Expression Stabilities of Ten Candidate Reference Genes for RT-qPCR in Zanthoxylum bungeanum Maxim


Abstract
:1. Introduction
2. Results
2.1. geNorm Analysis
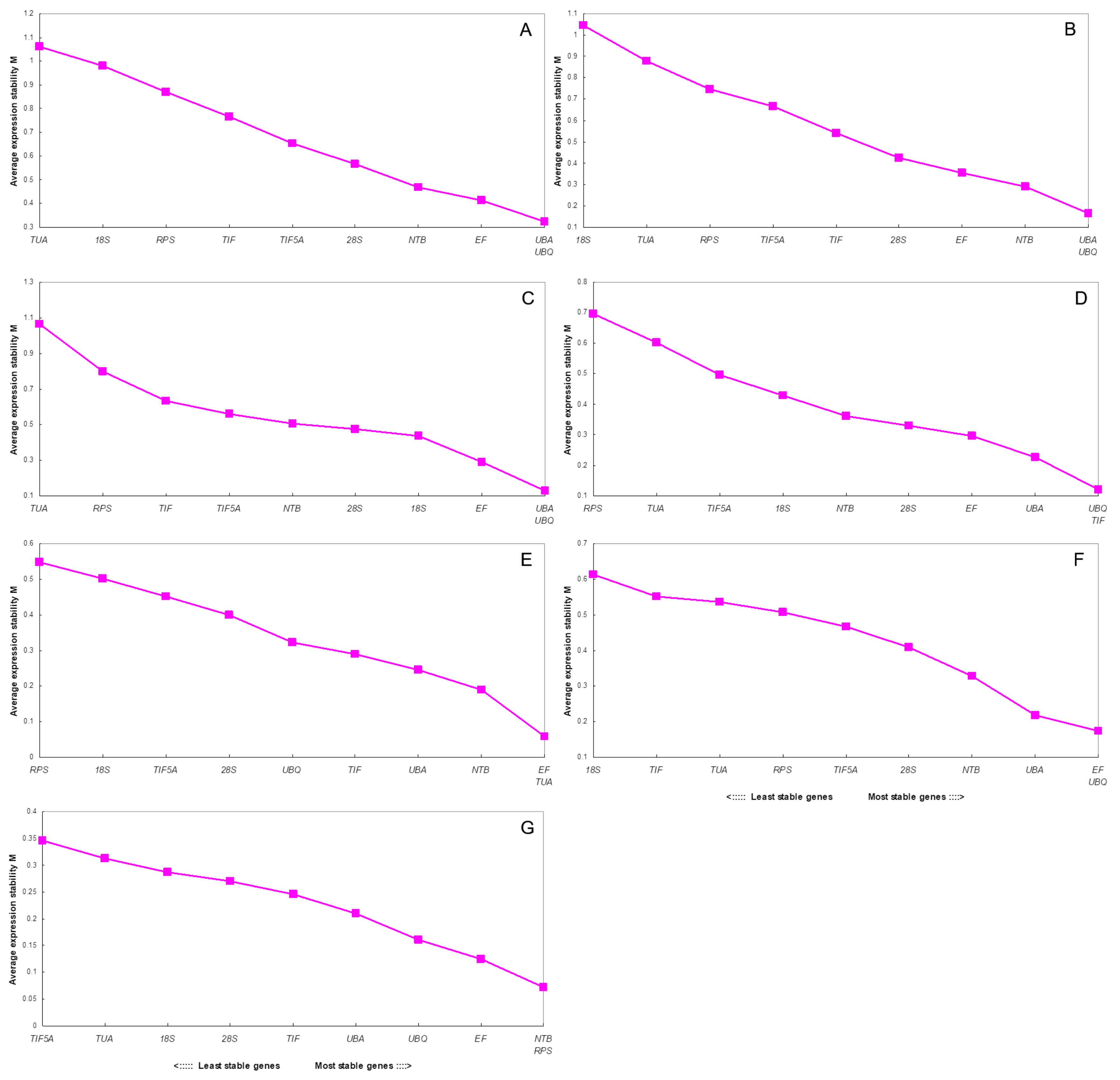
2.2. NormFinder Analysis
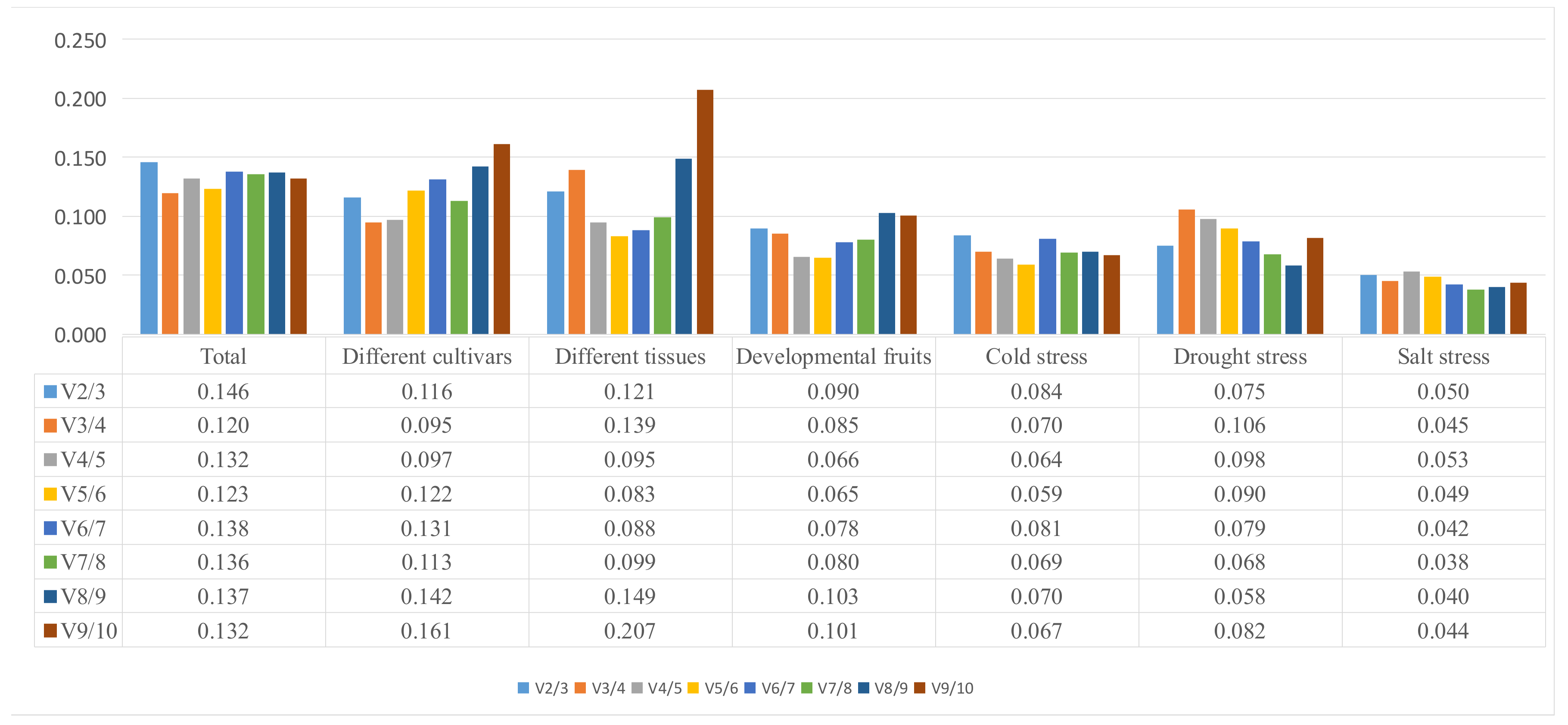
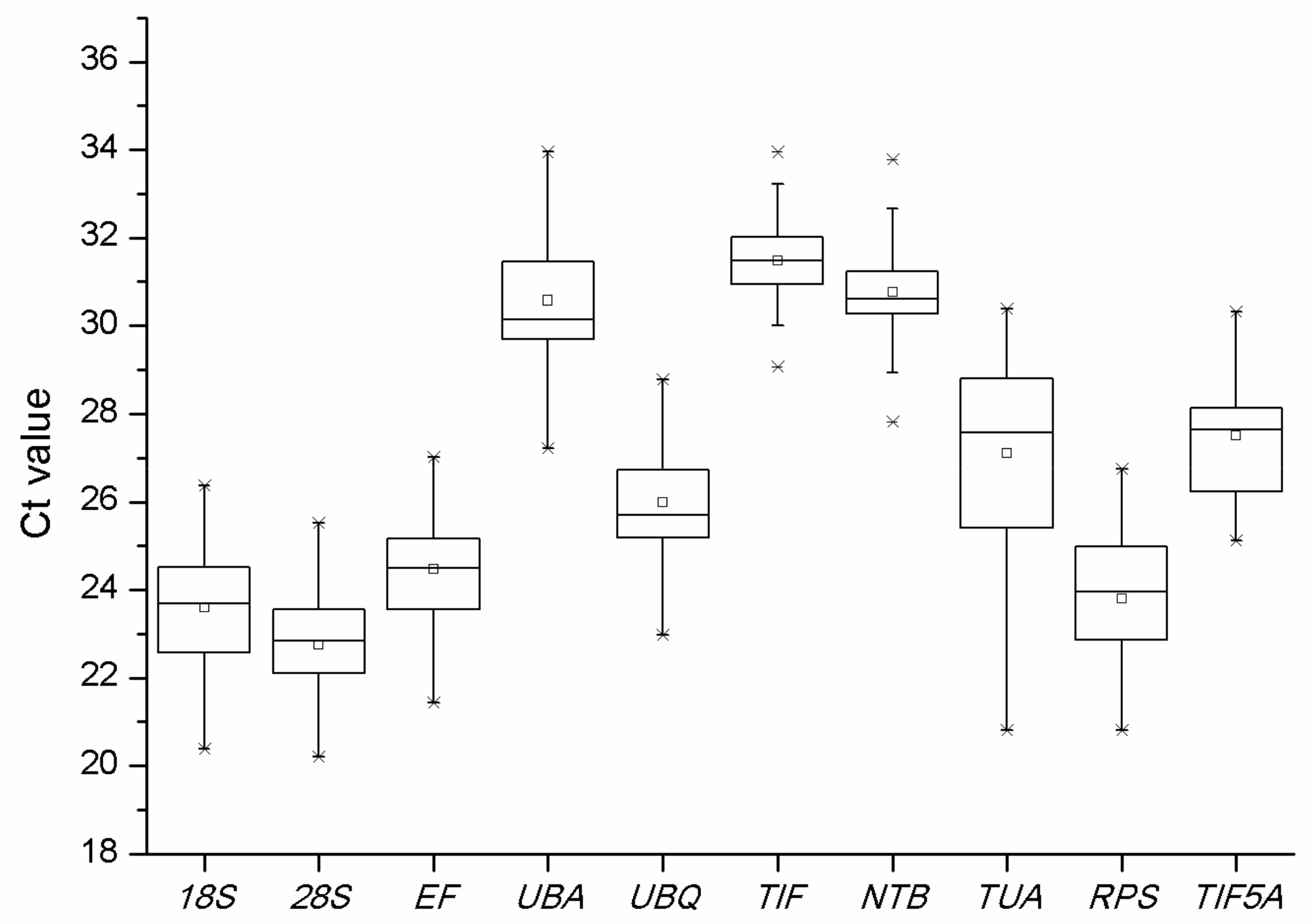
2.3. BestKeeper Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Plant Materials
4.1.2. Stress Treatments
4.2. Methods
4.2.1. Total RNA Extraction and cDNA Synthesis
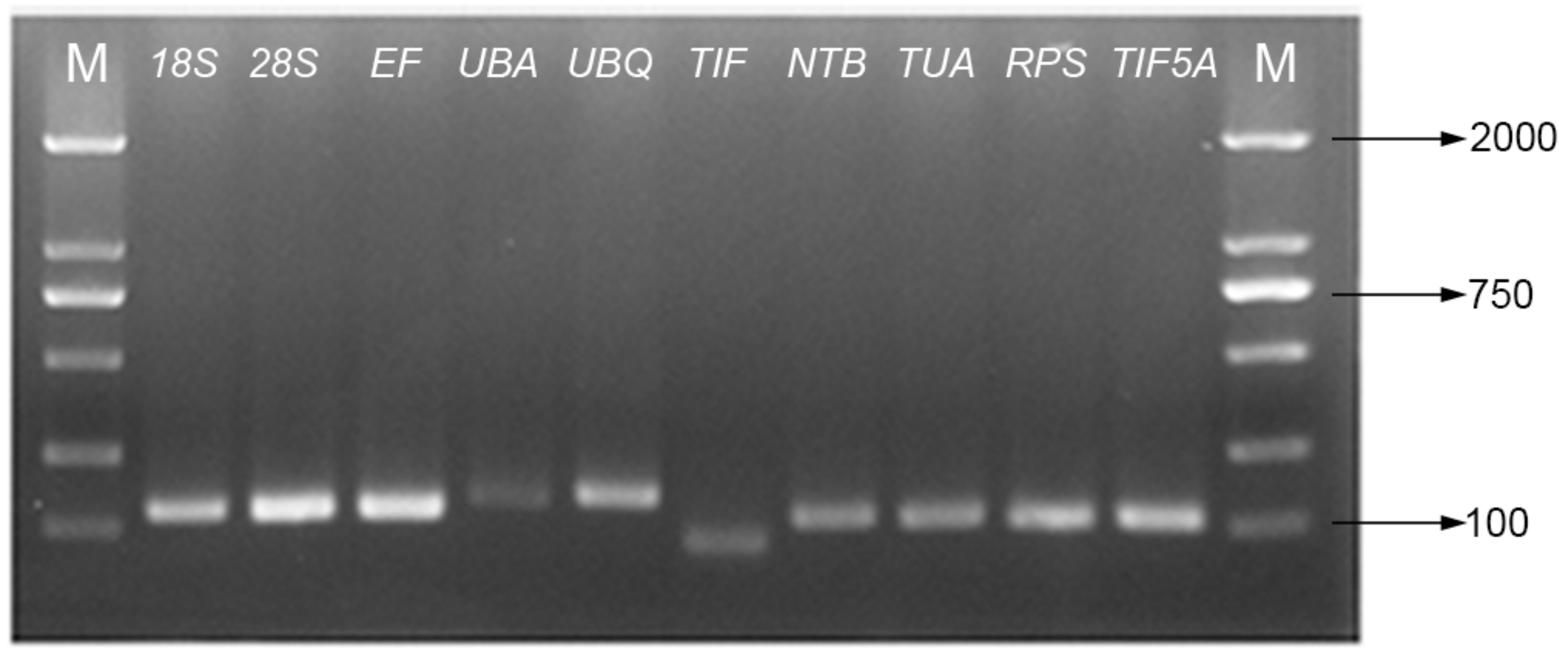
4.2.2. Primer Design and qPCR
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Feng, S.J.; Yang, T.X.; Liu, Z.S.; Chen, L.; Hou, N.; Wang, Y.; Wei, A. Genetic diversity and relationships of wild and cultivated Zanthoxylum germplasms based on sequence-related amplified polymorphism (SRAP) markers. Genet. Resour. Crop Evol. 2015, 62, 1193–1204. [Google Scholar] [CrossRef]
- Li, J.; Hui, T.; Wang, F.; Li, S.; Cui, B.; Cui, Y.; Peng, Z. Chinese red pepper (Zanthoxylum bungeanum Maxim.) leaf extract as natural antioxidants in salted silver carp (Hypophthalmichthys molitrix) in dorsal and ventral muscles during processing. Food Control 2015, 56, 9–17. [Google Scholar] [CrossRef]
- Huang, D.M.; Zhao, G.H.; Chen, Z.D.; Kan, J.Q. The food culture of Chinese prickly ash. China Condiment 2006, 1, 75–81. [Google Scholar]
- Wei, S.; Zhang, H.; Wang, Y.; Wang, L.; Li, X.; Wang, Y.; Zhang, H.; Xu, X.; Shi, Y. Ultrasonic nebulization extraction-heating gas flow transfer-headspace single drop microextraction of essential oil from pericarp of Zanthoxylum bungeanum Maxim. J. Chromatogr. A 2011, 29, 4599–4605. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, L. Acid-catalyzed esterification of Zanthoxylum bungeanum seed oil with high free fatty acids for biodiesel production. Bioresour. Technol. 2008, 99, 8995–8998. [Google Scholar] [CrossRef] [PubMed]
