and stem of Scutellaria baicalensis. J. Chengde Med. Coll. 2007, 24, 293–295. [Google Scholar]
- Pourcel, L.; Routaboul, J.M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [PubMed]
- Auger, B.; Marnet, N.; Gautier, V.; Maia-Grondard, A.; Leprince, F.; Renard, M.; Guyot, S.; Nesi, N.; Routaboul, J.M. A Detailed Survey of Seed Coat Flavonoids in Developing Seeds of Brassica napus L. J. Agric. Food Chem. 2010, 58, 6246–6256. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhang, Y.; Wang, G.; Hill, L.; Weng, J.K.; Chen, X.Y.; Xue, H.; Martin, C. A specialized flavone biosynthetic pathway has evolved in the medicinal plant, Scutellaria baicalensis. Sci. Adv. 2016, 2, e1501780. [Google Scholar] [CrossRef] [PubMed]
- Winsor, C.P. The gompertz curve as a growth curve. Proc. Natl. Acad. Sci. USA 1932, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Li, Q.B.; Yang, T.W.; Nie, Z.N.; Chen, G.X.; Hu, L.Y. Responses of plant development, biomass and seed production of direct sown oilseed rape (Brassica napus) to nitrogen application at different stages in Yangtze River Basin. Field Crop. Res. 2016, 194, 12–20. [Google Scholar] [CrossRef]
- Li, J.; Oulee, T.M.; Raba, R.; Amundson, R.G.; Last, R.L. Arabidopsis flavonoid mutants are hypersensitive to UV-B irradiation. Plant Cell 1993, 5, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Bieza, K.; Lois, R. An arabidopsis mutant tolerant to lethal Ultraviolet-B levels shows constitutively elevated accumulation of flavonoids and other phenolics. Plant Physiol. 2001, 126, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Wang, L.M.; Li, M.T.; Yan, Y.Y.; Ao, M.Z.; Wu, G.; Yu, L.J. Influence of flowering stage of Lonicera japonica thunb. on variation in volatiles and chlorogenic acid. J. Sci. Food Agric. 2009, 89, 953–957. [Google Scholar] [CrossRef]
- Zhu, W.; Zheng, W.; Hu, X.; Xu, X.; Zhang, L.; Tian, J. Variations of metabolites and proteome in Lonicera japonica thunb. buds and flowers under UV radiation. Biochim. Biophys. Acta 2017, 1865, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Horvath, C.R.; Martos, P.A.; Saxena, P.K. Identification and quantification of eight flavones in root and shoot tissues of the medicinal plant huang-qin (Scutellaria baicalensis Georgi) using high-performance liquid chromatography with diode array and mass spectrometric detection. J. Chromatogr. A 2005, 1062, 199–207. [Google Scholar] [CrossRef] [PubMed]
