Progress in Metabonomics of Type 2 Diabetes Mellitus

 and
and
Abstract
:1. Introduction
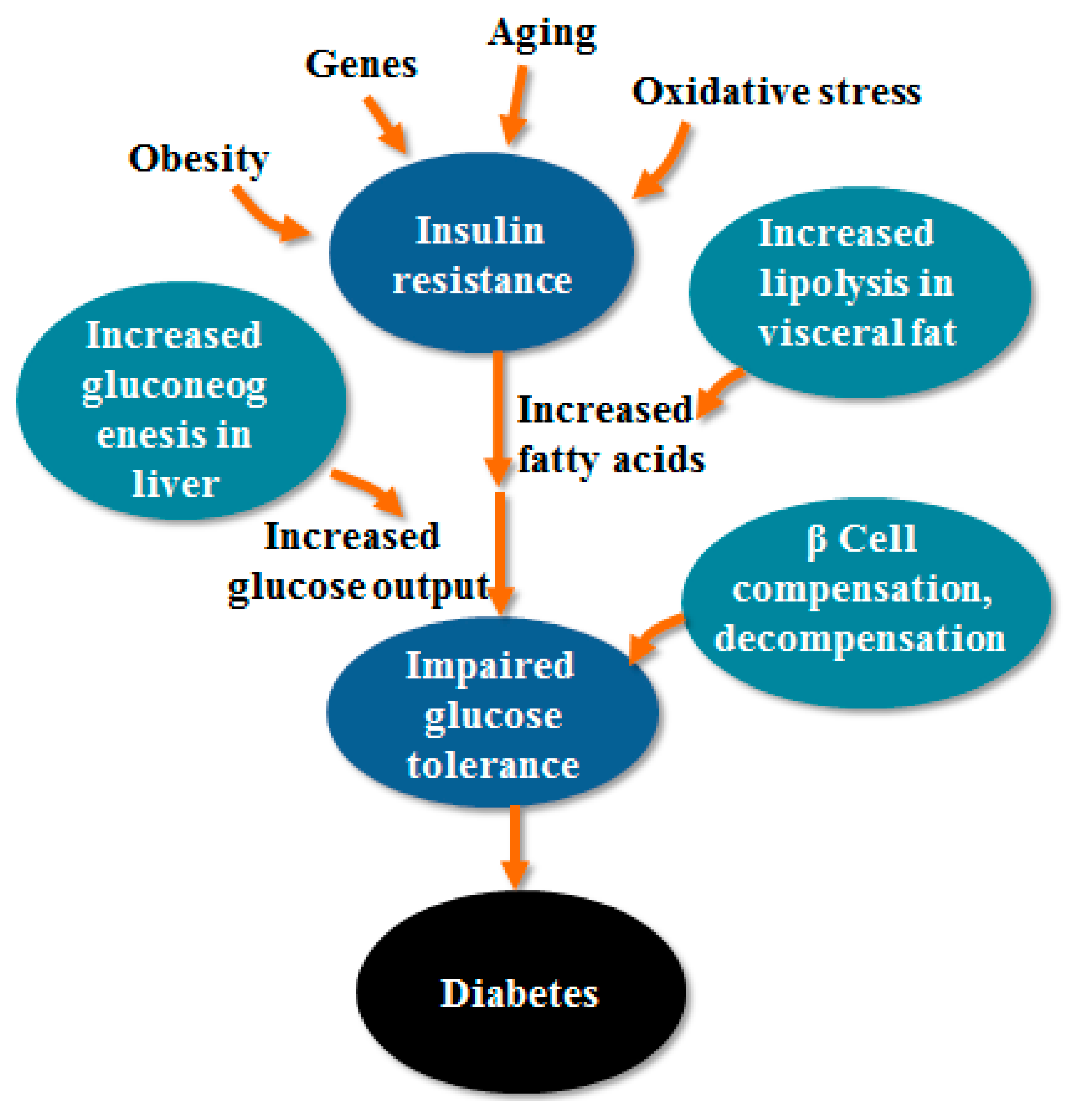
2. The Pathogenesis of T2DM
2.1. IR
2.2. Impaired Function of Islet Cells
2.3. Oxidative Stress
2.4. Gene Factors
2.5. Obesity
2.6. Other Pathogenesis
2.6.1. Inflammation
2.6.2. Hepatitis B Virus
3. Metabonomics Research Methods and Techniques
3.1. NMR
- (1)
- The NMR method hardly requires sample pretreatment, enables non-invasive and unbiased detection of the sample, and has good objectivity and reproducibility [29].
- (2)
- NMR is almost non-destructive to the sample (the stability of some samples is limited), which can be used for in vitro [29].
- (3)
- Peaks in the NMR spectrum can represent a certain metabolite, which means an NMR spectrum can provide qualitative and quantitative information on a large number of metabolites in the organism.
- (4)
- 1H-NMR responds to compounds containing H, which can complete the detection of most metabolites and meet the goal of detecting as many metabolites in metabonomics as possible.
- (5)
- High-flux NMR techniques with the use of automated liquid handling procedures takes only a few minutes to detect large amounts of metabolite information [30].