- Mafra, V.; Kubo, K.S.; Alves-Ferreira, M.; Ribeiro-Alves, M.; Stuart, R.M.; Boava, L.P.; Rodrigues, C.M.; Machado, M.A. Reference genes for accurate transcript normalization in citrus genotypes under different experimental conditions. PLoS ONE 2012, 7, e31263. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Zhao, J.; Liu, M. Expression Stabilities of Candidate Reference Genes for RT-qPCR in Chinese Jujube (Ziziphus jujuba Mill.) under a Variety of Conditions. PLoS ONE 2016, 11, e0154212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, J.; Li, X. Identification of appropriate reference genes for RT-qPCR analysis in Ziziphus jujuba Mill. Sci. Hortic. 2015, 197, 166–169. [Google Scholar] [CrossRef]
- Duan, M.; Wang, J.; Zhang, X.; Yang, H.; Wang, H.; Qiu, Y.; Song, J.; Guo, Y.; Li, X. Identification of Optimal Reference Genes for Expression Analysis in Radish (Raphanus sativus L.) and Its Relatives Based on Expression Stability. Front. Plant Sci. 2017, 8, 1605. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Qi, X.; Yan, H.; Huang, L.; Nie, G.; Zhang, X. Reference Gene Selection for Quantitative Real-Time Reverse-Transcriptase PCR in Annual Ryegrass (Lolium multiflorum) Subjected to Various Abiotic Stresses. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Li, P.; Luo, X.; Chang, T.; Li, J.; Zhao, J.; Xu, Y. Selection of appropriate reference genes for the detection of rhythmic gene expression via quantitative real-time PCR in Tibetan hulless barley. PLoS ONE 2018, 13, e0190559. [Google Scholar] [CrossRef] [PubMed]
- Han, P.-P.; Qin, L.; Li, Y.-S.; Liao, X.-S.; Xu, Z.-X.; Hu, X.-J.; Xie, L.-H.; Yu, C.-B.; Wu, Y.-F.; Liao, X. Identification of suitable reference genes in leaves and roots of rapeseed (Brassica napus L.) under different nutrient deficiencies. J. Integr. Agric. 2017, 16, 809–819. [Google Scholar] [CrossRef]