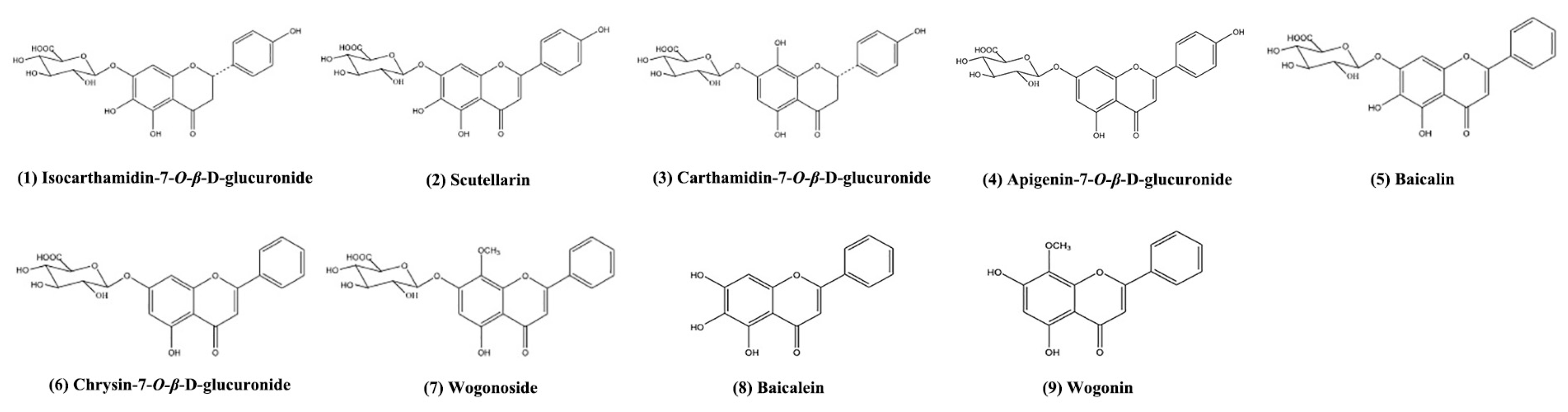
Sample Availability: Samples of the compounds including isocarthamidin-7-O-β-d-glucuronide, scutellarin, carthamidin-7-O-β-d-glucuronide, apigenin-7-O-β-d-glucuronide, baicalin, chrysin-7-O-β-d-glucuronide, wogonoside, baicalein and wogonin are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | tR(min) | Molecular Formula | [M − H]− Predicted | [M − H]− Measured | Error (ppm) | (−)-ESI-MS/MS Fragment Ions (m/z) | Identification [Reference] |
|---|---|---|---|---|---|---|---|
| 1 | 1.59 | C21H22O13 | 481.09876 | 481.09832 | −0.9 | 463.0880, 319.0452, 301.0345, 257.0457, 233.0465, 215.0337, 193.0141, 175.0039,151.0043, 125.0254 | Dihydromyricetin-3′-O-glucoside [17] |
| 2 | 2.24 | C21H22O13 | 481.09876 | 481.09866 | −0.2 | 301.0349, 257.0448, 215.0344, 193.0142, 175.0045, 151.0045, 125.0254, 137.0251 | Isomer of dihydromyricetin-3′-O-glucoside |
| 3 | 2.43 | C21H22O12 | 465.10385 | 465.10158 | −4.9 | 303.0398, 285.0396, 273.0295, 241.0461, 219.0260, 177.0181, 125.0267 | Taxifolin-7-O-glucoside [18] |
| 4 | 3.5 | C21H22O12 | 465.10385 | 465.10313 | −1.6 | 303.0505, 285.0398, 275.0551, 259.0600, 241.0511, 217.0509, 125.0254 | Isomer of Taxifolin-7-O-glucoside |
| 5 | 4.43 | C21H22O12 | 465.10385 | 465.10323 | −1.3 | 303.0504, 285.0388, 241.0522, 199.0364, 179.0001, 149.0248, 125.0243 | Isomer of Taxifolin-7-O-glucoside |
| 6 | 4.84 | C15H12O9 | 335.04086 | 335.04074 | −0.4 | 183.0294, 169.0135, 139.0404, 125.0252, 115.0405 | Methyl digallate [19] |
| 7 | 5.27 | C21H22O13 | 481.09876 | 481.09846 | −0.6 | 463.0893, 301.0352, 283.0247, 257.0551, 255.0291, 215.03.52, 193.0142, 162.0359, 153.0198, 125.0253 | Isomer of dihydromyricetin-3′-O-glucoside |
| 8 | 5.69 | C22H24O12 | 479.11950 | 479.11847 | −2.2 | 317.0648, 285.0444, 165.0204 | 3,5,7,4′-Tetrahydroxy-6-methoxy Flavanone 3-O-β-d-glucoside |
| 9 | 7.75 | C15H12O8 | 319.04594 | 319.04568 | −0.8 | 233.0475, 215.0341, 193.0138, 175.0034, 165.0189, 151.0037, 137.0246, 125.0253 | Dihydromyricetin [20] |
| 10 | 8.40 | C21H20O13 | 479.08311 | 479.08288 | −0.5 | 317.0294, 289.0319, 287.0181, 243.0296, 178.9981, 151.0033, 137.0250 | Myricetin-3-O-Galactoside [21] |
| 11 | 8.74 | C21H20O11 | 447.09329 | 447.09334 | 0.1 | 285.0460, 217.0147,151.0035 | 5,7,2′,6′-Tetrahydroxy Flavone 2′-O-β-d-glucoside [6] |
| 12 | 9.05 | C15H12O7 | 303.05103 | 303.05052 | −1.7 | 125.0256, 149.0231, 177.0176, 267.0288 | 3,5,7,2′,6′-Pentahydroxy Flavanone [6] |