- (6)
- Although NMR spectrometers and its recurring expenditures are expensive, NMR is very informative, the cost of a single sample is low, and ultimately the cost of analysis is generally reduced.
- (7)
- NMR can provide rich molecular information including metabolite composition, concentration, molecular dynamics, interactions, pH, and structure.
3.2. MS
- (1)
- A large amount of sample preparations is required, which are destructive to the sample and, therefore, cannot be studied in vivo or in situ.
- (2)
- There is a need for knowledge of the sample and high recurring costs and the equipment is also quite expensive.
4. Advances in the Pathogenesis of T2DM Based on Metabonomics
4.1. Biomarker
4.1.1. Biomarkers Related to Amino Acid Metabolism
4.1.2. Biomarkers Related to Lipid Metabolism
4.1.3. Biomarkers Related to Carbohydrate Metabolism
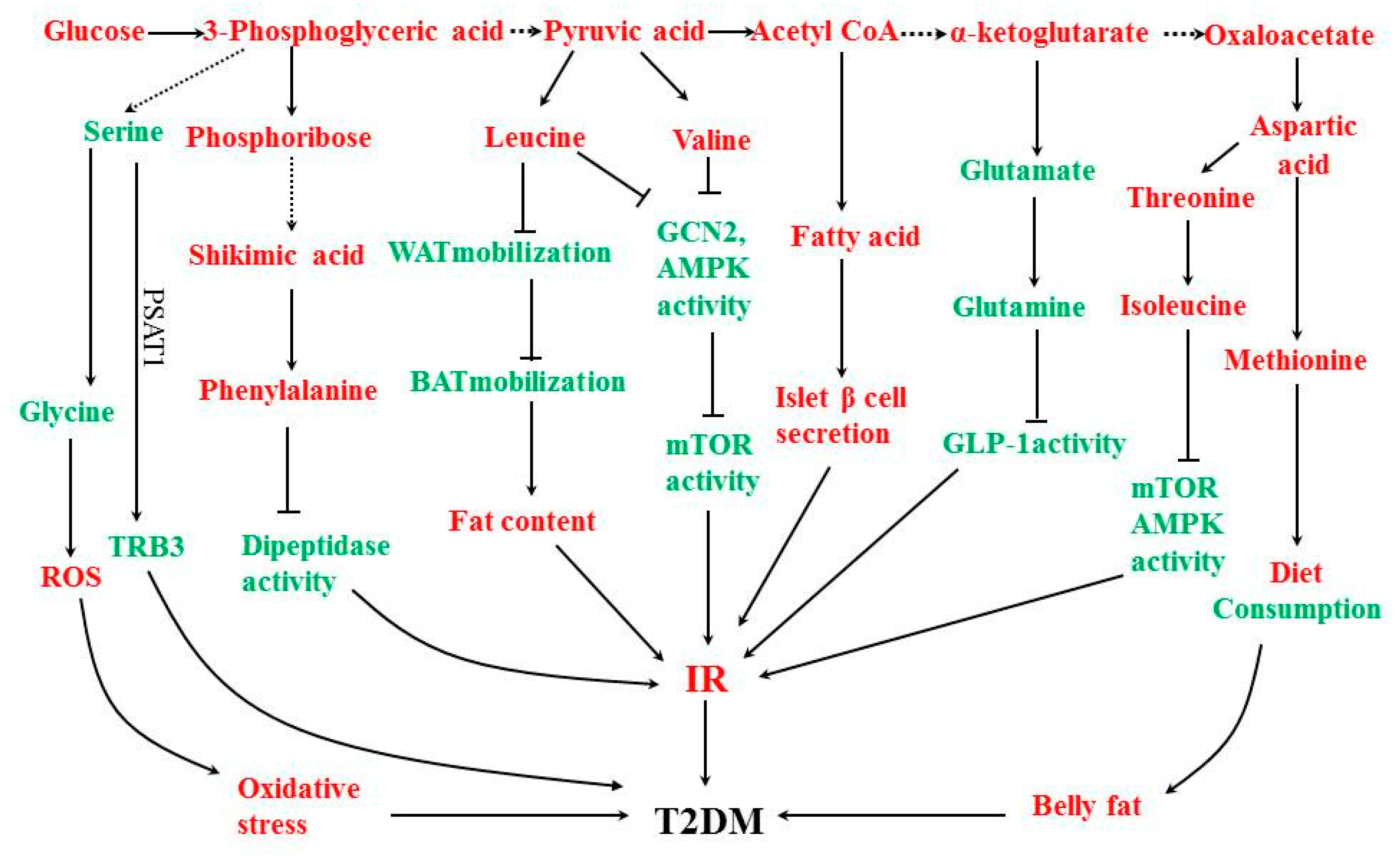
4.2. Metabolic Pathway
4.2.1. Serine Amino Acid Biosynthetic Pathway
4.2.2. Phosphate Pentose Pathway and Aromatic Amino Acid Biosynthesis Pathway
4.2.3. Alanine Amino Acid Biosynthesis Pathway
4.2.4. Fatty Acid Biosynthesis Pathway
4.2.5. Glutamic Acid Amino Acid Biosynthetic Pathway
4.2.6. Aspartate Amino Acid Biosynthetic Pathway
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF diabetes atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gao, P.; Zhang, M.; Huang, Z.; Zhang, D.; Deng, Q.; Li, Y.; Zhao, Z.; Qin, X.; Jin, D.; et al. Prevalence and ethnic pattern of diabetes and prediabetes in China in 2013. JAMA 2017, 317, 2515–2523. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Snehalatha, C.; Nanditha, A. Classification and diagnosis of diabetes. In Diabetes Care, 5th ed.; Holt, R.I.G., Cockram, C., Eds.; John Wiley & Sons, Ltd.: West Sussex, UK, 2016. [Google Scholar]
- Milburn, M.V.; Lawton, K.A. Application of metabolomics to diagnosis of insulin resistance. Annu. Rev. Med. 2013, 64, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Jeong, G.H.; Lee, I.K.; Yoon, Y.R.; Liu, K.H.; Gu, N.; Shin, K.H. A pharmacometabolomic approach to predictResponse to metformin in early-phase type 2 diabetes mellitus patients. Molecules 2018, 23, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Song, C.; Huo, H. Research of traditional Chinese medicine in treating type 2 diabetes and insulin resistance. World Chin. Med. 2015, 135–137. [Google Scholar]
- Devarshi, P.P.; Mcnabney, S.M.; Henagan, T.M. Skeletal muscle nucleo-mitochondrial crosstalk in obesity and type 2 diabetes. Int. J. Mol. Sci. 2017, 18, 831–850. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Duan, C.; Li, L. The pathogenesis mechanisms of type 2 diabetes mellitus. Med. Recapitulate 2015, 21, 3935–3938. [Google Scholar]
- Unger, R.H.; Orci, L. Glucagon and the A cell: Physiology and pathophysiology. In The Leukemias: Etiology, Pathophysiology, and Treatment; Unger, R.H., Orci, L., Eds.; Academic Press: Amsterdam, The Netherlands, 1981; pp. 443–449. [Google Scholar]
- Goodarzi, M.T.; Navidi, A.A.; Rezaei, M.; Babahmadi-Rezaei, H. Oxidative damage to DNA and lipids: correlation with protein glycation in patients with type 1 diabetes. J. Clin. Lab. Anal. 2010, 24, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Hooper, P.L.; Balogh, G.; Rivas, E.; Kavanagh, K.; Vigh, L. The importance of the cellular stress response in the pathogenesis and treatment of type 2 diabetes. Cell Stress Chaperon. 2014, 19, 447–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stumvoll, M.; Goldstein, B. Pathogenesis of type 2 diabetes. Endoc. Res. 2007, 32, 19–37. [Google Scholar] [CrossRef]
- Grant, S.F.; Thorleifsson, G.; Reynisdottir, I. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat. Genet. 2006, 38, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Miyake, K.; Horikawa, Y. Variants in KCNQ1 are associated with susceptibility to type 2 diabetes mellitus. Nat. Genet. 2008, 40, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Auwerx, J. PPARγ, the ultimate thrifty gene. Diabetologia 1999, 42, 1033–1049. [Google Scholar] [CrossRef] [PubMed]
