Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
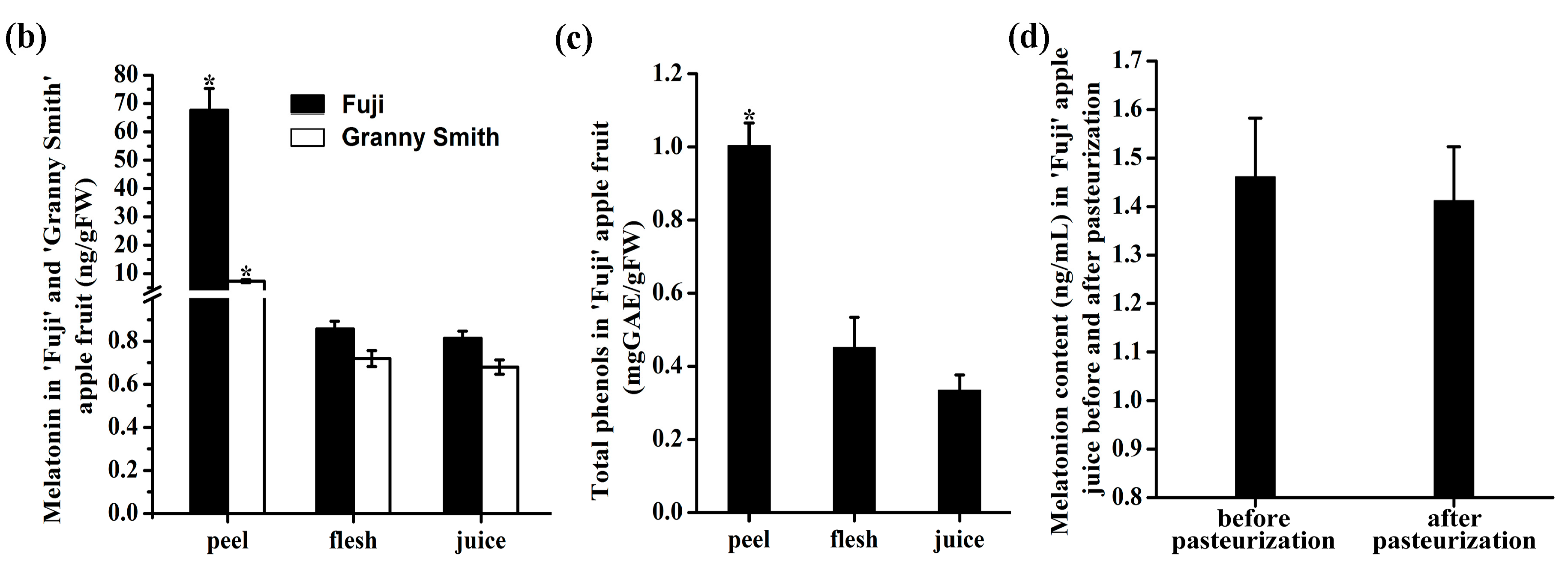
2.1. The Content of Melatonin and Total Phenols in ‘Fuji’ and ’Granny Smith’ Apple Fruit
2.2. The Effect of Pasteurization on the Melatonin Content in Apple Juice
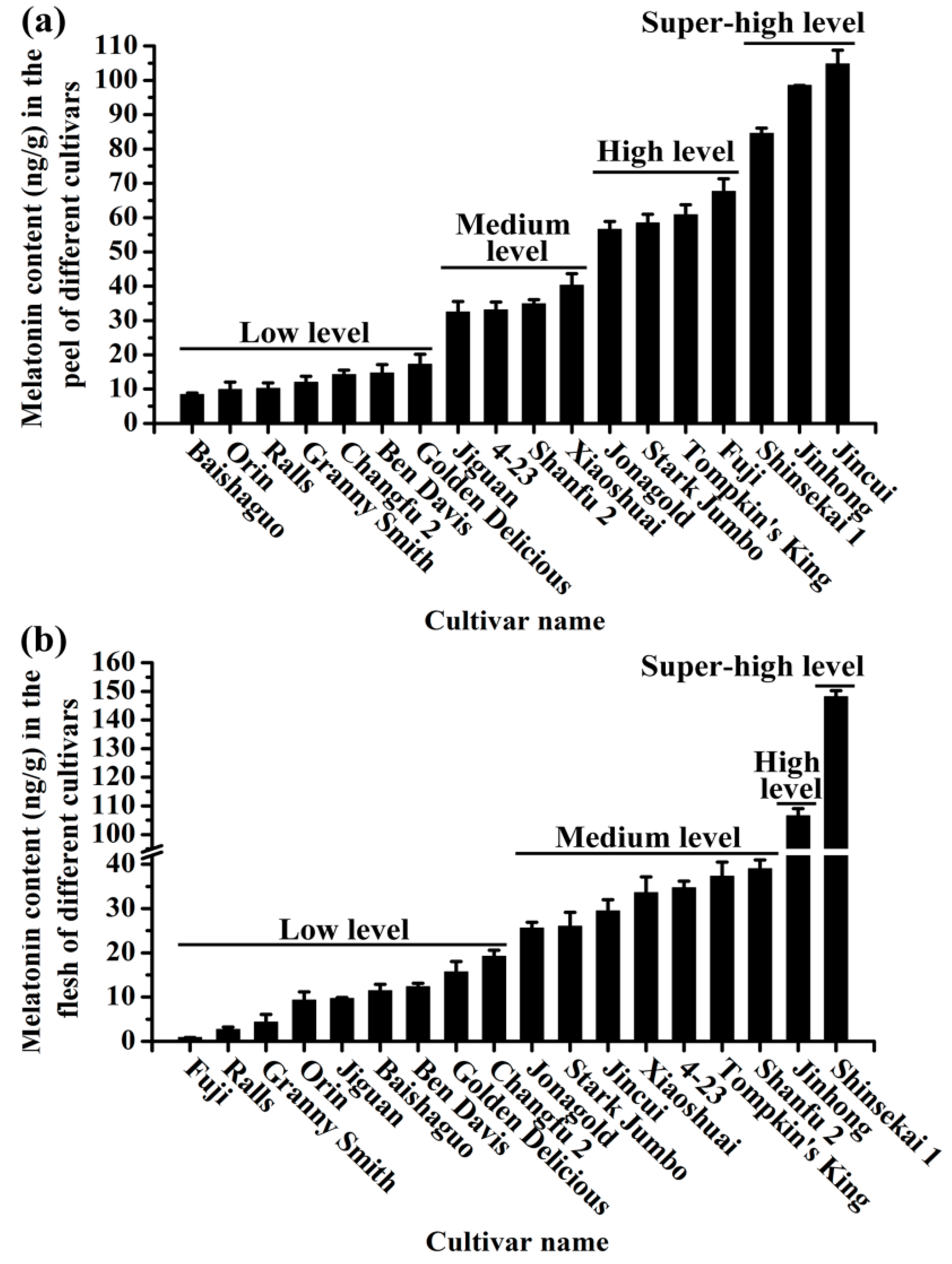
2.3. The Melatonin Content in Different Apple Cultivars
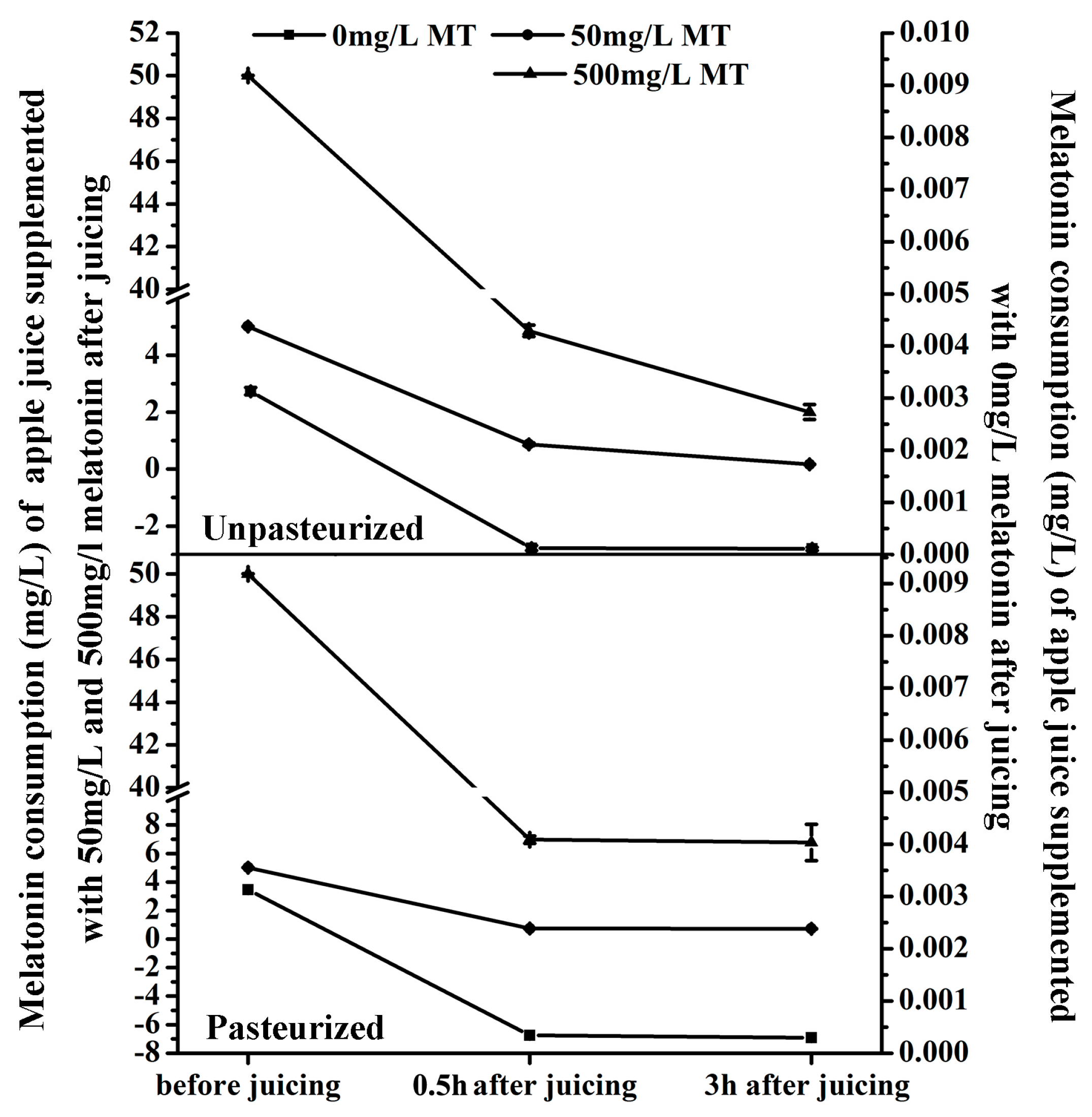
2.4. The Melatonin Content Reduced during Juicing Procedure and Storage Time
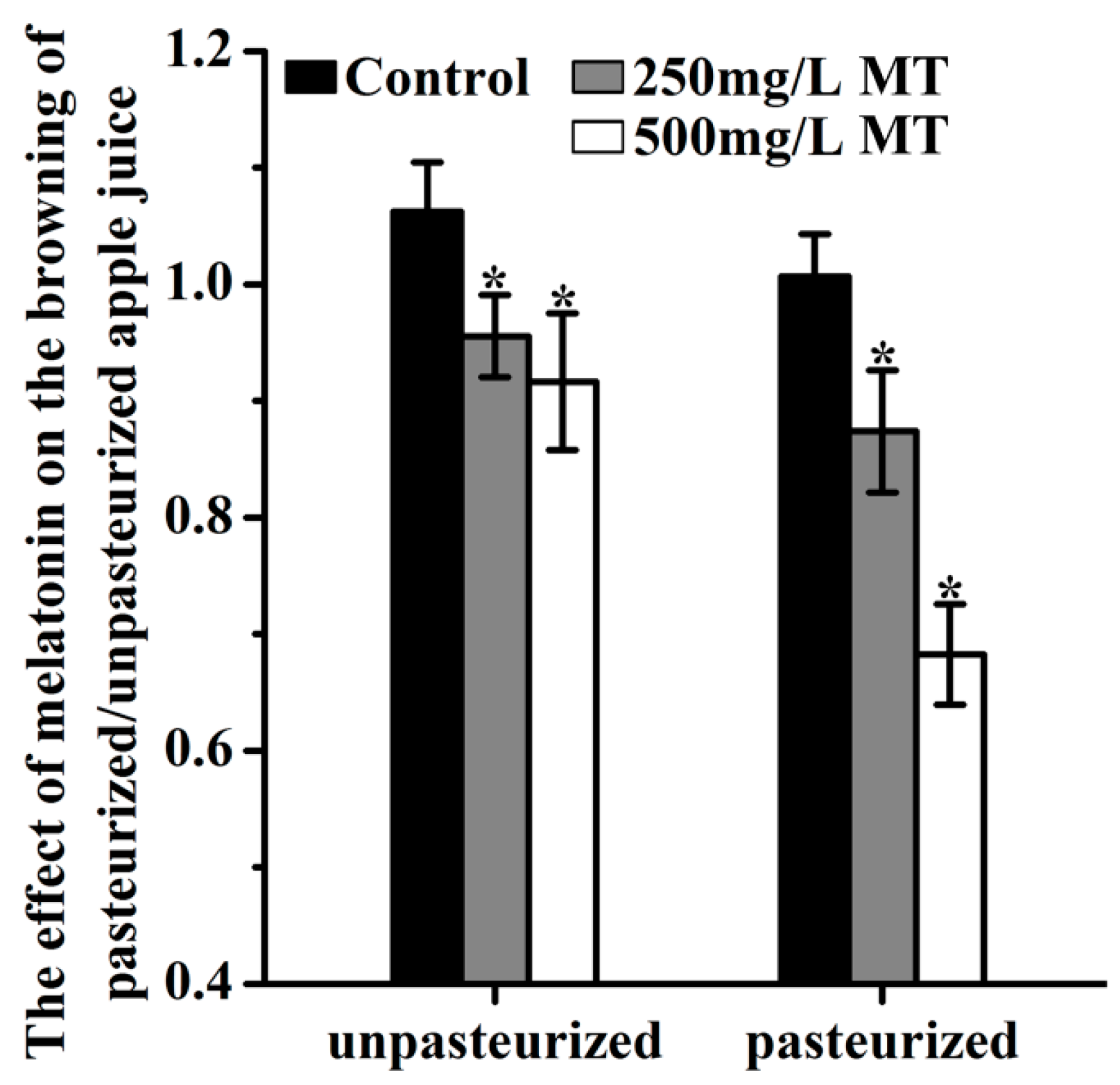
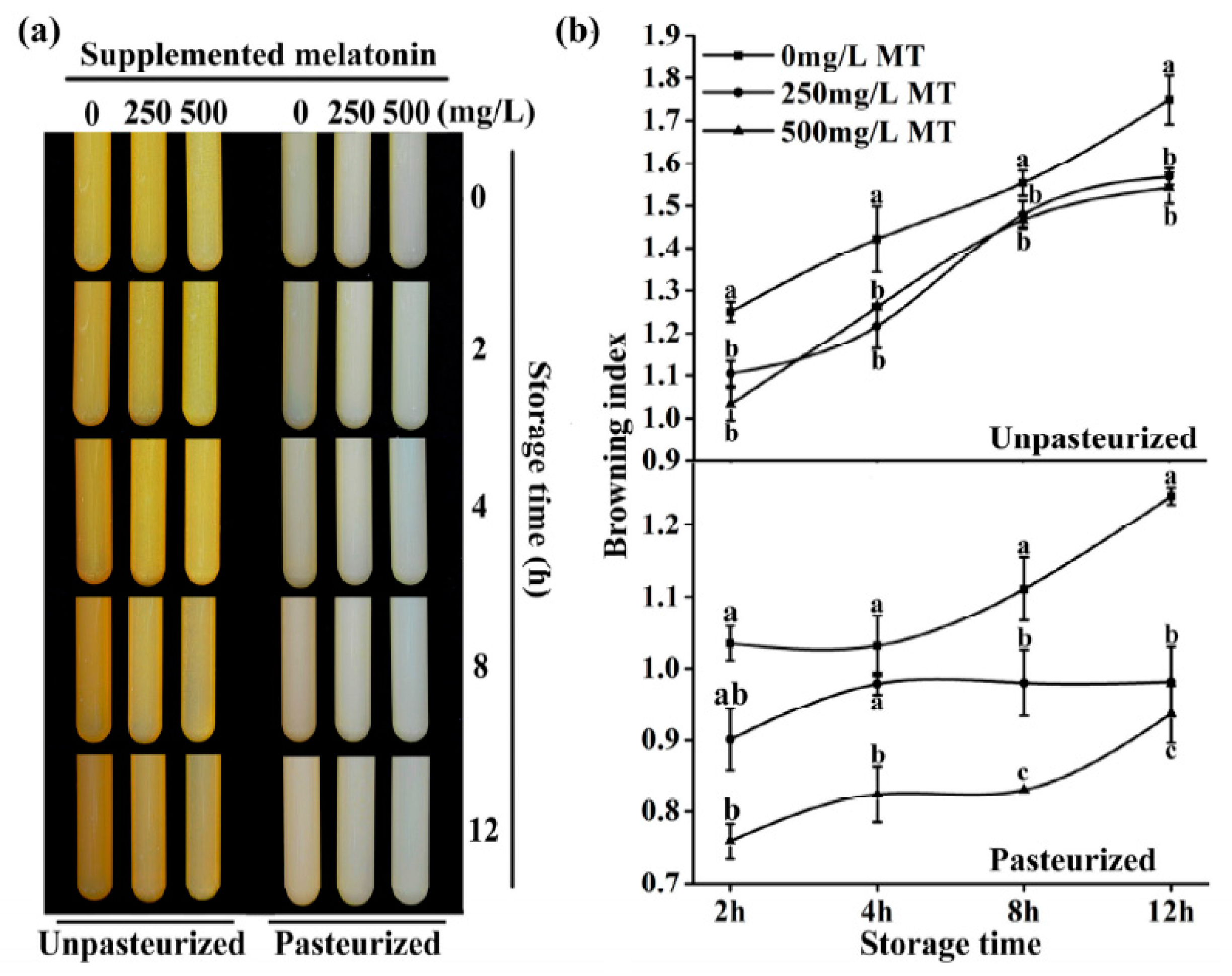
2.5. The Effect of Exogenous Melatonin on the Browning for ‘Fuji’ Apple Juicing
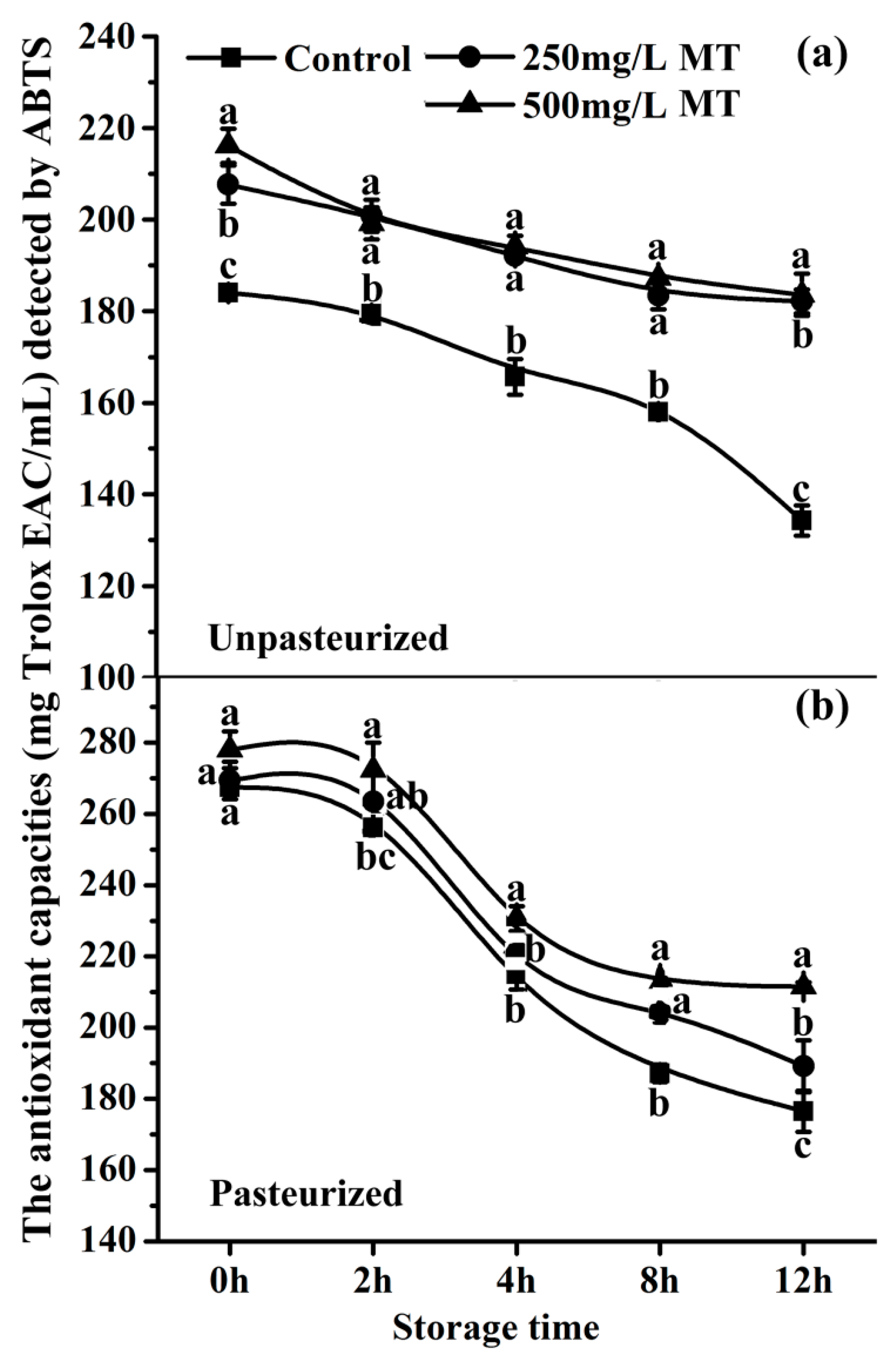
2.7. The Antioxidant Capacity of Exogenous Melatonin in Unpasteurized and Pasteurized ‘Fuji’ Apple Juice during Storage
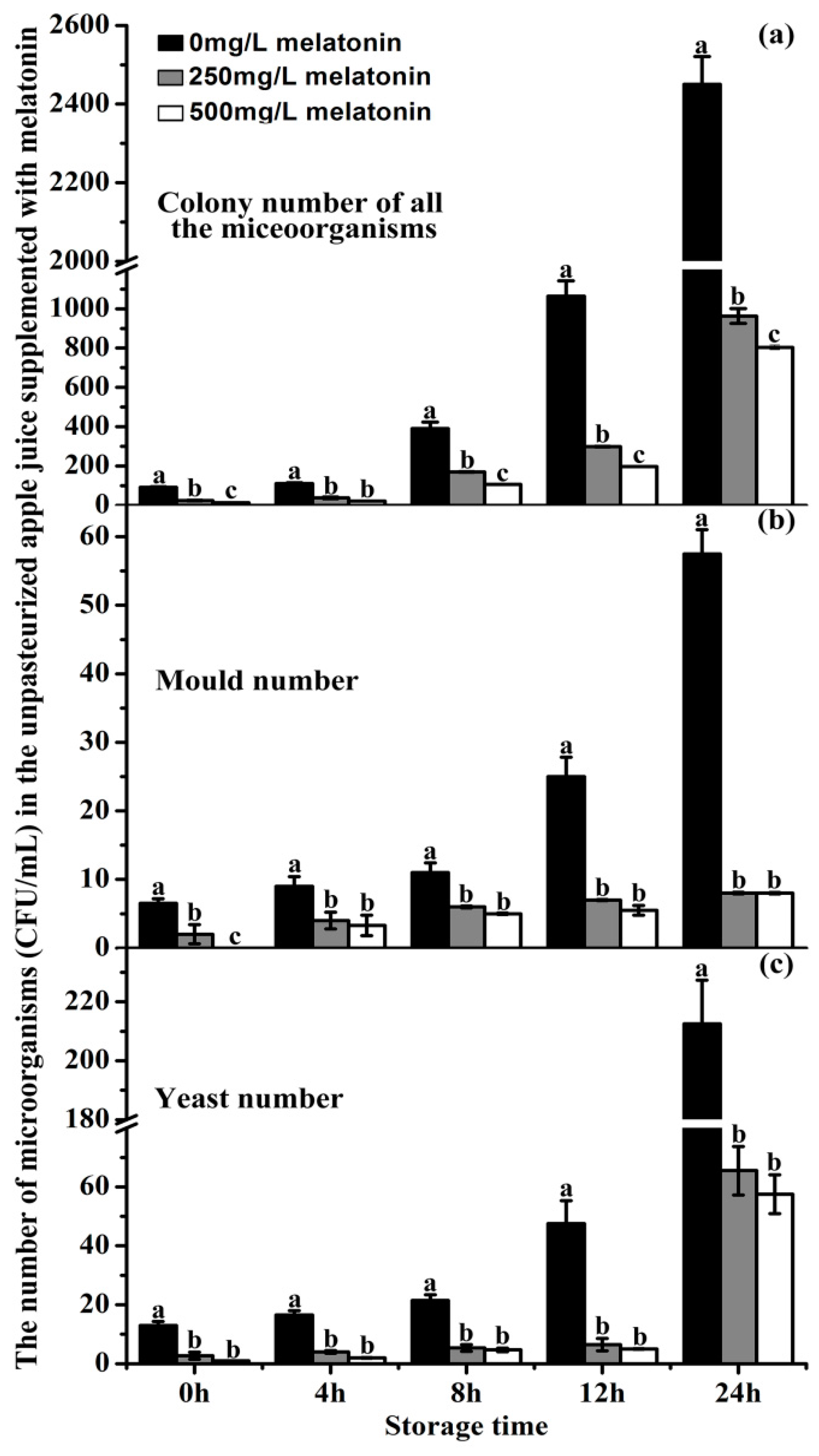
2.8. The Effects of Exogenous Melatonin on the Microorganism Growth in ‘Fuji’ Apple Juice during Storage
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Growth Conditions
4.3. Preparation of Apple Peel, Flesh and Juice
4.4. The Melatonin Supplementation and Juice Pasteurization
4.5. Melatonin and Total Phenols Detection of Apple Fruit
4.5.1. Melatonin Detection
4.5.2. Total Phenols Detection
4.6. Browning Observation and the Browning Index Detection of Apple Juice
4.6.1. Browning Observation
4.6.2. Browning Index Detection
4.7. Detection of Antioxidant Capacity of Apple Juice by the ABTS Assay
4.8. The inhibition of Melatonin on Microorganisms
4.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Available online: http://faostat.fao.org/default.aspx?alias=faostatclassic (accessed on 23 February 2018).
- US Apple Statistics, US Apple Association. Available online: http://www.usapple.org/industry/applestats/index.cfm (accessed on 23 February 2018).
- Ibanez, A.M.; Dandekar, A.M. Apple. In Transgenic Crops V; Pua, E.C., Davey, M.R., Eds.; Springer: Berlin and Heidelberg, Germany, 2007; p. 60. [Google Scholar]
- Hecke, K.; Herbinger, K.; Veberič, R.; Trobec, M.; Toplak, H.; Štampar, F.; Keppel, H.; Grill, D. Sugar-, acid- and phenol contents in apple cultivars from organic and integrated fruit cultivation. Eur. J. Clin. Nutr. 2006, 60, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Biedrzycka, E.; Amarowicz, R. Diet and health: apple polyphenols as antioxidants. Food Rev. Int. 2008, 24, 235–251. [Google Scholar] [CrossRef]