| 13 | 9.17 | C21H22O12 | 465.10385 | 465.10367 | −0.4 | 303.0505, 285.0408, 275.0556, 259.0609, 241.0498, 217.0511, 199.0389, 177.0196, 125.0250 | Isomer of Taxifolin-7-O-glucoside |
| 14 | 9.4 | C15H12O8 | 319.04594 | 319.04571 | −0.7 | 215.0349, 193.0139, 175.0037, 165.0189, 151.0039, 137.0249, 125.0249 | Isomer of dihydromyricetin |
| 15 | 9.63 | C27H30O17 | 625.14102 | 625.14242 | 2.2 | 463.0871, 300.0274, 271.0245, 255.0309, 243.0309, 178.9969 | Quercetin-4′-O-β-d-glucopyranosyl-(1→2)-β-d-glucopyranoside [22] |
| 16 | 9.67 | C21H20O13 | 479.08311 | 479.08305 | −0.1 | 303.0506, 285.0384, 181.0138, 166.9992, 135.0463, 113.0277 | 5,6,7,3′,4′-Pentahydroxy Flavanon 7-O-β-d-glucoronide [12] |
| 17 | 9.88 | C26H28O14 | 563.14063 | 563.13869 | −3.4 | 503.1135, 473.1102, 443.1020, 383.0779, 353.0672 | Schaftoside [15] |
| 18 | 9.92 | C26H28O14 | 563.14063 | 563.14154 | 1.6 | 503.1244, 473.1127, 443.0978, 383.0791, 353.0662 | Isoschaftoside [15] |
| 19 | 9.98 | C21H20O11 | 447.09329 | 447.09289 | −0.9 | 285.0403, 284.0319, 240.0448, 227.0302, 228.0412 | Kaempferol-3-O-glucoside [23] |
| 20 | 10.29 | C21H22O11 | 449.10894 | 449.10785 | −2.4 | 125.0252, 151.0029, 161.0244, 177.0204, 227.0360, 269.0466, 287.0558 | 5,7,2′,6′-TetrahydroxyFlavanone 2′-O-β-d-glucoside [6] |
| 21 | 10.46 | C22H22O12 | 477.10385 | 477.10258 | −2.7 | 301.0349, 283.0264, 164.9816 | 6-Hydroxyluteolin 7-O-β-d-glucoronide [6] |
| 22 | 11.04 | C21H22O12 | 465.10385 | 465.10384 | 0 | 303.0872, 285.0765, 259.0977, 244.0734, 137.0611 | Isomer of Taxifolin-7-O-glucoside |
| 23 | 11.16 | C27H30O16 | 609.14611 | 609.14694 | 1.4 | 489.1024, 447.0911, 429.0817, 284.0321, 255.0295, 227.0340, 211.0394, 178.9980, 151.0031 | Unknown |
| 24 | 12.13 | C26H28O13 | 547.14571 | 547.14391 | −3.3 | 487.1257, 457.1136, 427.1046, 367.0819, 337.0717 | Chrysin 6-C-arabinoside-8-C-glucoside [7] |
| 25 | 12.66 | C21H20O12 | 463.08820 | 463.08660 | −3.5 | 287.0560, 269.0541, 259.0602, 181.0142, 166.9993, 153.0201, 119.0517 | Isocarthamidin -7-O-β-d-glucuronide [24] |
| 26 | 12.91 | C21H18O12 | 461.07255 | 461.07073 | −3.9 | 285.0395, 257.0435, 164.9827, 136.9883, 119.0517 | Luteolin 7-O-glucuronide [25] |
| 27 | 13.27 | C21H20O11 | 447.09329 | 447.09284 | −1 | 285.0390, 284.0314, 271.0611, 256.0293 | Isomer of Kaempferol-3-O-glucoside |
| 28 | 13.47 | C23H24O13 | 507.11441 | 507.11317 | −2.4 | 345.0617, 330.0379, 315.0150 | Viscidulin III 6′-O-β-d-glucoside [6] |