- Arrington, D.D.; Van Vleet, T.R.; Schnellmann, R.G. Calpain 10: A mitochondrial calpain and its role in calcium-induced mitochondrial dysfunction. Am. J Physiol. Renal. Physiol. 2006, 291, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Urbanová, M.; Haluzik, M. The role of adipose tissue in pathogenesis of type 2 diabetes mellitus. Cesk. Fysiol. 2015, 64, 73–78. [Google Scholar] [PubMed]
- Erion, D.M.; Park, H.J.; Lee, H.Y. The role of lipids in the pathogenesis and treatment of type 2 diabetes and associated co-morbidities. BMB Rep. 2016, 49, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltiel, A.R. New perspectives into the molecular pathogenesis and treatment of type 2 diabetes. Cell 2001, 104, 517–529. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Zatalia, S.R.; Sanusi, H. The role of antioxidants in the pathophysiology, complications, and management of diabetes mellitus. Acta. Med. Indones. 2013, 45, 141–147. [Google Scholar] [PubMed]
- Akash, H.; Sajid, M.; Rehman, K. Tumor necrosis factor-alpha: Role in development of insulin resistance and pathogenesis of type 2 diabetes mellitus. J. Cell Biochem. 2018, 119, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Sosa, M.; Cabellos-Avelar, T.; Sanchez-Zamora, Y.; Juárez-Avelar, I.; García-Reyes, E.; Lira-León, A.; Benítez-Flores, J.C.; Pacheco-Fernández, T.; Hiriart, M.; Gutiérrez-Cirlos, E.B. Proinflammatory cytokine MIF plays a role in the pathogenesis of type-2 diabetes mellitus, but does not affect hepatic mitochondrial function. Cytokine 2017, 99, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Coope, A.; Torsoni, A.S.; Velloso, L.A. Mechanisms in endocrinology: metabolic and inflammatory pathways on the pathogenesis of type 2 diabetes. Eur. J. Endocrinol. 2016, 174, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Meshkani, R.; Vakili, S. Tissue resident macrophages: Key players in the pathogenesis of type 2 diabetes and its complications. Clin. Chim. Acta 2016, 462, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Custro, N.; Carroccio, A.; Ganci, A.; Scafidi, V.; Campagna, P.; Di Prima, L.; Montalto, G. Glycemic homeostasis in chronic viral hepatitis and liver cirrhosis. Diabetes Metab. 2001, 27, 476–481. [Google Scholar] [PubMed]
- Soverini, V.; Persico, M.; Bugianesi, E.; Forlani, G.; Salamone, F.; Massarone, M.; Mura, V.L.; Mazzotti, A.; Bruno, A.; Marchesini, G. HBV and HCV infection in type 2 diabetes mellitus: a survey in three diabetes units in different Italian areas. Acta Diabetol. 2011, 48, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. “Metabonomics”: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Li, A.P.; Li, Z.Y.; Sun, H.F.; Li, K.; Qin, X.M.; Du, G.H. Comparison of two different astragali radixby a1H-NMR-based metabolomic approach. J. Proteome Res. 2015, 14, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.H.M. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods Mol. Biol. 2015, 1277, 161–193. [Google Scholar] [PubMed]
- Zhang, A.H.; Sun, H.; Yan, G.L.; Yuan, Y.; Han, Y.; Wang, X.J. Metabolomics study of type 2 diabetes using ultra-performance LC-ESI/quadrupole-TOF high-definition MS coupled with pattern recognition methods. J. Phsyiol. Biochem. 2014, 70, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Römischmargl, W.; Prehn, C.; Bogumil, R.; Röhring, C.; Suhre, K.; Adamski, J. Procedure for tissue sample preparation and metabolite extraction for high-throughput targeted metabolomics. Metabolomics 2012, 8, 133–142. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Felig, P.; Marliss, E.C.G., Jr. Plasma amino acid levels and insulin secretion in obesity. N. Engl. J. Med. 1969, 281, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Felig, P.; Wahren, J.; Hendler, R.; Brundin, T. Splanchnic glucose and amino acid metabolism in obesity. J. Clin. Investig. 