Variable Levels of Tolerance to Water Stress (Drought) and Associated Biochemical Markers in Tunisian Barley Landraces


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
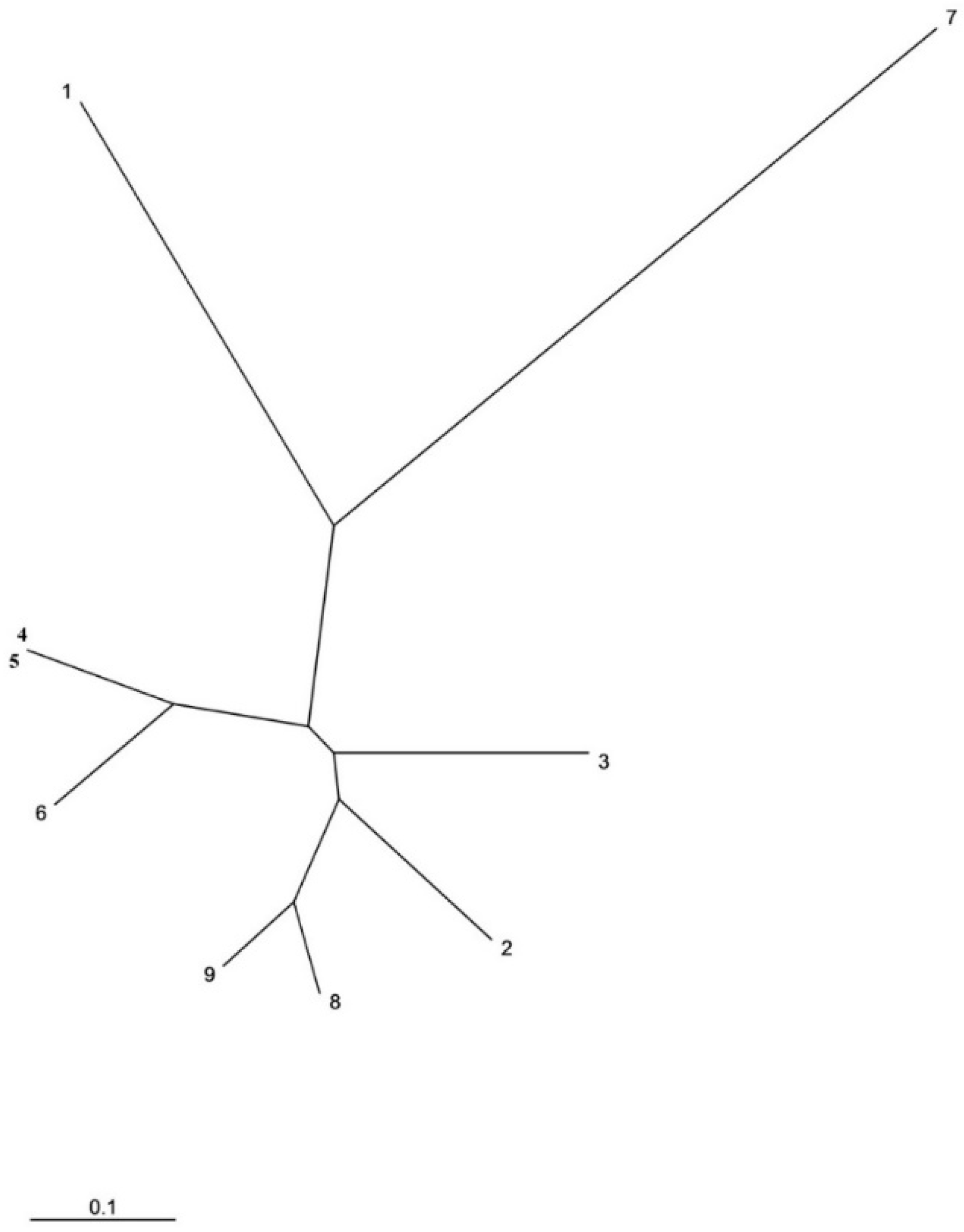
2.1. Molecular Characterization
2.2. Growth Parameters
2.3. Degradation of Photosynthetic Pigments
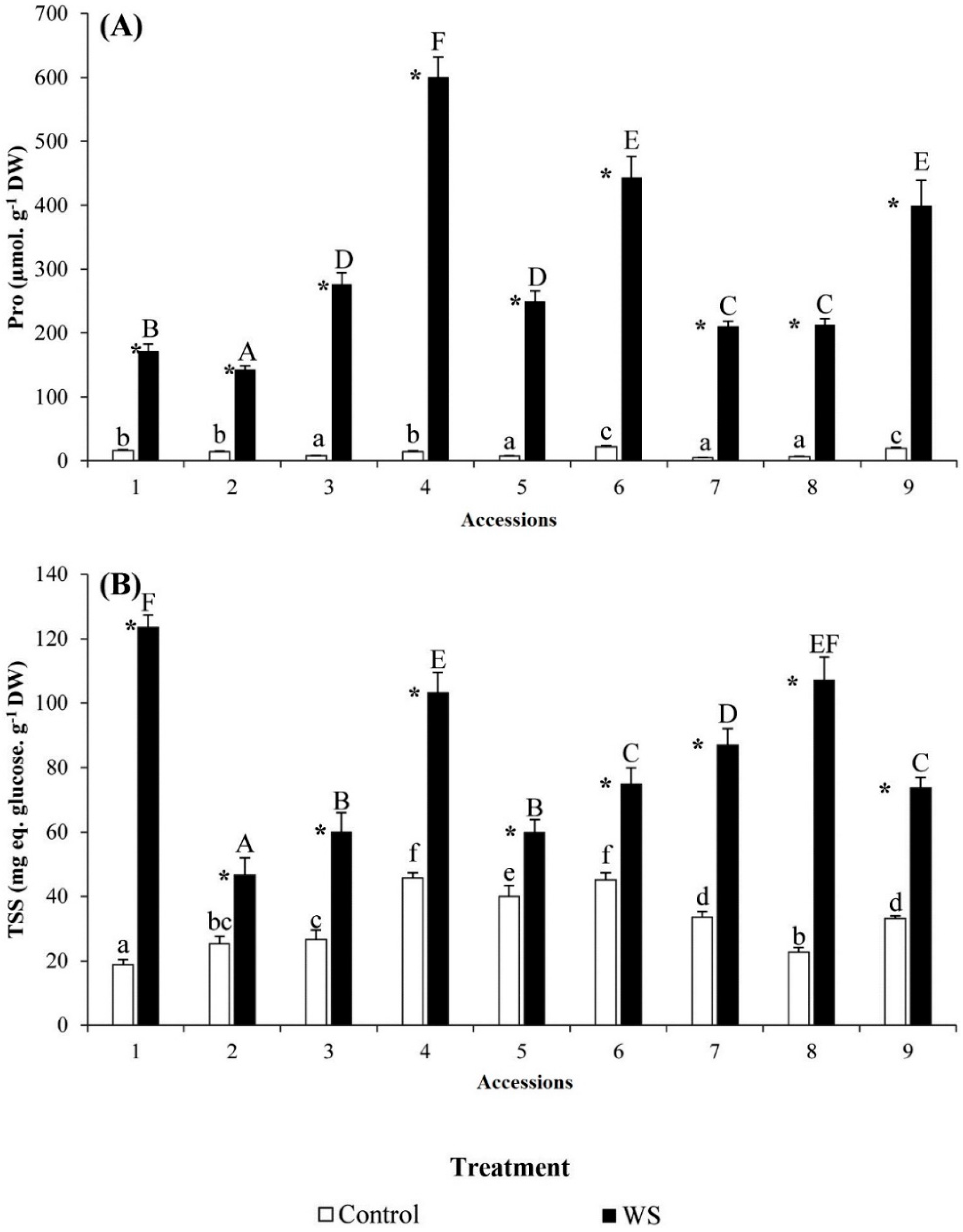
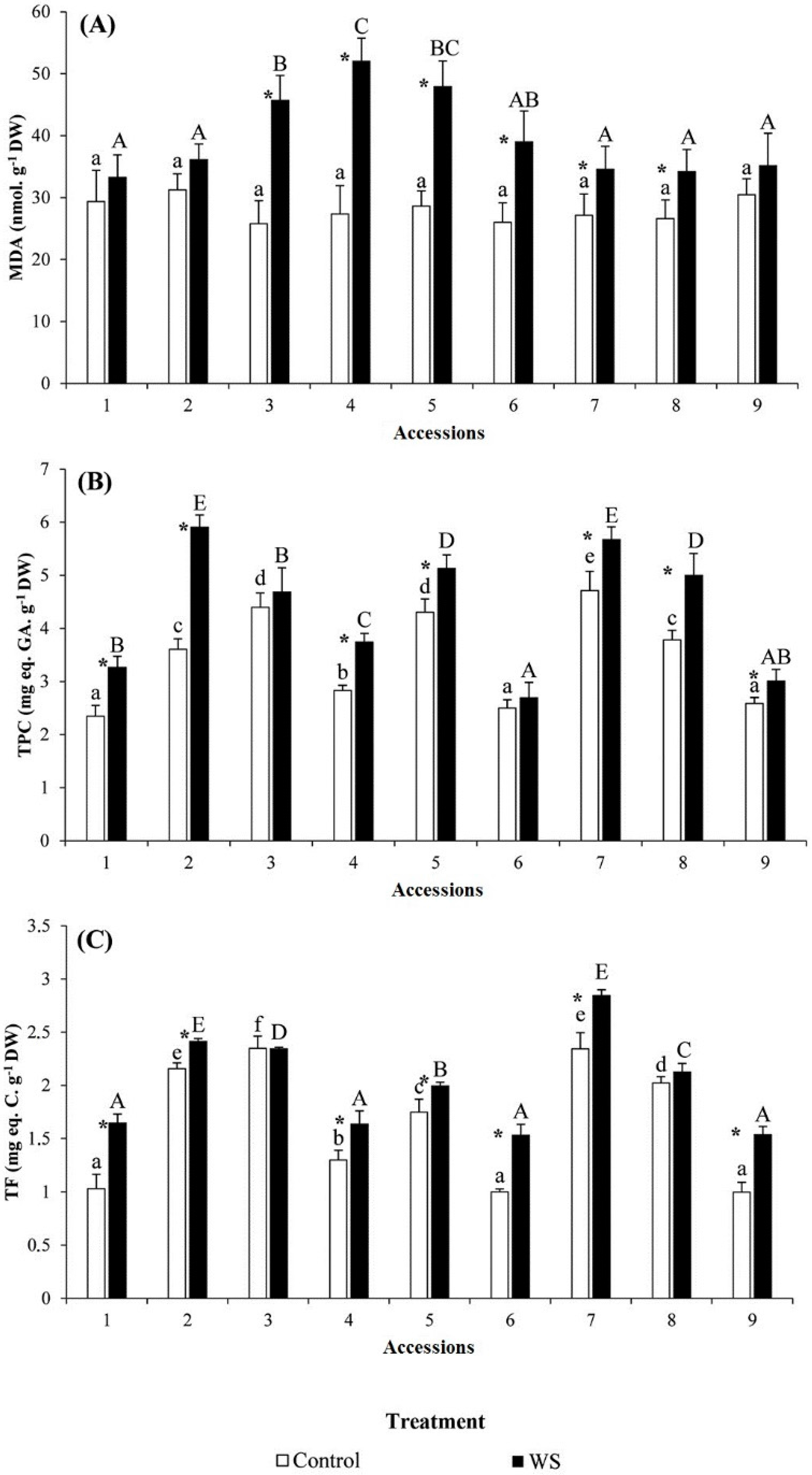
2.4. Osmolytes and Antioxidant Contents
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.1.1. Growth Conditions and Experimental Set-Up
4.1.2. DNA Extraction and Molecular Characterization
4.1.3. Measuring of Growth Parameters
4.1.4. Photosynthetic Pigments and Biochemical Parameters Measurement
4.2. Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Munns, R.; James, R.A.; Sirault, X.R.R.; Furbank, R.T.; Jones, H.J. New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J. Exp. Bot. 2010, 61, 3499–3507. [Google Scholar] [CrossRef] [PubMed]
- Gürel, F.; Öztürk, Z.N.; Uçarlı, C.; Rosellini, D. Barley genes as tools to confer abiotic stress tolerance in crops. Front. Plant Sci. 2016, 7, 1137. [Google Scholar] [CrossRef] [PubMed]
- Hellal, F.A.; El-Sayed, S.A.A.; El-Hady, M.A.; Khatab, I.A.; El-Shabrawi, H.M.; El-Menisy, A.M. Influence of salt stress on molecular and biochemical changes of barley at early seedling stage. Biosci. Res. 2017, 14, 417–426. [Google Scholar]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; de Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jin, Z.-Q.; Zhao, J.; Zhang, G.; Wu, F. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]