| 29 | 13.59 | C21H18O12 | 461.07255 | 461.07162 | −2.0 | 285.0401, 267.0292, 239.0363, 213.0548, 195.0455 | Scutellarin [24] |
| 30 | 14.00 | C29H36O15 | 623.19814 | 623.19657 | −2.5 | 461.1696, 161.0239 | Acteoside [6] |
| 31 | 14.13 | C26H28O13 | 547.14571 | 547.14518 | −1.0 | 457.1140, 427.1040, 367.0822, 337.0720 | Chrysin 6-C-glucoside-8-C-arabinoside [15] |
| 32 | 14.84 | C21H20O12 | 463.08820 | 463.08735 | −1.8 | 287.0542, 269.0453, 259.0616, 193.0147, 181.0147, 166.0021, 153.0203, 119.0520 | carthamidin -7-O-β-d-glucuronide [24] |
| 33 | 15.55 | C23H24O12 | 491.11950 | 491.11730 | −4.5 | 329.0659, 314.0432, 299.0189 | 5,2′,6′-Dihydroxy-7,8-dimethoxy Flavone 2′-O-β-d-glucoside [6] |
| 34 | 17.07 | C21H18O12 | 461.07255 | 461.07033 | −4.8 | 285.0408, 257.0463, 229.0451, 241.0514, 213.0581, 199.0387, 185.0590, 113.0268 | Kaempferol-3-O-β-d-glucuronide [26] |
| 35 | 17.17 | C21H20O9 | 415.10346 | 415.10199 | −3.5 | 325.0714, 295.0610, 267.0661 | Chrysin 8-C-β-d-glucoside [6] |
| 36 | 17.25 | C21H20O10 | 431.09837 | 431.09691 | −3.4 | 269.0447 | Apigenin-7-O-β-d-glucoside [25] |
| 37 | 17.43 | C22H20O12 | 475.08820 | 475.08685 | −2.8 | 299.0549, 271.0641, 256.0365, 227.0761, 165.0218 | 5,6,7-Trihydroxy-8-methoxy-7-O-β-d-glucuronide [27] |
| 38 | 17.53 | C21H20O11 | 447.09329 | 447.09216 | −2.5 | 285.0393, 284.0321, 269.0411, 257.0456, 255.0291, 227.0324, 213.0568, 151.0042, 107.0157 | Isomer of Kaempferol-3-O-glucoside |
| 39 | 17.64 | C24H26O13 | 521.13006 | 521.12836 | −3.3 | 359.0769, 344.0544, 329.0311, 314.0057 | 5,2′,6′-Trihydroxy-6,7,8,-trimethoxy Flavone 2′-O-β-d-glucoside [2] |
| 40 | 17.79 | C15H10O6 | 285.04046 | 285.04014 | −1.1 | 199.0388, 151.0054 | 5,7,2′,6′-Tetrahydroxy Flavone [6] |
| 41 | 17.90 | C15H12O6 | 287.05611 | 287.05510 | −3.5 | 259.0661, 177.0546, 125.0245 | Carthamidin [25] |
| 42 | 17.98 | C21H18O11 | 445.07764 | 445.07600 | −3.7 | 269.0462, 225.0563, 113.0261 | Apigenin-7-O-β-d-glucuronide [12] |
| 43 | 18.07 | C21H20O11 | 447.09239 | 447.09218 | −2.5 | 271.0604, 177.0223, 151.0043, 119.0516, 113.0261, 107.0160 | Naringenin-7-O-β-d-glucuronide [7] |
| 44 | 18.29 | C21H20O12 | 463.08820 | 463.08743 | −1.7 | 287.0570, 269.0420, 259.0658, 193.0149, 181.0149, 166.9982, 153.0196, 139.0041, 119.0531 | Eriodictyol-7-O-β-d-glucuronide [25] |
| 45 | 18.38 | C15H12O6 | 287.05611 | 287.05516 | −3.3 | 181.0142, 153.0188, 166.9973, 139.0038, 136.9891, 119.0497 | Isocarthamidin [28] |