1974, 53, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Luetscher, J.A. The metabolism of amino acids in dlabetes. J. Clin. Investig. 1942, 21, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Huffman, K.M.; Shah, S.H.; Stevens, R.D.; Bain, J.R.; Muehlbauer, M.; Slentz, C.A.; Tanner, C.J.; Kuchibhatla, M.; Houmard, J.A.; Newgard, C.B.; et al. Relationships between circulating metabolic intermediates and insulin action in overweight to obese, inactive men and women. Diabetes Care 2009, 32, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Tai, E.S.; Tan, M.L.S.; Stevens, R.D.; Low, Y.L.; Muehlbauer, M.J.D.; Goh, L.M.; Ilkayeva, O.R.; Wenner, B.R.; Bain, J.R.; Lee, J.J.M.; et al. Insulin resistance is associated with a metabolic profile of altered protein metabolism in Chinese and Asian-Indian men. Diabetologia 2010, 53, 757–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guaschferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F. Metabolomics in prediabetes and diabetes: A systematic review and meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menni, C.; Fauman, E.; Erte, I.; Perry, J.R.B.; Kastenmüller, G.; Shin, S.Y.; Petersen, A.K.; Hyde, C.; Psatha, M.; Ward, K.J.; et al. Biomarkers for type 2 diabetes and impaired fasting glucose using a nontargeted metabolomics approach. Diabetes 2013, 62, 4270–4276. [Google Scholar] [CrossRef] [PubMed]
- Salek, R.M.; Maguire, M.L.; Bentley, E.; Rubtsov, D.V.; Hough, T.; Cheeseman, M.; Nunez, D.; Sweatman, B.C.; Haselden, J.N.; Cox, R.D.; et al. A metabolomic comparison of urinary changes in type 2 diabetes in mouse, rat, and human. Physiol. Genom. 2007, 29, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, Y.; Hao, F.; Zhou, X.; Han, X.; Tang, H.; Ji, L. Human serum metabonomic analysis reveals progression axes for glucose intolerance and insulin resistance statuses. J. Proteome Res. 2009, 8, 5188–5195. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zhao, T.; Wang, X.; Qiu, Y.; Su, M.; Jia, W.; Jia, W. Metabonomic variations in the drug-treated type 2 diabetes mellitus patients and healthy volunteers. J. Proteome Res. 2009, 8, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Mochida, T.; Tanaka, T.; Shiraki, Y.; Tajiri, H.; Matsumoto, S.; Shimbo, K.; Ando, T.; Nakamura, K.; Okamoto, M.; Endo, F. Time-dependent changes in the plasma amino acid concentration in diabetes mellitus. Mol. Genet. Metab. 2011, 103, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Merino, J.; Leong, A.; Liu, C.T.; Porneala, B.; Walford, G.A.; Grotthuss, M.; Wang, T.J.; Flannick, J.; Dupuis, J.; Levy, D.; et al. Metabolomics insights into early type 2 diabetes pathogenesis and detection in individuals with normal fasting glucose. Diabetologia 2018, 61, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Stefan, N.; Yu, Z.; Mühlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Häring, H.U.; Hrabě de Angelis, M.; Peters, A.; et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Wang-Sattler, R.; Yu, Z.; Herder, C.; Messias, A.C.; Floegel, A.; He, Y.; Heim, K.; Campillos, M.; Holzapfel, C.; Thorand, B.; et al. Novel biomark-ers for pre-diabetes identified by metabolomics. Mol. SystBiol. 2012, 8, 615–626. [Google Scholar]
- Phillips, J.D.; Kushner, J.P. Fast track to the porphyrias. Nat. Med. 2005, 11, 1049–1050. [Google Scholar] [CrossRef] [PubMed]