- Andersen CL, J.J.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A Model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Michael, W.; Pfaffl, A.T.C.P.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar]
- Xiao, X.M.J.; Wang, J.; Wu, X.; Li, P.; Yao, Y. Validation of suitable reference genes for gene expression analysis in the halophyte Salicornia europaea by real-time quantitative PCR. Front. Plant Sci. 2014, 788. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.D.P.K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Ma, J.; Guo, Q.; Li, X.; Wang, H.; Lu, M. Selection of Reference Genes for Quantitative Real-Time PCR in Bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shivhare, R.; Lata, C. Selection of suitable reference genes for assessing gene expression in pearl millet under different abiotic stresses and their combinations. Sci. Rep. 2016, 6, 23036. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Seo, P.J.; Yang, M.S.; Xiang, F.; Park, C.M. Exploring valid reference genes for gene expression studies in Brachypodium distachyon by real-time PCR. BMC Plant Biol. 2008, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Pollier, J.; Vanden Bossche, R.; Rischer, H.; Goossens, A. Selection and validation of reference genes for transcript normalization in gene expression studies in Catharanthus roseus. Plant Physiol. Biochem. 2014, 83, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Wang, F.; Jiang, Q.; Wang, G.L.; Tian, C.; Xiong, A.-S. Validation and Comparison of Reference Genes for qPCR Normalization of Celery (Apium graveolens) at Different Development Stages. Front. Plant Sci. 2016, 7, 313. [Google Scholar] [CrossRef]
- Maroufi, A. Selection of reference genes for real-time quantitative PCR analysis of gene expression in Glycyrrhiza glabra under drought stress. Biol. Plant. 2016, 60, 645–654. [Google Scholar] [CrossRef]