| 46 | 18.59 | C15H10O8 | 317.03029 | 317.02999 | −1 | 289.0304, 271.0231, 261.0358, 243.0274, 227.0334, 193.0143, 178.9985, 165.0192, 151.0044, 137.0254, 125.0246, 109.0310, 107.0148 | Myricetin [29] |
| 47 | 18.64 | C22H20O12 | 475.08820 | 475.08692 | −2.7 | 299.0560, 284.0314 | 5,7,8-Trihydroxy-6-methoxy Flavone-7-O-β-d-glucuronide [30] |
| 48 | 18.68 | C21H18O12 | 461.07255 | 461.07194 | −1.3 | 285.0353, 139.0017, 257.0466, 164.9834, 136.9886, 119.0510 | Isoscutellarein 8-O-β-d-glucuronide [12] |
| 49 | 18.98 | C22H22O12 | 477.10385 | 477.10144 | −5.0 | 331.0311, 301.0334, 180.9795, 155.0067, 119.0530 | 5,7,2′-Trihydroxy-6-methoxy Flavanone 7-O-β-d-glucuronoide [15] |
| 50 | 19.01 | C22H22O12 | 477.10385 | 477.10234 | −3.2 | 301.0697, 286.0475, 181.0148 | 5,7,2′-Trihydroxy-8-methoxyFlavanone 7-O-β-d-glucuronoide [15] |
| 51 | 20.28 | C21H20O11 | 447.09329 | 447.09211 | −2.6 | 285.0397, 267.0289, 241.0539, 199.0371, 151.0062 | Isomer of Kaempferol-3-O-glucoside |
| 52 | 20.58 | C16H14O7 | 315.05103 | 315.05069 | −1.1 | 300.0276, 272.0319, 244.0378, 216.0419, 187.0398, 180.9781, 152.9843, 124.9893 | Pedalitin |
| 53 | 20.67 | C15H10O6 | 285.04046 | 285.04042 | −0.1 | 239.0327, 167.0010, 137.0260, 117.0353 | Scutellarein [25] |
| 54 | 21.17 | C23H24O12 | 491.11950 | 491.11970 | 0.4 | 329.0635, 314.0426, 299.0252 | 5,2′,6′-Dihydroxy-6,7-dimethoxy Flavone 2′-O-β-d-glucoside [30] |
| 55 | 21.28 | C21H20O10 | 431.09837 | 431.09702 | −3.1 | 269.0445, 251.0354, 195.0465, 167.0506 | Baicalein 7-O-β-d-glucoside [6] |
| 56 | 21.36 | C21H18O11 | 445.07764 | 445.07622 | −3.2 | 269.0456, 251.0345, 223.0397, 197.0621, 113.0262 | Baicalin [12] |
| 57 | 21.70 | C15H10O5 | 269.04555 | 269.04489 | −2.4 | 251.0376, 241.0485, 223.0416, 195.0458, 139.0061, 111.0072 | Islandicin |
| 58 | 22.08 | C15H10O6 | 285.04046 | 285.03977 | −2.4 | 241.0509, 213.0540, 195.0482, 239.0333 | Isoscutellarein [26] |
| 59 | 22.68 | C21H18O12 | 461.07255 | 461.07031 | −4.9 | 285.0415, 267.0285, 239.0317 | Isoscutellarein-7-O-β-d-glucuronide [12] |
| 60 | 23.50 | C31H40O15 | 651.22944 | 651.22685 | −4.0 | 475.1832, 193.0499, 175.0398, 160.0171 | Cistanoside D [6] |
| 61 | 24.31 | C22H20O12 | 475.08820 | 475.08707 | −2.4 | 299.0565, 284.0332 | 5,7,2′-Trihydroxy-6-methoxy Flavone 7-O-β-d-glucuronoide [2] |
| 62 | 24.37 | C21H20O11 | 447.09329 | 447.09150 | −4.0 | 271.0610, 243.0652, 113.0237 | Dihydrobaicalin [6] |