- Wittenbecher, C.; Mühlenbruch, K.; Kröger, J.; Jacobs, S.; Kuxhaus, O.; Floegel, A.; Fritsche, A.; Pischon, T.; Prehn, C.; Adamski, J.; et al. Amino acids, lipid metabolites, and ferritin as potential mediators linking red meat consumption to type 2 diabetes. Am. J. Clin. Nutr. 2015, 101, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wang, S.; Puhl, M.D.; Jiang, X.; Hyrc, K.L.; Laciny, E.; Wallendorf, M.J.; Pappan, K.L.; Coyle, J.T.; Wice, B.M.; et al. Global biochemical profiling identifies β-hydroxypyruvate as a potential mediator of type 2 diabetes in mice and humans. Diabetes 2015, 64, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Ngo, D.; Psychogios, N.; Wang, T.J.; Ngo, D.; Psychogios, N.; Dejam, A.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; et al. 2-Aminoadipic acid is a biomarker for diabetes risk. J. Clin. Investig. 2013, 123, 4309–4317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiehn, O.; Garvey, W.T.; Newman, J.W.; Lok, K.H.; Hoppel, C.L.; Adams, S.H. Plasma metabolomic profiles reflective of glucose homeostasis in non-diabetic and type 2 diabetic obese African-American women. PLoS ONE 2010, 5, 15234. [Google Scholar] [CrossRef] [PubMed]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.P.; Mitchell, M.W.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. α-Hydroxybutyrate is an early biomarker of insulin resistance and glucose intolerance in a nondiabetic population. PLoS ONE 2010, 5, 10883. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, Z.; Lu, X.; Yang, X.; Yin, P.; Kong, H.; Yu, Y.; Xu, G. Comprehensive two-dimensional gas chromatography/time-of-flight mass spectrometry for metabonomics: Biomarker discovery for diabetes mellitus. Anal. Chim. Acta 2009, 633, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Mosharov, E.; Cranford, M.R.C.; Banerjee, R. The Quantitatively Important Relationship between Homocysteine Metabolism and Glutathione Synthesis by the Transsulfuration Pathway and Its Regulation by Redox Changes. Biochemistry 2000, 39, 13005–13011. [Google Scholar] [CrossRef] [PubMed]
- Persa, C.; Pierce, A.; Ma, Z.; Kabil, O.; Loua, M.F. The presence of a transsulfuration pathway in the lens: A new oxidative stress defense system. Exp. Eye Res. 2004, 79, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhou, J.; Bao, Y.; Chen, T.; Zhang, Y.; Zhao, A.; Qiu, Y.; Xie, G.; Wang, C.; Jia, W.; et al. Serum metabolic signatures of fulminant type 1 diabetes. J. Proteome Res. 2012, 11, 4705–4711. [Google Scholar] [CrossRef] [PubMed]
- Kostolanska, J.; Jakuš, V.; Barak, L. HbA1c and serum levels of advanced glycation and oxidation protein products in poorly and well controlled children and adolescents with type 1 diabetes mellitus. J. Pediatr. Endocr. Metab. 2009, 22, 433–442. [Google Scholar] [CrossRef]
- Ferrannini, E.; Natali, A.; Camastra, S.; Nannipieri, M.; Mari, A.; Adam, K.P.; Milburn, M.V.; Kastenmüller, G.; Adamski, J.; Tuomi, T.; et al. Early metabolic markers of the development of dysglycemia and type 2 diabetes and their physiological significance. Diabetes 2013, 62, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Rhee, E.P.; Larson, M.G.; Lewis, G.D.; McCabe, E.L.; Shen, D.; Palma, M.J.; Roberts, L.D.; Dejam, A.; Souza, A.L.; et al. Metabolite Profiling Identifies Pathways Associated with Metabolic Risk in Humans. Circulation 2012, 125, 2222–2231. [Google Scholar] [CrossRef] [PubMed]
- Kannel, W.B. Lipids, diabetes, and coronary heart disease: insights from the framingham study. Am. Heart J. 1985, 110, 1100–1107. [Google Scholar] [CrossRef]
- Krauss, R.M.; Siri, P.W. Dyslipidemia in type 2 diabetes. Med. Clin. N. Am. 2004, 88, 897. [Google Scholar] [CrossRef] [PubMed]
- Taskinen, M.R. Diabetic dyslipidaemia: From basic research to clinical practice. Diabetologia 2003, 46, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Cao, H.; King, I.B.; Lemaitre, R.N.; Song, X.; Siscovick, D.S.; Hotamisligil, G.S. Circulating palmi-toleic acid and risk of metabolic abnormalities and new-onset diabetes. Am. J. Clin. Nutr. 2010, 92, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Li, Y.; Guan, C.M.; Li, K.; Wang, C.; Feng, R.; Sun, C. Free fatty acid metabolic profile andbiomarkers of isolated post-challenge diabetes and type 2 diabetes mellitus based on GC-MS and multivariate statistical analysis. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2010, 878, 2817–2825. [Google Scholar] [CrossRef] [PubMed]