- Chmielewska, K.; Rodziewicz, P.; Swarcewicz, B.; Sawikowska, A.; Krajewski, P.; Marczak, Ł.; Ciesiołka, D.; Kuczynska, A.; Mikołajczak, K.; Ogrodowicz, P.; et al. Analysis of drought-induced proteomic and metabolomic changes in barley (Hordeum vulgare L.) leaves and roots unravels some aspects of biochemical mechanisms involved in drought tolerance. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cai, K.; Zhang, G.; Zeng, F. Metabolite profiling of barley grains subjected to water stress: To explain the genotypic difference in drought-induced impacts on malting quality. Front. Plant Sci. 2017, 8, 1547. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.B.; Cassin, A.; Keeble-Gagnère, G.; Doblin, M.S.; Bacic, A.; Roessner, U. De novo transcriptome assembly and analysis of differentially expressed genes of two barley genotypes reveal localized responses along salinized roots. Sci. Rep. 2016, 6, 31558. [Google Scholar] [CrossRef] [PubMed]
- Kintlová, M.; Blavet, N.; Cegan, R.; Hobza, R. Transcriptome of barley under three different heavy metal stress reaction. Genom. Data 2017, 13, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Zeng, X.; Qin, C.; Wang, Y.; Bai, L.; Xu, Q.; Yuan, H.; Tang, Y.; Nyima, T. Comparative transcriptome analysis revealed genes commonly responsive to varied nitrate stress in leaves of Tibetan hulless barley. Front. Plant Sci. 2016, 7, 1067. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Bai, L.; Wei, Z.; Yuan, H.; Wang, Y.; Xu, Q.; Tang, Y.; Nyim, T. Transcriptome analysis revealed the drought-responsive genes in Tibetan hulless barley. BMC Genom. 2016, 17, 386. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Qiu, L.; Xu, L.; Ye, L.; Chen, M.; Sun, D.; Chen, Z.; Zhang, H.; Jin, X.; Dai, F.; et al. Genetic variation of HvCBF genes and their association with salinity tolerance in Tibetan annual wild barley. PLoS ONE 2011, 6, e22938. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Li, R.; Bai, G.; Siddique, K.H.M.; Varshney, R.K.; Baum, M.; Yan, G.; Guo, P. Genetic variations of HvP5CS1 and their association with drought tolerance related traits in barley (Hordeum vulgare L.). Sci. Rep. 2017, 7, 7870. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fan, Y.; Mak, M.; Babla, M.; Holford, P.; Wang, F.; Chen, G.; Scott, G.; Wang, G.; Shabala, S.; et al. QTLs for stomatal and photosynthetic traits related to salinity tolerance in barley. BMC Genom. 2017, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Wójcik-Jagła, M.; Fiust, A.; Kościelniak, J.; Rapacz, M. Association mapping of drought tolerance-related traits in barley to complement a traditional biparental QTL mapping study. Theor. Appl. Genet. 2018, 131, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Newman, C.W.; Newman, R.K. A brief history of barley foods. Cereal Food World 2006, 51, 4–7. [Google Scholar] [CrossRef]