- Niu, L.; Tao, Y.B.; Chen, M.S.; Fu, Q.; Li, C.; Dong, Y.; Wang, X.; He, H.; Xu, Z.F. Selection of Reliable Reference Genes for Gene Expression Studies of a Promising Oilseed Crop, Plukenetia volubilis, by Real-Time Quantitative PCR. Int. J. Mol. Sci. 2015, 16, 12513–12530. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, X.; Chen, S.; Li, B.; Wang, X.; Ni, H. Selection of relatively exact reference genes for gene expression studies in flixweed (Descurainia sophia) by quantitative real-time polymerase chain reaction. Pestic. Biochem. Physiol. 2016, 127, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, X.; Chen, W.; Chen, J.; Lu, W.; Chen, L.; Fu, D. Evaluation of new reference genes in papaya for accurate transcript normalization under different experimental conditions. PLoS ONE 2012, 7, e44405. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Chen, L.L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.; Hao, B.; Lyon, M.P.; et al. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59–66. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Description | Primer Sequence (5′-3′) | GeneBank Accession Number | Amplicon Size | E(%) | R2 | |
|---|---|---|---|---|---|---|---|
| 18S | 18S ribosomal RNA gene | F-GCGGATTGTGCCAAGGAA | R-ATATCCGTTGCCGAGAGTCG | KC502933.1 | 100 | 98.9 | 0.998 |
| 28S | 28S ribosomal RNA gene | F-GTCGCCTTCTTTCGCTCTGTC | R-GGTTCACGGGATTCTGCAATT | HM851494.1 | 118 | 105.8 | 0.999 |
| EF | Elongation factor 1-alpha | F-GTGCTTGACTGCCACACCTC | R-TTCCGGCATCTCCATTCTTC | XM_006488084.2 | 107 | 92.8 | 0.993 |
| UBA | ubiquitin-60S ribosomal protein L40 | F-GACTTAGGGGAGGGATTATTGAG | R-TTCTTCTTCCGACAGTTTACAGC | XM_006481681.2 | 123 | 96.7 | 0.998 |