- Petkovsek, M.M.; Stampar, F.; Veberic, F. Changes in the inner quality parameters of apple fruit from technological to edible maturity. Acta Agric. Slov. 2009, 93, 17–29. [Google Scholar]
- Radunić, M.; Klepo, T.; Strikić, F.; Lukić, D.; Maretić, M. Karakteristike sorata jabuka (Malus x domestica Borkh.) uzgajanih na području Žrnovnice. Pomologia Croat. 2011, 17, 11–17. [Google Scholar]
- Balík, J.; Rop, O.; Mlček, J.; Híc, P.; Horák, M.; Řezníček, V. Assessment of nutritional parameters of native apple cultivars as new gene sources. Acta Univ. Agric. Silvic. Mendelianae Brun. 2012, 60, 27–38. [Google Scholar] [CrossRef]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Marinoni, D.T.; Cerutti, A.K.; Canterino, S.; Bounous, G. Application of sensory, nutraceutical and genetic techniques to create a quality profile of ancient apple cultivars. J. Food Qual. 2012, 35, 169–181. [Google Scholar] [CrossRef]
- Candrawinata, V.I.; Golding, J.B.; Roach, P.D.; Stathopoulos, C.E. From apple to juice—the fate of polyphenolic compounds. Food Rev. Int. 2013, 29, 276–293. [Google Scholar] [CrossRef]
- Biegańska-Marecik, R.; Radziejewska-Kubzdela, E.; Marecik, R. Characterization of phenolics, glucosinolates and antioxidant activity of beverages based on apple juice with addition of frozen and freeze-dried curly kale leaves (Brassica oleracea L. var. acephala L.). Food Chem. 2017, 230, 271–280. [Google Scholar]
- Hyson, D.A. A comprehensive review of apples and apple components and their relationship to human health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; HARA, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Afzalia, D.; Azadmehra, F.; Torkzadeha, M. Vortex-assisted dispersive liquid–liquid microextraction trace amounts of melatonin prior to HPLC determination in fruit juice samples. Sep. Sci. Technol. 2016, 51, 1509–1514. [Google Scholar] [CrossRef]
- Howatson, G.; Bell, P.G.; Tallent, J.; Middleton, B.; McHugh, M.P.; Ellis, J. Effect of tart cherry juice (Prunus cerasus) on melatonin levels and enhanced sleep quality. Eur. J. Nutr. 2012, 51, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pachón, M.S.; Medina, S.; Herrero-Martín, G.; Cerrillo, I.; Berna, G.; Escudero-López, B.; Ferreres, F.; Martín, F.; García-Parrilla, M.C.; Gil-Izquierdo, A. Alcoholic fermentation induces melatonin synthesis in orange juice. J. Pineal Res. 2014, 56, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Herrera, T.; Benítez, V.; Arribas, S.M.; López de Pablo, A.L.; Esteban, R.M.; Martín-Cabrejas, M.A. Estimation of scavenging capacity of melatonin and other antioxidants: Contribution and evaluation in germinated seeds. Food Chem. 2015, 170, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.F.; Shi, T.C.; Song, S.; Zhang, Z.W.; Fang, Y.L. Melatonin in grapes and grape-related foodstuffs: A review. Food Chem. 2017, 231, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Wang, L.; Tan, D.X.; Zhao, Y.; Zheng, X.D.; Chen, H.; Li, Q.T.; Zuo, B.X.; Kong, J. Identification of genes for melatonin synthetic enzymes in ‘Red Fuji’ apple (Malus domestica Borkh.cv. Red) and their expression and melatonin production during fruit development. J. Pineal Res. 2013, 55, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S.; Daglia, M.; Sureda, A.; Selamoglu, Z.; Gulhan, M.F.; Nabavi, S.M. Melatonin and Respiratory Diseases: A Review. Curr. Top. Med. Chem. 2017, 17, 467–488. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, Z.; Zheng, H.Y.; Ho, J.; Chan, M.T.V.; Wu, W.K.K. Protective roles of melatonin in central nervous system diseases by regulation of neural stem cells. Cell Prolif. 2017, 50, e12323. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; RosalesCorral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2012, 63, 577–597. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Zubero, E.; López-Pingarrón, L.; Alatorre-Jiménez, M.A.; Ochoa-Moneo, P.; Buisac-Ramón, C.; Rivas-Jiménez, M.; Castán-Ruiz, S.; Antoñanzas-Lombarte, Á.; Tan, D.X.; García, J.J.; et al. Melatonin’s role as a co-adjuvant treatment in colonic diseases: A review. Life Sci. 2017, 170, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Asghari, M.H.; Abdollahi, M.; De Oliveira, M.R.; Nabavi, S.M. A review of the protective role of melatonin during phosphineinduced cardiotoxicity: Focus on mitochondrial dysfunction, oxidative stress and apoptosis. J. Pharm. Pharmacol. 2017, 69, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sci. 2017, 173, 94–106. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, G.; Marseglia, L.; Manti, S.; Colavita, L.; Cuppari, C.; Impellizzeri, P.; Arena, S.; Arrigo, T.; Salpietro, C.; Gitto, E. Atopy and autoimmune thyroid diseases: Melatonin can be useful? Ital. J. Pediatr. 2016, 42, 95. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Hsieh, M.J.; Yang, W.E.; Chung, W.H.; Reiter, R.J.; Yang, S.F. Cancer metastasis: Mechanisms of inhibition by melatonin. J. Pineal Res. 2017, 62, e12370. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N. Developmental Programming of Adult Disease: Reprogramming by Melatonin? Int. J. Mol. Sci. 2017, 18, 426. [Google Scholar] [CrossRef] [PubMed]