| 63 | 24.41 | C22H20O12 | 475.08820 | 475.08747 | −1.5 | 460.1019, 299.0550, 297.0401, 284.0195, 282.0195, 254.0195 | Isomer of cirsimaritin 4′-glucoside |
| 64 | 24.72 | C16H14O6 | 301.07176 | 301.07067 | −3.6 | 286.0474, 181.0144, 165.9916, 137.9972, 119.0515, 110.0026 | 5,7,4′-Trihydroxy-6-methoxy Flavanone [15] |
| 65 | 25.56 | C21H18O11 | 445.07764 | 445.07576 | −4.2 | 269.0449, 241.0494, 225.0556, 197.0611, 171.0451 | Norwogonin 7-O-β-d-glucuronide [6] |
| 66 | 25.86 | C22H22O12 | 477.10385 | 477.10214 | −3.6 | 301.0712, 283.0620, 273.0774, 268.0393, 139.0037, 113.0262 | 5,7,2′,6′-Tetrahydroxy flavonol 7-O-β-d-glucuronoide [26] |
| 67 | 26.22 | C22H22O11 | 461.10894 | 461.10732 | −3.5 | 299.0536, 284.0328, 171.0449 | 5,7,2′-Trihydroxy-6-methoxy Flavone 7-O-β-d-glucoside [6] |
| 68 | 27.11 | C21H18O11 | 445.07764 | 445.07556 | −4.7 | 269.0450, 241.0485, 225.0560, 171.0472 | Norwogonin 8-O-β-d-glucuronide [6] |
| 69 | 27.76 | C21H18O10 | 429.08272 | 429.08064 | −4.8 | 253.0499, 209.0601,143.0503,113.0260 | Chrysin 7-O-β-d-glucuronide [24] |
| 70 | 27.96 | C22H20O11 | 459.09329 | 459.09148 | −3.9 | 283.0612, 268.0376 | Oroxylin A 7-O-β-d-glucuronide [6] |
| 71 | 28.68 | C22H20O12 | 475.0882 | 475.08663 | −3.3 | 299.0549, 284.0320 | Isomer of cirsimaritin 4′-glucoside |
| 72 | 28.87 | C17H14O7 | 329.06668 | 329.06548 | −3.6 | 314.0451, 299.0185, 271.0237, 164.9808, 136.9872 | 5,2′,6′-Trihydroxy-7,8-dimethoxy Flavone [2] |
| 73 | 29.64 | C16H14O6 | 301.07176 | 301.07086 | −3.0 | 286.0459, 181.0153, 165.9924, 137.9976, 119.0521, 110.0027 | 5,7,4′-Trihydroxy-8-methoxy Flavanone [15] |
| 74 | 29.84 | C21H18O11 | 445.07764 | 445.07594 | −3.8 | 269.0455, 251.0341, 223.0383, 113.0273 | Baicalein 6-O-β-d-glucuronide [6] |
| 75 | 30.6 | C15H10O6 | 285.04046 | 285.03945 | −3.6 | 151.0030, 107.0150 | Luteolin [12] |
| 76 | 30.81 | C22H20O11 | 459.09329 | 459.09140 | −4.1 | 283.0613, 268.0375 | Wogonoside [12] |
| 77 | 31.34 | C23H24O11 | 475.12459 | 475.12307 | −3.2 | 460.0994, 445.0748, 313.0665, 297.0390, 283.0199, 282.0144, 254.0212, 226.0276, 183.0437 | Cirsimaritin 4′-O-glucoside [31] |
| 78 | 32.05 | C21H20O10 | 431.09837 | 431.09633 | −4.7 | 255.0674, 213.0562, 187.0791, 151.0039, 113.0259 | 5,7-dihydroxy Flavanone 7-O-β-d-glucuronoide [12] |
| 79 | 32.65 | C23H22O12 | 489.10385 | 489.10194 | −3.9 | 313.0712, 298.0485, 283.0243, 255.0361, 211.0416, 113.0274 | 5,7-Dihydroxy-2′,8-dimethoxy Flavone 7-O-β-d-glucuronide [24] |