| UBQ | ubiquitin extension protein | F-TCGAAGATGGCCGTACATTG | R-TCCTCTAAGCCTCAGCACCA | AB906612.1 | 122 | 95.7 | 0.996 |
| TIF | Translation initiation factor | F-TTCCTCCCATTACGTTGCT | R-GCTGGTTACGGACTCTTTG | JK724818.1 | 165 | 93.4 | 0.987 |
| NTB | Nucleotide tract-binding protein | F-CTTTGGACGGGAGAAGTAT | R-TCACAGGAAGATAGGGATTT | XM_006482555.2 | 150 | 98.4 | 0.999 |
| TUA | tubulin | F-GTAGGCGGAGGAGATGATGC | R-GATGGAAGAGTTGGCGGTAAGT | GU911362.1 | 142 | 108.4 | 0.992 |
| RPS | ribosomal protein S16 | F-GAAATCCAAAAGCAAGGGG | R-AAAATGGCAGCAACACACC | KJ364714.1 | 135 | 92.8 | 0.998 |
| TIF5A | translation initiation factor 5A | F-ATATTGTGCCTTCCTCCC | R-GCCTCAGATCATCCTTGG | XM_006473932.1 | 122 | 94.8 | 0.995 |
| Genes | Total (Ct ± SD) | Different Cultivars (Ct ± SD) | Different Tissues (Ct ± SD) | Developmental Fruits (Ct ± SD) | Cold Stress (Ct ± SD) | Drought Stress (Ct ± SD) | Salt Stress (Ct ± SD) |
|---|---|---|---|---|---|---|---|
| 18S | 23.61 ± 1.52 | 23.14 ± 1.79 | 22.79 ± 1.82 | 23.25 ± 0.71 | 24.59 ± 1.03 | 25.31 ± 0.90 | 24.16 ± 0.53 |
| 28S | 22.76 ± 1.21 | 21.53 ± 0.59 | 21.40 ± 0.62 | 22.86 ± 0.25 | 24.22 ± 0.89 | 23.18 ± 0.60 | 23.77 ± 0.51 |
| EF | 24.46 ± 1.24 | 23.24 ± 0.26 | 23.20 ± 0.24 | 24.57 ± 0.49 | 26.49 ± 0.37 | 24.83 ± 0.56 | 24.71 ± 0.50 |
| UBA | 30.58 ± 1.70 | 28.88 ± 0.62 | 28.81 ± 0.58 | 30.85 ± 0.66 | 33.58 ± 0.35 | 30.86 ± 0.70 | 30.63 ± 0.78 |
| UBQ | 25.99 ± 1.45 | 24.58 ± 0.55 | 24.48 ± 0.55 | 26.10 ± 0.49 | 28.54 ± 0.24 | 26.30 ± 0.48 | 26.08 ± 0.52 |
| TIF | 31.48 ± 1.00 | 31.49 ± 0.57 | 31.49 ± 0.51 | 31.41 ± 0.50 | 33.07 ± 0.67 | 31.40 ± 0.75 | 30.77 ± 0.81 |
| NTB | 30.76 ± 1.22 | 29.77 ± 0.68 | 29.75 ± 0.61 | 31.01 ± 0.44 | 32.90 ± 0.60 | 30.71 ± 0.20 | 30.91 ± 0.51 |
| TUA | 27.11 ± 2.12 | 25.67 ± 1.50 | 25.53 ± 1.38 | 26.45 ± 0.66 | 29.78 ± 0.41 | 28.04 ± 0.53 | 28.32 ± 0.44 |