- Ramos, E.; Patiño, P.; Reiter, R.J.; Gil-Martín, E.; Marco-Contelles, J.; Parada, E.; De Los Rios, E.; Romero, A.; Egea, J. Ischemic brain injury: New insights on the protective role of melatonin. Free Radic. Biol. Med. 2017, 104, 32–53. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Ojike, N.I.; Akinseye, O.A.; Kendzerska, T.; Buttoo, K.; Dhandapany, P.S.; Brown, G.M.; Cardinali, D.P. Melatonin and Human Cardiovascular Disease. Pharmacol. Ther. 2017, 22, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and the pathologies of weakened or dysregulated circadian oscillators. J. Pineal Res. 2017, 62, e12377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, S.; Li, Y.; Liu, Y. Melatonin as a promising agent of regulating stem cell biology and its application in disease therapy. Pharmacol. Res. 2017, 117, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Watson, N.; Diamandis, T.; Gonzales-Portillo, C.; Reyes, S.; Borlongan, C.Y. Melatonin as an Antioxidant for Stroke Neuroprotection. Cell Transplant. 2016, 25, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Yin, T.C.; Sung, P.H.; Chiang, J.Y.; Sun, C.K.; Yip, H.K. Melatonin enhances survival and preserves functional integrity of stem cells: A review. J. Pineal Res. 2017, 62, e12372. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Zubero, E.; García-Gil, F.A.; López-Pingarrón, L.; Alatorre-Jiménez, M.A.; Ramírez, J.M.; Tan, D.X.; García, J.J.; Reiter, R.J. Melatonin role preventing steatohepatitis and improving liver transplantation results. Cell Mol. Life Sci. 2016, 73, 2911–2927. [Google Scholar] [CrossRef] [PubMed]
- Nooshinfar, E.; Safaroghli-Azar, A.; Bashash, D.; Akbari, M.E. Melatonin, an inhibitory agent in breast cancer. Breast Cancer 2017, 24, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.W.; Zhang, X.; Huang, W.J. Pain control by melatonin: Physiological and pharmacological effects. Exp. Ther. Med. 2016, 12, 1963–1968. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Ma, Z.Q.; Di, S.; Jiang, S.; Li, Y.; Fan, C.X.; Yang, Y.; Wang, D.J. Snapshot: Implications for melatonin in endoplasmic reticulum homeostasis. Br. J. Pharmacol. 