| 80 | 33.30 | C18H16O8 | 359.07724 | 359.07546 | −4.9 | 344.0565, 329.0297, 314.0125, 286.0098, 194.9928 | 5,2′,5′-Trihydroxy-6,7,8-trimethoxy Flavone [2] |
| 81 | 34.84 | C16H12O6 | 299.05611 | 299.05566 | −1.5 | 284.0323, 255.0232, 227.0358, 211.0477, 183.0436, 164.0130, 136.9906 | Isomer of 5,7,4′-Trihydroxy-8-methoxy Flavone |
| 82 | 34.91 | C15H10O5 | 269.04555 | 269.04478 | −2.9 | 241.0454, 225.0531, 195.0443, 171.0439, 117.0358 | Apigenin [24] |
| 83 | 36.07 | C17H14O7 | 329.06668 | 329.06584 | −2.5 | 314.0461, 299.0201, 227.0325, 165.9927, 137.9951, 110.0020 | 5,8,2′-Trihydroxy-6,7-dimethoxy Flavone [6] |
| 84 | 36.28 | C17H14O6 | 313.07176 | 313.07085 | −2.9 | 298.0542, 283.0251, 244.8595, 211.0396, 166.8658, 155.0508 | 5,8-Dihydroxy-6,7-dimethoxy Flavone [30] |
| 85 | 36.59 | C16H12O6 | 299.05611 | 299.05552 | −2.0 | 284.0318, 255.0274, 239.0370, 171.0454, 153.9920, 125.9966 | 5,7,4′-Trihydroxy-8-methoxy Flavone [30] |
| 86 | 37.11 | C18H16O7 | 343.08233 | 343.08170 | −1.8 | 328.0591, 313.0331, 298.0112, 270.0167, 241.0502, 226.0271, 198.0335, 185.0629, 155.0521 | SkullcapFlavone I [27] |
| 87 | 37.16 | C15H10O5 | 269.04555 | 269.04510 | −1.7 | 251.0348, 241.0499, 223.0399, 195.0442, 169.0660, 136.9880 | Baicalein [6] |
| 88 | 37.54 | C15H12O5 | 271.06120 | 271.06073 | −1.7 | 243.0638, 225.0535, 185.0601, 152.0182, 139.0070, 124.0182 | Dihydronorwogonin [2] |
| 89 | 37.67 | C17H14O7 | 329.06668 | 329.06602 | −2.0 | 314.0367, 299.0139 | 5,7,2′-Trihydroxy-8,6′-dimethoxy Flavone [32] |
| 90 | 38.30 | C16H12O6 | 299.05611 | 299.05531 | −2.7 | 284.0312, 165.9925 | 5,6,7-Trihydroxy-4′-methoxy Flavone [33] |
| 91 | 44.26 | C18H16O7 | 343.08233 | 343.08158 | −2.2 | 328.0584, 313.0347, 298.0130, 285.0400, 164.9831 | SkullcapFlavone [6] |
| 92 | 44.65 | C16H12O5 | 283.06120 | 283.06097 | −0.8 | 268.0371, 239.0353, 211.0401, 184.0532, 163.0037, 110.0021 | Wogonin [6] |
| 93 | 45.47 | C15H10O4 | 253.05063 | 253.05033 | −1.2 | 209.0616, 166.8665, 143.0516, 107.0165 | Chrysin [24] |
| 94 | 46.31 | C19H18O8 | 373.09289 | 373.09147 | −3.8 | 358.0616, 343.0434, 328.0199, 300.0248 | 5,6′-Dihydroxy-6,7,8,2′-tetramethoxy Flavone [6] |
| 95 | 46.59 | C16H12O5 | 283.06120 | 283.06056 | −2.3 | 268.0373, 239.0358, 211.0363, 184.0525, 110.0016 | Oroxylin A [6] |
| 96 | 46.66 | C15H12O4 | 255.06628 | 255.06670 | 1.6 | 213.0545, 201.8339, 182.9022, 166.8653 | Pinocembrin [12] |