| RPS | 23.8 ± 1.67 | 21.39 ± 0.84 | 21.29 ± 0.79 | 24.13 ± 0.95 | 25.76 ± 0.95 | 23.75 ± 0.29 | 24.55 ± 0.56 |
| TIF5A | 27.5 ± 1.42 | 25.88 ± 0.63 | 25.8 ± 0.61 | 27.08 ± 0.58 | 29.69 ± 0.65 | 27.72 ± 0.47 | 28.44 ± 0.84 |
| Rank | Total | Different Cultivars | Different Tissues | Developmental Fruits | Cold Stress | Drought Stress | Salt Stress | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | |
| 1 | EF | 0.790 | UBQ | 0.731 | EF | 0.763 | 28S | 0.531 | TUA | 0.407 | TIF5A | 0.507 | RPS | 0.267 |
| 2 | UBQ | 0.819 | UBA | 0.764 | 18S | 0.794 | UBQ | 0.536 | EF | 0.418 | 28S | 0.512 | NTB | 0.274 |
| 3 | NTB | 0.867 | EF | 0.772 | UBA | 0.809 | TIF | 0.554 | NTB | 0.434 | UBQ | 0.568 | UBQ | 0.285 |
| 4 | 28S | 0.895 | 28S | 0.817 | UBQ | 0.818 | EF | 0.559 | UBA | 0.544 | NTB | 0.574 | EF | 0.315 |
| 5 | UBA | 0.937 | NTB | 0.851 | 28S | 0.834 | UBA | 0.610 | 28S | 0.544 | TUA | 0.584 | 28S | 0.341 |
| 6 | TIF5A | 0.994 | TIF | 1.058 | NTB | 0.887 | NTB | 0.633 | TIF | 0.558 | RPS | 0.587 | UBA | 0.343 |
| 7 | TIF | 1.229 | TIF5A | 1.140 | TIF5A | 0.955 | TIF5A | 0.711 | TIF5A | 0.585 | TIF | 0.606 | 18S | 0.365 |
| 8 | RPS | 1.315 | RPS | 1.204 | TIF | 1.134 | 18S | 0.807 | UBQ | 0.591 | EF | 0.662 | TIF | 0.384 |
| 9 | 18S | 1.381 | TUA | 1.397 | RPS | 1.513 | TUA | 0.952 | 18S | 0.670 | UBA | 0.674 | TUA | 0.408 |
| 10 | TUA | 1.395 | 18S | 1.706 | TUA | 2.132 | RPS | 1.077 | RPS | 0.736 | 18S | 0.865 | TIF5A | 0.485 |
| Rank | Total | Different Cultivars | Different Tissues | Developmental Fruits | Cold Stress | Drought Stress | Salt Stress | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability Value | Gene | Stability Value | Gene | Stability Value | Gene | Stability Value | Gene | Stability Value | Gene | Stability Value | Gene | Stability Value | |
| 1 | UBA | 0.001 | UBA | 0.000 | TIF5A | 0.005 | UBA | 0.000 | UBA | 0.003 | TIF5A | 0.003 | UBQ | 0.004 |