- Suhre, K.; Meisinger, C.; Döring, A.; Altmaier, E.; Belcredi, P.; Gieger, C.; Chang, D.; Milburn, M.V.; Gall, W.E.; Weinberger, K.M.; et al. Metabolic footprint of diabetes: a multiplatform metabolomics study in an epidemiological setting. PLoS ONE 2010, 5, 13953. [Google Scholar] [CrossRef] [PubMed]
- Blaak, E.E.; van Aggel-Leijssen, D.P.C.; Wagenmakers, A.J.M.; Saris, W.H.; van Baak, M.A. Impaired oxidation of plasma-derived fatty acids in type 2 diabetic subjects during moderate-intensity exercise. Diabetes 2000, 49, 2102–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, D.E. Skeletal muscle fat oxidation: timing and flexibility are everything. The J. Clin. Investig. 2005, 115, 1699–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, D.E.; Goodpaster, B.; Wing, R.R.; Simoneau, J.A. Skeletal muscle fatty acid metabolism in association with insulin resistance, obesity, and weight loss. Am. J. Physiol. Endocrinol. Metab. 1999, 277, 1130–1141. [Google Scholar] [CrossRef]
- Adams, S.H.; Hoppel, C.L.; Lok, K.H.; Zhao, L.; Wong, S.W.; Minkler, P.E.; Hwang, D.H.; Newman, J.W.; Garvey, W.T. Plasma acylcarnitine profiles suggest incomplete long-chain fatty acid β-oxidation and altered tricarboxylic acid cycle activity in type 2 diabetic African-American women. J. Nutr. 2009, 139, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Kramer, H.F.; Goodyear, L. Exercise, MAPK, and NF-κB signaling in skeletal muscle. J. Appl. Physiol. 2007, 103, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R. Inflammatory mechanisms in the regulation of insulin resistance. Mol. Med. 2008, 14, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.G.S.; DeLany, J.P. Increased levels of plasma acylcarnitines in obesity and type 2 diabetes and identification of a marker of glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.Y.; Kim, J.Y.; Paik, J.K.; Kim, O.Y.; Paik, Y.H.; Lee, E.J.; Lee, J.H. The association of specific metabolites of lipid metabolism with markers of oxidative stress, inflammation and arterial stiffness in men with newly diagnosed type 2 diabetes. Clin. Endocrinol. 2012, 76, 674–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, L.K.; Vance, J.E.; Vance, D.E. Phosphatidylcholine biosynthesis and lipoprotein metabolism. Biochim. Biophys. Acta 2012, 1821, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yan, L.; Chen, W.; Lin, L.; Song, X.; Yan, X.; Hang, W.; Huang, B. Metabonomics research of diabetic nephropathy and type 2 diabetes mellitusbased on UPLC–oa TOF-MS system. Anal. Chim. Acta 2009, 650, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhu, Y.; Hyun, N.; Zeng, D.; Uppal, K.; Tran, V.T.; Yu, T.; Jones, D.; He, J.; Lee, E.T.; et al. Novel metabolic markersfor the risk of diabetes development in American Indians. Diabetes Care 2015, 38, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Pamungkas, A.D.; Medriano, C.A.D.; Park, J.; Hong, S.; Jee, S.H.; Park, Y.H. High-resolution metabolomics determines the mode of onset of type 2 diabetes in a 3-year prospective cohort study. Int. J. Mol. Med. 2018, 41, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Messana, I.; Forni, F.; Ferrari, F.; Rossi, C.; Giardina, B.; Zuppi, C. Proton nuclear magnetic resonance spectral profiles of urine in type II diabetic patients. Clin. Chem. 1998, 44, 1529–1534. [Google Scholar] [PubMed]