- Guasmi, F.; Touil, L.; Fères, K.; Elfelah, W.; Triki, T.; Ferchichi, A. Genetic diversity of Tunisian barley accessions based on microsatellite markers. Biotechnology 2008, 7, 781–782. [Google Scholar] [CrossRef]
- Ben Khaled, A.; Hayek, T.; Mansour, E.; Hannachi, H.; Lachiheb, B.; Ferchichi, A. Evaluating salt tolerance of 14 barley accessions from Southern Tunisia using multiple parameters. J. Agric. Sci. 2012, 4, 27–38. [Google Scholar] [CrossRef]
- El Faleh, M.; Mdimagh, S. Food barley in Tunisia. In Food Barley: Importance, Uses and Local Knowledge; Grando, S., Macpherson, H.G., Eds.; ICARDA: Aleppo, Syria, 2005; pp. 29–35. [Google Scholar]
- Thameur, A.; Lefi, E.; Ferchichi, A. Comparative response of barley cultivars and varieties to deficit irrigation in a Mediterranean environment. J. Arid Land 2009, 19, 153–156. [Google Scholar]
- Thameur, A.; Ferchichi, A.; López-Carbonell, M. Quantification of free and conjugated abscisic acid in five genotypes of barley (Hordeum vulgare L.) under water stress conditions. S. Afr. J. Bot. 2011, 77, 222–228. [Google Scholar] [CrossRef]
- Thameur, A.; Lachibeb, B.; Ferchichi, A. Drought effect on growth, gas exchange and yield, in two strains of local barley Ardhaoui, under water deficit conditions in southern Tunisia. J. Environ. Manag. 