2016, 173, 3431–3442. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Cordaro, M.; Esposito, E.; Cuzzocrea, S. The antioxidative property of melatonin against brain ischemia. Expert Rev. Neurother. 2016, 16, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.A.; Garde, A.H.; Kristiansen, J.; Nabe-Nielsen, K.; Hansen, A.M. The effect of the number of consecutive night shifts on diurnal rhythms in cortisol, melatonin and heart rate variability (HRV): A systematic review of field studies. Int. Arch. Occup. Environ. Health 2016, 89, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Campa, C.; Menéndez-Menéndez, J.; Alonso-González, C.; González, A.; Álvarez-García, V.; Cos, S. What is known about melatonin, chemotherapy and altered gene expression in breast cancer. Oncol. Lett. 2017, 13, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchster, L.C.; Qi, W.B.; Karbownik, M.; Calvo, J.R. Significance of melatonin in antioxidative defense system: Reactions and products. Biol. Signals Recept. 2000, 9, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchster, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: a never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchster, L.C.; Di Mascio, P.; Martinez, G.R.; Prado, F.M.; Reiter, R.J. Novel rhythms of N1-acetyl-N2-formyl-5-methoxykynuramine and its precursor melatonin in water hyacinth:importance for phytoremediation. Faseb. J. 2007, 21, 1724–1729. [Google Scholar] [CrossRef] [PubMed]
- Guenther, A.L.; Schmidt, S.I.; Laatsch, H.; Fotso, S.; Ness, H.; Ressmeyer, A.R.; Poeggeler, B.; Hardeland, R. Reactions of the melatonin metabolite AMK (N1-acetyl-5-methoxykynuramine) with reactive nitrogen species: formation of novel compounds, 3-acetamidomethyl-6-methoxycinnolinone and 3-nitro-AMK. J. Pineal Res. 2005, 39, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: a physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Blask, D.E.; Dauchy, R.T.; Sauer, L.A.; Krause, J.A. Melatonin uptake and growth prevention in rat hepatoma 7288CTC in response to dietary melatonin: Melatonin receptor-mediated inhibition of tumor linoleic acid metabolism to the growth signaling molecule 13-hydroxyoctadecadienoic acid and the potential role of phytomelatonin. Carcinogenesis 2004, 25, 951–960. [Google Scholar] [PubMed]