- Yamanouchi, T.; Akanuma, H.; Asano, T.; Konishi, C.; Akaoka, I.; Akanuma, Y. Reduction and recovery of plasma 1,5-anhydro-d-glucitol level in diabetes mellitus. Diabetes 1987, 36, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, T.; Tachibana, Y.; Akanuma, H.; Minoda, S.; Shinohara, T.; Moromizato, H.; Miyashita, H.; Akaoka, I. Origin and disposal of 1,5-anhydroglucitol, a major polyol in the human body. Am. J. Physiol. Renal. Physiol. 1992, 263, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Dungan, K.M.; Buse, J.B.; Largay, J.; Kelly, M.M.; Button, E.A.; Kato, S.; Wittlin, S. 1,5-anhydroglucitol and postprandial hyperglycemia as measured by continuous glucose monitoring system in moderately controlled patients with diabetes. Diabetes Care 2006, 29, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, T.; Inoue, T.; Ogata, E.; Kashiwabara, A.; Ogata, N.; Sekino, N.; Yoshimura, T.; Ichiyanagi, K.; Kawasaki, T. Post-load glucose measurements in oral glucose tolerance tests correlate well with 1,5-anhydroglucitol, an indicator of overall glycaemic state, in subjects with impaired glucose tolerance. Clin. Sci. 2001, 101, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, T.; Ogata, N.; Tagaya, T.; Kawasaki, T.; Sekino, N.; Funato, H.; Akaoka, I.; Miyashita, H. Clinical usefulness of serum 1,5-anhydroglucitol in monitoring glycaemic control. Lancet 1996, 347, 1514–1518. [Google Scholar] [CrossRef]
- Hanefeld, M.; Fischer, S.; Julius, U.; Schulze, J.; Schwanebeck, U.; Schmechel, H.; Ziegelasch, H.J.; Lindner, J. Risk factors for myocardial infarction and death in newly detected NIDDM: The diabetes Intervention Study, 11-year follow-up. Diabetologia 1996, 39, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Muggeo, M.; Zoppini, G.; Bonora, E.; Brun, E.; Bonadonna, R.C.; Moghetti, P.; Verlato, G. Fasting plasma glucose variability predicts 10-year survival of type 2 diabetic patients: The verona diabetes study. Diabetes Care 2000, 23, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Temelkovakurktschiev, T.S.; Koehler, C.; Henkel, E.; Leonhardt, W.; Fuecker, K.; Hanefeld, M. Postchallenge plasma glucose and glycemic spikes are more strongly associated with atherosclerosis than fasting glucose or HbA1c level. Diabetes Care 2000, 23, 1830–1834. [Google Scholar] [CrossRef] [Green Version]
- Erlinger, T.; Brancati, F. Postchallenge hyperglycemia in a national sample of U.S. adults with type 2 diabetes. Diabetes Care 2001, 24, 1734–1738. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Xie, G.; Jia, W.; Jia, W. Metabolomics in human type 2 diabetes research. Front. Med. 2013, 7, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Xu, M. Application and progress of metabonomics in the study of type 2 diabetes. J. Int. Med. 2016, 191–194. [Google Scholar]
- Pallares-Méndez, R.; Aguilar-Salinas, C.A.; Cruz-Bautista, I.; del Bosque-Plata, L. Metabolomics in diabetes, a review. Ann. Med. 2016, 48, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Xiao, F.; Guo, Y.; Deng, J.; Liu, B.; Zhang, Q.; Li, K.; Wang, C.; Chen, S.; Guo, F. Hepatic phosphoserine aminotransferase 1 (PSAT1) regulates insulin sensitivity in mice via tribbles homolog 3 (TRB3). Diabetes 2015, 64, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.T.; Bos, N.A.; Harthoorn, L.F.; Stellaard, F.; Beijer-Liefers, S.; Rozing, J.; van Tol, E.A.F. Potential mechanisms explaining why hydrolyzed casein-based diets outclass single amino acid-based diets in the prevention of autoimmune diabetes in diabetes-prone BB rats. Diabetes 2012, 28, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, F. Chain amino acids and metabolic homeostasis regulation. Chin. Sci. Bull. 2013, 14, 1367. [Google Scholar]
- Malloy, V.L.; Krajcik, R.A.; Bailey, S.J.; Hristopoulos, G.; Plummer, J.D.; Orentreich, N. Methionine restriction decreases visceral fat mass and preserves insulin action in aging male Fischer 344 rats independent of energy restriction. Aging Cell 2006, 5, 305–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | NMR | MS |
|---|---|---|
| Objectivity | yes | yes |
| Repeatability | fine | fine |
| Sensibility | general | selective sensitivity |
| Resolution | general | general |
| Flux | high | high |
| Sample preparation | little | a large number |
| Molecular information | abundant | general |
| Labor intensity | low | general |
| Recurrent costs | low | high |
| Single sample cost | low | general |