2012, 113, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Sbei, H.; Hammami, Z.; Trifa, Y.; Hamza, S.; Harrabi, M. Phenotypic diversity analysis for salinity tolerance of Tunisian barley populations (Hordeum vulgare L.). J. Arid Land 2012, 22, 57–60. [Google Scholar]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Towards an evolved concept of landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Allel, D.; Ben-Amar, A.; Lamine, M.; Abdelly, C. Relationships and genetic structure of North African barley (Hordeum vulgare L.) germplasm revealed by morphological and molecular markers: Biogeographical considerations. S. Afr. J. Bot. 2017, 112, 1–10. [Google Scholar] [CrossRef]
- Ben Khaled, A.; Hayek, T.; Manosur, E.; Ferchichi, A. Comparative effect of salinity on ion accumulation, grain yield and stability salt tolerance degree of barley (Hordeum vulgare L.) in different growth stage. J. Agric. Environ. Int. Dev. 2016, 110, 249–266. [Google Scholar]
- Stein, N.; Prasad, M.; Scholz, U.; Thiel, T.; Zhang, H.; Wolf, M.; Kota, R.; Varshney, R.K.; Perovic, D.; Grosse, I.; et al. A 1000-loci transcript map of the barley genome: New anchoring points for integrative grass genomics. Theor. Appl. Genet. 2007, 114, 823–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2014: Impacts, Adaptation, and Vulnerability. In Proceedings of the 5th Assessment Report, WGII; Cambridge University Press: Cambridge, UK; Available online: http://www.ipcc.ch/report/ar5/wg2/ (accessed on 7 March 2018).