| 97 | 47.20 | C17H14O6 | 313.07176 | 313.07047 | −4.1 | 298.0505, 283.0256, 255.0307, 183.0454, 164.9835 | 5,7-Dihydroxy-6,8-dimethoxy Flavone [27] |
| 98 | 49.45 | C18H16O7 | 343.08233 | 343.08130 | −3.0 | 328.0598, 313.0346, 298.0124, 270.0161 | Tenaxin I [6] |
| Number of the Compound | |
|---|---|
| Different growth stage | |
| Seed stage | 83 |
| Seedling stage | 37, 59, 67, 72, 75, 77, 80, 97 |
| Mature stage | 3, 8, 48, 88 |
| Different Organ | |
| Root | 11, 12, 18, 28, 35, 39, 70, 74, 79, 85, 91, 98 |
| Stem | 60, 78 |
| Leaf | 40 |
| Reproductive organs | 2, 3, 4, 6, 7, 8, 10, 11, 12, 14, 15, 23, 36, 38, 41, 46, 50, 51, 96 |
| Model | Equation | Chem | Parameter Values | ||||
|---|---|---|---|---|---|---|---|
| a | b | c | R2 | SSE | |||
| Gompertz | Y = a × exp[−exp(b − c × t)] | Baicalin | 235.492 | 3.303 | 0.076 | 0.936 | 19,527.371 |
| Baicalin * | 5.485 | 1.172 | 0.058 | 0.977 | 1.172 | ||
| Wogonoside | 67.777 | 3.246 | 0.07 | 0.877 | 2928.026 | ||
| Wogonoside * | 4.205 | 0.896 | 0.043 | 0.923 | 2.609 | ||
| Logistic | Y = a/[1 + b × exp(−c × t)] | Baicalin | 233.686 | 366.795 | 0.12 | 0.933 | 20,164.7 |
| Baicalin * | 5.468 | 8.437 | 0.078 | 0.984 | 0.816 | ||
| Wogonoside | 64.517 | 171.301 | 0.098 | 0.878 | 2902.031 | ||
| Wogonoside * | 4.189 | 5.864 | 0.058 | 0.938 | 2.102 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Yu, Y.; Shi, R.; Xie, G.; Zhu, Y.; Wu, G.; Qin, M. Organ-Specific Metabolic Shifts of Flavonoids in Scutellaria baicalensis at Different Growth and Development Stages. Molecules 2018, 23, 428. https://doi.org/10.3390/molecules23020428
Xu J, Yu Y, Shi R, Xie G, Zhu Y, Wu G, Qin M. Organ-Specific Metabolic Shifts of Flavonoids in Scutellaria baicalensis at Different Growth and Development Stages. Molecules. 2018; 23(2):428. https://doi.org/10.3390/molecules23020428
Chicago/Turabian StyleXu, Jingyuan, Yilan Yu, Ruoyun Shi, Guoyong Xie, Yan Zhu, Gang Wu, and Minjian Qin. 2018. "Organ-Specific Metabolic Shifts of Flavonoids in Scutellaria baicalensis at Different Growth and Development Stages" Molecules 23, no. 2: 428. https://doi.org/10.3390/molecules23020428
APA StyleXu, J., Yu, Y., Shi, R., Xie, G., Zhu, Y., Wu, G., & Qin, M. (2018). Organ-Specific Metabolic Shifts of Flavonoids in Scutellaria baicalensis at Different Growth and Development Stages. Molecules, 23(2), 428. https://doi.org/10.3390/molecules23020428