| Pathway | Metabolite | Change of Direction (vs. Healthy Control) | Sample | Platform | Reference |
|---|---|---|---|---|---|
| Amino acid | Leucine | Up | Serum | NMR,UPLC-MS,GC-MS | [68] |
| metabolism | Up | Plasma | UPLC-MS | [76] | |
| Up | Plasma | GC-MS | [54] | ||
| Isoleucine | Up | Serum | NMR,UPLC-MS,GC-MS | [68] | |
| Valine | Up | Serum | NMR,UPLC-MS,GC-MS | [68] | |
| Up | Serum | GC-MS | [45] | ||
| Glutamine | Up | Urine | NMR | [43] | |
| Up | Serum | GC-MS | [59] | ||
| Glutamique | Down | Serum | NMR | [44] | |
| lysine | Down | Plasma | GC-MS | [54] | |
| Down | Serum | NMR | [44] | ||
| Down | Serum | GC-MS | [45] | ||
| Glysine | Down | Plasma | GC-MS | [54] | |
| Down | Serum | LC-MS | [49] | ||
| Down | Serum | LC-MS | [48] | ||
| Down | Plasma | LC-MS/MS | [47] | ||
| 2-Hydroxybutyrate | Up | Plasma | GC*GC-MS | [56] | |
| Up | Plasma | GC-MS | [54] | ||
| Up | Serum | GC-MS | [59] | ||
| Up | Plasma | GC-MS | [54] | ||
| Serine | Down | Plasma | UPLC-MS | [76] | |
| Tyrosine | Down | Serum | NMR | [44] | |
| Phenylalanine | Up | Plasma | UPLC-MS | [76] | |
| Up | Plasma | LC-MS/MS | [47] | ||
| Up | Serum | GC-MS | [59] | ||
| Up | Serum | LC-MS | [48] | ||
| Tryptophan | Down | Urine | NMR | [43] | |
| Alanine | Down | Serum | NMR | [44] | |
| Methionine | Up | Serum | GC-MS | [59] | |
| Histidine | Down | Serum | NMR | [44] | |
| Down | Urine | NMR | [43] | ||
| Hippurate | Up | Urine | NMR | [81] | |
| Taurine | Up | Urine | NMR | [43] | |
| Up | Plasma | LC-MS/MS | [47] | ||
| Lipid | 3-Hydroxybutyrate | Up | Plasma | GC-MS | [54] |
| metabolism | Up | Serum | NMR,UPLC-MS,GC-MS | [68] | |
| Up | Urine | NMR | [43] | ||
| Up | Serum | GC-MS | [59] | ||
| Acetoacetate | Up | Urine | NMR | [43] | |
| Fatty acids | Up | Plasma | GC-MS | [54] | |
| Up | Serum | GC-MS | [59] | ||
| Lyso PCs | Up | Plasma | UPLC-MS | [76] | |
| Up | Plasma | UPLC-MS | [76] | ||
| Lyso PC (18:2) | Down | Serum | LC-MS | [49] | |
| Down | Serum | LC-MS | [48] | ||
| Lyso PEs | Up/Down | Plasma | UPLC-MS | [76] | |
| Up/Down | Plasma | UPLC-MS | [76] | ||
| PCs Acetylcarnitines | Up/Down | Serum | LC-MS | [48] | |
| Up | Plasma | UPLC-MS | [76] | ||
| Up | Plasma | UPLC-MS | [75] | ||
| Up | Plasma | UPLC-MS | [72] | ||
| Up | Plasma | UPLC-MS | [76] | ||
| Palmitic acid | Up | Plasma | GC-MS | [66] | |
| Up | Plasma | GCxGC-TOFMS | [56] | ||
| Up | Serum | GC-MS | [67] | ||
| linolenic acid | Up | Plasma | GCxGC-TOFMS | [56] | |
| Dihydrosphingosine | Down | Serum | UPLC-oaTOF | [78] | |
| Phytosphingosine | Down | Serum | UPLC-oaTOF | [78] | |
| cholesterol | Up | Serum | LC-MS | [80] | |
| Carbohydrate | citric acid | Up | Urine | NMR | [81] |
| metabolism | Up | Urine | NMR | [43] | |
| 1,5-Anhydrogluticol | Down | Serum | NMR,UPLC-MS,GC-MS | [68] | |
| Down | Serum | GC-MS | [59] | ||
| Pyruvate | Down | Serum | GC-MS | [59] | |
| Lactate | Up | Serum | GC-MS | [59] | |
| Malate | Down | Urine | NMR | [43] | |
| Succinate | Down | Urine | NMR | [43] | |
| Fumarate | Down | Urine | NMR | [43] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Li, Y.; Wang, M.; Tang, Z.; Wang, T.; Liu, C.; Wang, C.; Zhao, B. Progress in Metabonomics of Type 2 Diabetes Mellitus. Molecules 2018, 23, 1834. https://doi.org/10.3390/molecules23071834
Ma Q, Li Y, Wang M, Tang Z, Wang T, Liu C, Wang C, Zhao B. Progress in Metabonomics of Type 2 Diabetes Mellitus. Molecules. 2018; 23(7):1834. https://doi.org/10.3390/molecules23071834
Chicago/Turabian StyleMa, Quantao, Yaqi Li, Min Wang, Ziyan Tang, Ting Wang, Chenyue Liu, Chunguo Wang, and Baosheng Zhao. 2018. "Progress in Metabonomics of Type 2 Diabetes Mellitus" Molecules 23, no. 7: 1834. https://doi.org/10.3390/molecules23071834
APA StyleMa, Q., Li, Y., Wang, M., Tang, Z., Wang, T., Liu, C., Wang, C., & Zhao, B. (2018). Progress in Metabonomics of Type 2 Diabetes Mellitus. Molecules, 23(7), 1834. https://doi.org/10.3390/molecules23071834