- Billaud, C.; Brun-Mérimée, S.; Louarme, L.; Nicolas, J. Effect of glutathione and Maillard reaction products prepared from glucose or fructose with glutathione on polyphenoloxidase from apple—I: Enzymatic browning and enzyme activity inhibition. Food Chem. 2004, 84, 223–233. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Bioche. 1976, 72, 248–257. [Google Scholar] [CrossRef]
- Burdurlu, H.S.; Karadeniz, F. Effect of storage on nonenzymatic browning of apple juice concentrates. Food Chem. 2003, 80, 91–97. [Google Scholar] [CrossRef]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Hellens, R.P. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, C.Y.; Park, I. Prevention of enzymatic browning of pear by onion extract. Food Chem. 2005, 89, 181–184. [Google Scholar] [CrossRef]
- Sudha, G.; Priya, M.S.; Shree, R.I.; Vadivukkarasi, S. In vitro free radical scavenging activity of raw pepino fruit (Solanum muricatum aiton). Int. J. Curr. Pharm. Res. 2011, 3, 137–140. [Google Scholar]
- Lu, S.M.; Luo, Y.G.; Turner, E.; Feng, H. Efficacy of sodium chlorite as an inhibitor of enzymatic browning in apple slices. Food Chem. 2007, 104, 824–829. [Google Scholar] [CrossRef]
- López-Nicolás, J.M.; Núñez-Delicado, E.; Sánchez-Ferrer, Á.; García-Carmona, F. Kinetic model of apple juice enzymatic browning in the presence of cyclodextrins: The use of maltosyl-β-cyclodextrin as secondary antioxidant. Food Chem. 2007, 101, 1164–1171. [Google Scholar] [CrossRef]
- Luo, Y.; Lu, S.; Zhou, B.; Feng, H. Dual effectiveness of sodium chlorite for enzymatic browning inhibition and microbial inactivation on fresh-cut apples. LWT-Food Sci. Technol. 2011, 44, 1621–1625. [Google Scholar] [CrossRef]
- Zhao, D.Y.; Yuan, J.C.; Xu, K.; Cheng, C.G.; Yan, S. Analysis of apple production and marketing at home and abroad in the past 10 years. Chinese Tree 2016, 3, 87–93. [Google Scholar]
- Cerezo, A.B.; Leal, A.; Alvarez-Fernandez, M.A.; Hornedo-Ortega, R.; Troncoso, A.M.; Garcia-Parrilla, M.C. Quality control and determination of melatonin in food supplements. J. Food Compos. Anal. 2016, 45, 80–86. [Google Scholar] [CrossRef]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.; Kim, Y.J.; Kim, D.O.; Lee, H.J.; Lee, C.Y. Major phenolics in apple and their contribution to the total antioxidant capacity. J. Agric. Food Chem. 2003, 51, 6516–6520. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant activity of apple peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Petkovšek, M.M.; Štampar, F.; Veberič, R. Parameters of inner quality of the scab resistant and susceptible apple in organic and integrated production. Sci. Hortic. 2007, 114, 37–44. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Growth conditions determine different melatonin levels in Lupinus albus L. J. Pineal Res. 2013, 55, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.R.; Fan, J.B.; Xie, Y.; Amombo, E.; Liu, A.; Gitau, M.M.; Khaldun, A.B.M.; Chen, L.; Fu, J.M. Comparative photosynthetic and metabolic analyses reveal mechanism of improved cold stress tolerance in bermudagrass by exogenous melatonin. Plant Physio. Biochem. 2016, 100, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.H.; Huang, B.; Ding, C.B.; Zhang, Z.W.; Chen, Y.E.; Hu, C.; Zhou, L.J.; Huang, Y.; Liao, J.Q.; Yuan, S.; et al. Effects of melatonin on anti-oxidative systems and photosystem II in cold-stressed rice seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Yao, J.F.; Wang, X.N.; You, C.X.; Wang, X.F.; Hao, Y.J. MdHY5 positively regulates cold tolerance via CBF-dependent and CBF independent pathways in apple. J. Plant Physiol. 2017, 218, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, S.V.; Shukla, R.M.; Sherif, M.S.; Murch, J.S.; Saxena, K.P. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Turk, H.; Erdal, S.; Genisel, M.; Atici, O.; Demir, Y.; Yanmis, D. The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul. 2014, 74, 139–152. [Google Scholar] [CrossRef]
- Zheng, X.D.; Tan, D.X.; Allan, C.A.; Zuo, B.X.; Zhao, Y.; Reiter, J.R.; Wang, L.; Wang, Z.; Guo, Y.; Zhou, J.Z.; et al. Chloroplastic biosynthesis of melatonin and its involvement in protection of plants from salt stress. Sci. Rep. 2017, 7, 41236. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.D.; Zhou, J.Z.; Tan, D.X.; Wang, N.; Wang, L.; Shan, D.Q.; Kong, J. Melatonin improves waterlogging tolerance of Malus baccata (Linn.) Borkh. seedlings by maintaining aerobic respiration, photosynthesis and ROS migration. Front. Plant Sci. 2017, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Tan, D.X.; Reiter, R.J. Clinical uses of melatonin: Evaluation of human trials. Curr Med. Chem. 2010, 17, 2070–2095. [Google Scholar] [CrossRef] [PubMed]
- SFDA. State Food and Drug Administration. Available online: http://www.sfda.gov.cn/WS01/CL0055/10396_7.html China (accessed on 20 May 2005).