| 2 | NTB | 0.001 | NTB | 0.000 | UBA | 0.017 | TIF | 0.000 | TUA | 0.003 | TUA | 0.005 | TIF5A | 0.005 |
| 3 | TIF | 0.001 | TIF | 0.001 | UBQ | 0.018 | NTB | 0.001 | TIF5A | 0.003 | NTB | 0.021 | TUA | 0.015 |
| 4 | TIF5A | 0.008 | TUA | 0.013 | NTB | 0.022 | 28S | 0.006 | NTB | 0.004 | TIF | 0.024 | NTB | 0.023 |
| 5 | UBQ | 0.025 | UBQ | 0.014 | TIF | 0.023 | UBQ | 0.006 | 28S | 0.004 | 28S | 0.024 | UBA | 0.023 |
| 6 | EF | 0.112 | TIF5A | 0.024 | 28S | 0.023 | TIF5A | 0.011 | TIF | 0.005 | UBA | 0.024 | TIF | 0.025 |
| 7 | TUA | 0.119 | EF | 0.025 | EF | 0.079 | EF | 0.056 | UBQ | 0.025 | UBQ | 0.031 | 28S | 0.026 |
| 8 | 28S | 0.124 | 28S | 0.243 | 18S | 0.127 | TUA | 0.059 | EF | 0.091 | EF | 0.129 | 18S | 0.069 |
| 9 | 18S | 0.309 | RPS | 0.333 | TUA | 0.269 | 18S | 0.269 | 18S | 0.117 | 18S | 0.143 | EF | 0.113 |
| 10 | RPS | 0.338 | 18S | 0.403 | RPS | 0.376 | RPS | 0.318 | RPS | 0.127 | RPS | 0.210 | RPS | 0.117 |
| Rank | Total | Different Cultivars | Different Tissues | Developmental Fruits | Cold Stress | Drought Stress | SALT STRESS | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | CV ± SD | Gene | CV ± SD | Gene | CV ± SD | Gene | CV ± SD | Gene | CV ± SD | Gene | CV ± SD | Gene | CV ± SD | |
| 1 | TIF | 2.30 ± 0.72 | EF | 0.92 ± 0.21 | TIF | 2.41 ± 0.74 | 28S | 0.89 ± 0.20 | UBQ | 0.59 ± 0.17 | NTB | 0.55 ± 0.17 | TUA | 1.00 ± 0.28 |
| 2 | NTB | 2.77 ± 0.85 | TIF | 1.19 ± 0.37 | NTB | 2.65 ± 0.79 | NTB | 1.08 ± 0.33 | UBA | 0.80 ± 0.27 | RPS | 0.91 ± 0.22 | NTB | 1.27 ± 0.39 |
| 3 | EF | 3.90 ± 0.95 | UBQ | 1.48 ± 0.36 | 28S | 3.98 ± 0.86 | TIF | 1.30 ± 0.41 | TUA | 1.01 ± 0.30 | UBQ | 1.26 ± 0.33 | EF | 1.56 ± 0.39 |
| 4 | TIF5A | 3.96 ± 1.09 | UBA | 1.50 ± 0.43 | TIF5A | 4.04 ± 1.09 | UBQ | 1.51 ± 0.40 | EF | 1.04 ± 0.27 | TUA | 1.35 ± 0.38 | UBQ | 1.71 ± 0.45 |