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Kholová, J.; Prásil, I.T. Breeding for enhanced drought resistance in barley and wheat–drought-associated traits, genetic resources and their potential utilization in breeding programmes. Czech J. Genet. Plant Breed. 2014, 50, 247–261. [Google Scholar] [CrossRef]
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Amatriaín, M.; Cuesta-Marcos, A.; Endelman, J.B.; Comadran, J.; Bonman, J.M.; Bockelman, H.E.; Chao, S.; Russell, J.; Waugh, R.; Hayes, P.M. The USDA barley core collection: Genetic diversity, population structure, and potential for Genome-Wide Association studies. PLoS ONE 2014, 9, e94688. [Google Scholar] [CrossRef] [PubMed]
- Templer, S.E.; Ammon, A.; Pscheidt, D.; Ciobotea, O.; Schuy, C.; McCollum, C.; Sonnewald, U.; Hanemann, A.; Förster, J.; Ordon, F.; et al. Metabolite profiling of barley flag leaves under drought and combined heat and drought stress reveals metabolic QTLs for metabolites associated with antioxidant defense. J. Exp. Bot. 2017, 68, 1697–1713. [Google Scholar] [CrossRef] [PubMed]
- Abidi, I.; Mansouri, S.; Radhouane, L.; Ksouri, R.; El Felah, R.; Bouzid, S. Phenolic, flavonoid and tannin contents of Tunisian barley landraces. Int. J. Agric. Innov. Res. 2015, 3, 1317–1323. [Google Scholar]
- Hammami, Z.; Sbei, H.; Kadri, K.; Jmel, Z.; Sahli, A.; Belhaj Fraj, M.; Naser, H.; Teixeira da Silva, J.A.; Trifa, Y. Evaluation of performance of different barley genotypes irrigated with saline water in South Tunisian Saharan conditions. Environ. Exp. Bot. 2016, 14, 15–21. [Google Scholar] [CrossRef]
- Chéour, F.; Kaddachi, I.; Achouri, D.; Bannour, S.; Zorgu, L. Effects of water stress on relative water, chlorophylls and proline contents in barley (Hordeum vulgare L.) leaves. IOSR J. Agric. Vet. Sci. IOSR-JAVS 2014, 7, 13–16. [Google Scholar] [CrossRef]
- Bagues, M.; Soul, I.; Boussora, F.; Lachiheb, B.; Nagaz, K.K. Response of two barley accessions “Ardhaoui” to deficit irrigation using saline water in southern Tunisia. J. New Sci. 2017, 37, 2013–2023. [Google Scholar]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front. Plant Sci. 2015, 6, 1241. [Google Scholar] [CrossRef] [PubMed]
- Hura, T.; Grzesiak, S.; Hura, K.; Thiemt, E.; Tokarz, K.; Wedzony, M. Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter triticale: Accumulation of ferulic acid correlates with drought tolerance. Ann. Bot. Lond. 2007, 100, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Schiop, S.T.; Al Hassan, M.; Sestras, A.F.; Boscaiu, M.; Sestras, R.E.; Vicente, O. Identification of salt stress biomarkers in Romanian Carpathian populations of Picea abies (L.) Karst. PLoS ONE 2015, 10, e0135419. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Al Hassan, M.; Naranjo, M.A.; Agrawal, V.; Boscaiu, M.; Vicente, O. Effects of salinity and drought on growth, ionic relations, compatible solutes and activation of antioxidant systems in oleander (Nerium oleander L.). PLoS ONE 2017, 12, e0185017. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Martín, I.; Ayerbe, L. Response of barley genotypes to terminal soil moisture stress: Phenology, growth, and yield. Aust. J. Agric. Res. 2006, 58, 29–37. [Google Scholar] [CrossRef]