- Martínez, M.V.; Whitaker, J.R. The biochemistry and control of enzymatic browning. Trends Food Sci. Tech. 1995, 6, 195–200. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Zhou, Z.; Cruz, M.H.; Fuentes-Broto, L.; Galano, A. Phytomelatonin: Assisting plants to survive and thrive. Molecules 2015, 20, 7396–7437. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Zheng, X.; Kong, J.; Manchester, L.C.; Hardeland, R.; Kim, S.J.; Xu, X.; Reiter, R.J. Fundamental issues related to the origin of melatonin and melatonin isomers during evolution: relation to their biological functions. Int. J. Mol. Sci. 2014, 15, 15858–15890. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, Y.; Tan, D.X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin induces nitric oxide and the potential mechanisms relate to innate immunity against bacterial pathogen infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Qian, Y.; Tan, D.X.; Reiter, R.J.; He, C. Melatonin induces the transcripts of CBF/DREBs and their involvement in abiotic and biotic stresses in Arabidopsis. J. Pineal Res. 2015, 59, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Wiid, L.; Helden, E.H.V.; Hon, D.; Lombard, C.; Helden, A.P.V. Potentiation of Isoniazid Activity against Mycobacterium tuberculosis by Melatonin. Antimicrob. Agents Chemother. 1999, 4, 975–977. [Google Scholar]
- Atroshia, F.; Rizzob, A.; Westermarck, A.; Ali-vehmas, T. Effects of tamoxifen, melatonin, coenzyme Q, and 10 L-carnitine supplementation on bacterial growth in the presence of mycotoxins. Pharmacol. Res. 1998, 38, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Tekbas, O.F.; Ogur, R.; Korkmaz, A.; Kilic, A.; Reiter, R.J. Melatonin as an antibiotic: new insights into the actions of this ubiquitous molecule. J. Pineal Res. 2008, 44, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Sumaira; Khan, T.; Abbasi, B.H.; Afridi, M.S.; Tanveer, F.; Ullah, I.; Bashira, S.; Hanod, C. Melatonin-enhanced biosynthesis of antimicrobial AgNPs by improving the phytochemical reducing potential of a callus culture of Ocimum basilicum L. var. thyrsiflora. Rsc. Adv. 2017, 7, 38699. [Google Scholar] [CrossRef]
- Romić, M.D.; Klarić, M.Š.; Lovrić, J.; Pepić, I.; Čižmek, B.C.; Grčić, J.F.; Hafner, A. Melatonin-loaded chitosan/Pluronic® F127 microspheres as in situ forming hydrogel: An innovative antimicrobial wound dressing. Eur. J. Pharm. Biopharm. 2016, 107, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Deng, C.; Ma, Z.Q.; Wang, D.J.; Fan, C.X.; Li, T.; Di, S.Y.; Gong, B.; Reiter, R.J.; Yang, Y. Utilizing melatonin to combat bacterial infections and septic injury. Br. J. Pharmacol. 2017, 174, 754–768. [Google Scholar] [CrossRef] [PubMed]
- GB 4789.2-2010. National food safety standard. Food microbiological examination: Aerobic plate count. Ministry of Health: Beijing, China, 2010.
- GB 4789.15-2010. National food safety standard. Food microbiological examination: Enumeriation of moulds and yeasts. Ministry of Health: Beijing, China, 2010.
- Beveridge, T. Haze and cloud in apple juice. Crit. Rev. Food Sci. Nutr. 1997, 37, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tan, D.X.; Lei, Q.; Chen, H.; Wang, L.; Li, Q.T.; Gao, Y.A.; Kong, J. Melatonin and its potential biological functions in the fruits of sweet cherry. J. Pineal Res. 2012, 55, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.; Reiter, R.J.; He, C.J.; Liu, G.S.; Lei, Q.; Zuo, B.X.; Zheng, X.D.; Li, Q.T.; Kong, J. Changes in melatonin levels in transgenic ‘Micro-Tom’ tomato overexpressing ovine AANAT and ovine HIOMT genes. J. Pineal Res. 2014, 56, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colourimetry of total phenolics with phosphomolybdic-phospholungistic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Valentina, U.; Jerneja, F.; Franci, Š. Sugars, organic acids, phenolic composition and antioxidant activity of sweet cherry (Prunus avium L.). Food Chem. 2008, 107, 185–192. [Google Scholar]
- Meydav, S.; Saguy, I.; Kopelman, I.J. Browning determination in citrus products. J. Agric. Food Chem. 1977, 25, 602–604. [Google Scholar] [CrossRef]
- Cruz-Cansino, N.D.; Ramirez-Moreno, E.; Leon-Rivera, J.E.; Delgado-Olivares, L.; Alanis-Garcia, E.; Ariza-Ortega, J.A.; Manriquez-Torres, J.D.; Jaramillo-Bustos, D.P. Shelf life, physicochemical, microbiological and antioxidant properties of purple cactus pear (Opuntia ficus indica) juice after thermoultrasound treatment. Ultrason. Sonochem. 2015, 27, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.H.; Kim, Y.M.; Nam, S.H.; Yim, S.H.; Eun, J.B. Enzymatic browning inhibition and antioxidant activity of pear juice from a new cultivar of asian pear (Pyrus pyrifolia Nakai cv. Sinhwa) with different concentrations of ascorbic acid. Food Sci. Biotechnol. 2016, 25, 153–158. [Google Scholar] [CrossRef]
- Lee, B.; Seo, J.D.; Rhee, J.K.; Kim, C.Y. Heated apple juice supplemented with onion has greatly improved nutritional quality and browning index. Food Chem. 2016, 201, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Valero, M.; Recrosio, N.; Saura, D.; Munoz, N.; Marti, N.; Lizama, V. Effects of ultrasonic treatments in orange juice processing. J. Food Eng. 2007, 80, 509–516. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Liu, X.; Chen, T.; Ji, Y.; Shi, K.; Wang, L.; Zheng, X.; Kong, J. Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice. Molecules 2018, 23, 521. https://doi.org/10.3390/molecules23030521
Zhang H, Liu X, Chen T, Ji Y, Shi K, Wang L, Zheng X, Kong J. Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice. Molecules. 2018; 23(3):521. https://doi.org/10.3390/molecules23030521
Chicago/Turabian StyleZhang, Haixia, Xuan Liu, Ting Chen, Yazhen Ji, Kun Shi, Lin Wang, Xiaodong Zheng, and Jin Kong. 2018. "Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice" Molecules 23, no. 3: 521. https://doi.org/10.3390/molecules23030521
APA StyleZhang, H., Liu, X., Chen, T., Ji, Y., Shi, K., Wang, L., Zheng, X., & Kong, J. (2018). Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice. Molecules, 23(3), 521. https://doi.org/10.3390/molecules23030521