| 5 | 28S | 4.16 ± 0.91 | NTB | 1.56 ± 0.47 | UBA | 4.13 ± 1.21 | TIF5A | 1.58 ± 0.43 | NTB | 1.33 ± 0.44 | TIF5A | 1.46 ± 0.40 | 18S | 1.77 ± 0.43 |
| 6 | UBQ | 4.29 ± 1.12 | TIF5A | 2.00 ± 0.52 | 18S | 4.16 ± 0.91 | EF | 1.60 ± 0.39 | TIF | 1.57 ± 0.52 | EF | 1.47 ± 0.36 | RPS | 1.78 ± 0.44 |
| 7 | UBA | 4.38 ± 1.34 | 28S | 2.20 ± 0.47 | EF | 4.21 ± 0.99 | TUA | 1.69 ± 0.45 | TIF5A | 1.74 ± 0.52 | UBA | 1.64 ± 0.51 | 28S | 1.82 ± 0.43 |
| 8 | 18S | 5.06 ± 1.19 | RPS | 2.76 ± 0.59 | UBQ | 4.62 ± 1.15 | UBA | 1.84 ± 0.57 | 28S | 2.70 ± 0.66 | 28S | 2.01 ± 0.47 | UBA | 2.09 ± 0.64 |
| 9 | RPS | 5.46 ± 1.30 | TUA | 4.67 ± 1.20 | RPS | 6.52 ± 1.55 | 18S | 2.53 ± 0.59 | RPS | 2.72 ± 0.70 | TIF | 2.04 ± 0.64 | TIF5A | 2.10 ± 0.60 |
| 10 | TUA | 6.06 ± 1.64 | 18S | 6.41 ± 1.48 | TUA | 8.61 ± 2.19 | RPS | 3.31 ± 0.80 | 18S | 2.98 ± 0.73 | 18S | 2.22 ± 0.56 | TIF | 2.26 ± 0.69 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fei, X.; Shi, Q.; Yang, T.; Fei, Z.; Wei, A. Expression Stabilities of Ten Candidate Reference Genes for RT-qPCR in Zanthoxylum bungeanum Maxim. Molecules 2018, 23, 802. https://doi.org/10.3390/molecules23040802
Fei X, Shi Q, Yang T, Fei Z, Wei A. Expression Stabilities of Ten Candidate Reference Genes for RT-qPCR in Zanthoxylum bungeanum Maxim. Molecules. 2018; 23(4):802. https://doi.org/10.3390/molecules23040802
Chicago/Turabian StyleFei, Xitong, Qianqian Shi, Tuxi Yang, Zhaoxue Fei, and Anzhi Wei. 2018. "Expression Stabilities of Ten Candidate Reference Genes for RT-qPCR in Zanthoxylum bungeanum Maxim" Molecules 23, no. 4: 802. https://doi.org/10.3390/molecules23040802
APA StyleFei, X., Shi, Q., Yang, T., Fei, Z., & Wei, A. (2018). Expression Stabilities of Ten Candidate Reference Genes for RT-qPCR in Zanthoxylum bungeanum Maxim. Molecules, 23(4), 802. https://doi.org/10.3390/molecules23040802