- Fathi, A.; Tari, D.B. Effect of drought stress and its mechanism in plants. Int. J. Life Sci. 2016, 10, 1–6. [Google Scholar] [CrossRef]
- Efeoglu, B.; Ekmekci, Y.; Cicek, N. Physiological responses of three maize cultivars to drought stress and recovery. S. Afr. J. Bot. 2009, 75, 34–42. [Google Scholar] [CrossRef]
- Guo, P.; Baum, M.; Grando, S.; Ceccarelli, S.; Bai, G.; Li, R.; von Korff, M.; Varshney, R.K.; Graner, A.; Valkoun, J. Differentially expressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage. J. Exp. Bot. 2009, 60, 3531–3544. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Guo, P.; Baum, M.; Grando, S.; Ceccarelli, S. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.N.; Aspinall, D.; Paleg, L.G. Proline accumulation and varietal adaptability to drought in barley: A potential metabolic measure of drought resistance. Nat. New Biol. 1972, 236, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Abdellaoui, R.; Tarhouni, M.; Chaabane, R.; Ben Naceur, M.; El Faleh, M.; Abdelli, C.; Ramla, D.; Nada, A.; Sakr, M.; Ben Hmid, J. Behaviour of Tunisian local barley accessions under progressive water deficit: Physiological and biochemical approach. J. Phytol. 2010, 2, 88–97. [Google Scholar]
- Velicevici, G.; Madosa, E.; Sumalan, R.; Ciulca, S.; Bitea, N.; Petolescu, C.; Petrescu, I.; Cretescu, I. Proline accumulation in some barley genotypes exposed to drought. J. Hortic. For. Biotechnol. 2011, 15, 48–54. [Google Scholar]
- Bandurska, H.; Niedziela, J.; Pietrowska-Borek, M.; Nuc, K.; Chadzinikolau, T.; Radzikowska, D. Regulation of proline biosynthesis and resistance to drought stress in two barley (Hordeum vulgare L.) genotypes of different origin. Plant Physiol. Biochem. 2017, 118, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Kameli, A.; Lösel, D.M. Contribution of carbohydrates and other solutes to osmotic adjustment in wheat leaves under water stress. J. Plant Physiol. 1995, 145, 363–366. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Cao, F.; Zhang, M.; Chen, X.; Zhang, G.; Wu, F. Difference in yield and physiological features in response to drought and salinity combined stress during anthesis in Tibetan wild and cultivated barleys. PLoS ONE 2013, 8, e77869. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The Water Culture Method for Growing Plants without Soil; California Agricultural Experimental Station Circular No. 347; University of California: Berkeley, CA, USA, 1950; pp. 1–32. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, L.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Reberd, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.C.; Palazzo de Mello, J.C. Application and analysis of the Folin Ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D.M. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity Statistics | SSR Marker | |||||
|---|---|---|---|---|---|---|
| GBM1176 | GBM1221 | GBM1256 | GBM1405 | GBM1459 | GBM1461 | |
| Linkage group | 5H | 4H | 6H | 3H | 2H | 1H |
| Number of alleles (Na) | 4 | 4 | 2 | 3 | 2 | 5 |
| Frequency of the major allele (f) | 0.667 | 0.500 | 0.778 | 0.667 | 0.500 | 0.556 |
| Effective number of alleles (Ne) | 2.05 | 2.84 | 1.53 | 1.98 | 2.00 | 2.79 |
| Polymorphic information content (PIC) | 0.473 | 0.593 | 0.286 | 0.438 | 0.375 | 0.610 |
| Accession | Treatment | Stem Length (cm) a | Number of Leaves a | Fresh Weight (g) a | Dry Weight (g) a |
|---|---|---|---|---|---|
| 1 | Control | 25.90 ± 1.29 | 8.00 ± 0.95 | 3.30 ± 0.41 | 0.22 ± 0.02 |
| WS | 20.56 ± 2.15 * | 5.26 ± 0.41 * | 0.71 ± 0.11 * | 0.17 ± 0.01 * | |
| 2 | Control | 24.15 ± 1.12 | 13.00 ± 1.15 | 4.46 ± 0.70 | 0.53 ± 0.05 |
| WS | 17.55 ± 0.45 * | 9.90 ± 0.69 * | 2.58 ± 0.22 * | 0.44 ± 0.03 * | |
| 3 | Control | 23.15 ± 1.84 | 22.25 ± 0.76 | 9.45 ± 0.51 | 0.91 ± 0.02 |
| WS | 9.71 ± 0.79 * | 9.54 ± 0.33 * | 1.22 ± 0.07 * | 0.37 ± 0.01 * | |
| 4 | Control | 25.90 ± 3.30 | 23.40 ± 0.92 | 8.69 ± 0.71 | 0.82 ± 0.05 |
| WS | 11.21 ± 0.34 * | 8.53 ± 0.55 * | 0.86 ± 0.04 * | 0.48 ± 0.02 * | |
| 5 | Control | 24.89 ± 0.78 | 18.00 ± 0.95 | 7.98 ± 0.29 | 0.85 ± 0.03 |
| WS | 9.74 ± 0.96 * | 9.86 ± 0.88 * | 1.26 ± 0.07 * | 0.40 ± 0.03 * | |
| 6 | Control | 22.08 ± 0.64 | 19.60 ± 0.81 | 6.87 ± 0.60 | 0.50 ± 0.04 |
| WS | 13.00 ± 0.19 * | 8.77 ± 0.59 * | 1.84 ± 0.22 * | 0.32 ± 0.02 * | |
| 7 | Control | 21.63 ± 1.01 | 17.80 ± 0.37 | 7.36 ± 0.67 | 0.63 ± 0.05 |
| WS | 12.11 ± 1.65 * | 8.50 ± 0.64 * | 1.57 ± 0.19 * | 0.33 ± 0.02 * | |
| 8 | Control | 22.02 ± 1.39 | 11.66 ± 0.68 | 5.71 ± 0.38 | 0.40 ± 0.02 |
| WS | 15.50 ± 1.03 * | 6.16 ± 0.35 * | 1.37 ± 0.24 * | 0.30 ± 0.03 * | |
| 9 | Control | 19.88 ± 0.56 | 30.20 ± 2.13 | 6.96 ± 0.47 | 0.61 ± 0.03 |
| WS | 15.20 ± 0.42 * | 9.31 ± 0.34 * | 1.37 ± 0.19 * | 0.44 ± 0.04 * |
| Code | Accession | Zone | Alt. (m) | T (°C) | R (mm) |
|---|---|---|---|---|---|
| 1 | Midon | Island | 50 | 21.25 | 311.25 |
| 2 | El May | Island | 35 | 20.37 | 336.13 |
| 3 | Guermassa | Mountain | 349 | 21.12 | 151.50 |
| 4 | Ksar Hdada | Mountain | 328 | 20.46 | 181.91 |
| 5 | Graguer | Mountain | 255 | 21.01 | 161.45 |
| 6 | Zarzis | Coast | 11 | 20.86 | 259.57 |
| 7 | Eljdaria | Coast | 14 | 20.47 | 204.05 |
| 8 | Ksar Bayada | Ridge | 185 | 21.08 | 239.14 |
| 9 | Smar Tataouine | Ridge | 85 | 21.09 | 158.20 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dbira, S.; Al Hassan, M.; Gramazio, P.; Ferchichi, A.; Vicente, O.; Prohens, J.; Boscaiu, M. Variable Levels of Tolerance to Water Stress (Drought) and Associated Biochemical Markers in Tunisian Barley Landraces. Molecules 2018, 23, 613. https://doi.org/10.3390/molecules23030613
Dbira S, Al Hassan M, Gramazio P, Ferchichi A, Vicente O, Prohens J, Boscaiu M. Variable Levels of Tolerance to Water Stress (Drought) and Associated Biochemical Markers in Tunisian Barley Landraces. Molecules. 2018; 23(3):613. https://doi.org/10.3390/molecules23030613
Chicago/Turabian StyleDbira, Sameh, Mohamad Al Hassan, Pietro Gramazio, Ali Ferchichi, Oscar Vicente, Jaime Prohens, and Monica Boscaiu. 2018. "Variable Levels of Tolerance to Water Stress (Drought) and Associated Biochemical Markers in Tunisian Barley Landraces" Molecules 23, no. 3: 613. https://doi.org/10.3390/molecules23030613
APA StyleDbira, S., Al Hassan, M., Gramazio, P., Ferchichi, A., Vicente, O., Prohens, J., & Boscaiu, M. (2018). Variable Levels of Tolerance to Water Stress (Drought) and Associated Biochemical Markers in Tunisian Barley Landraces. Molecules, 23(3), 613. https://doi.org/10.3390/molecules23030613