A Simple Method for On-Gel Detection of Myrosinase Activity

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
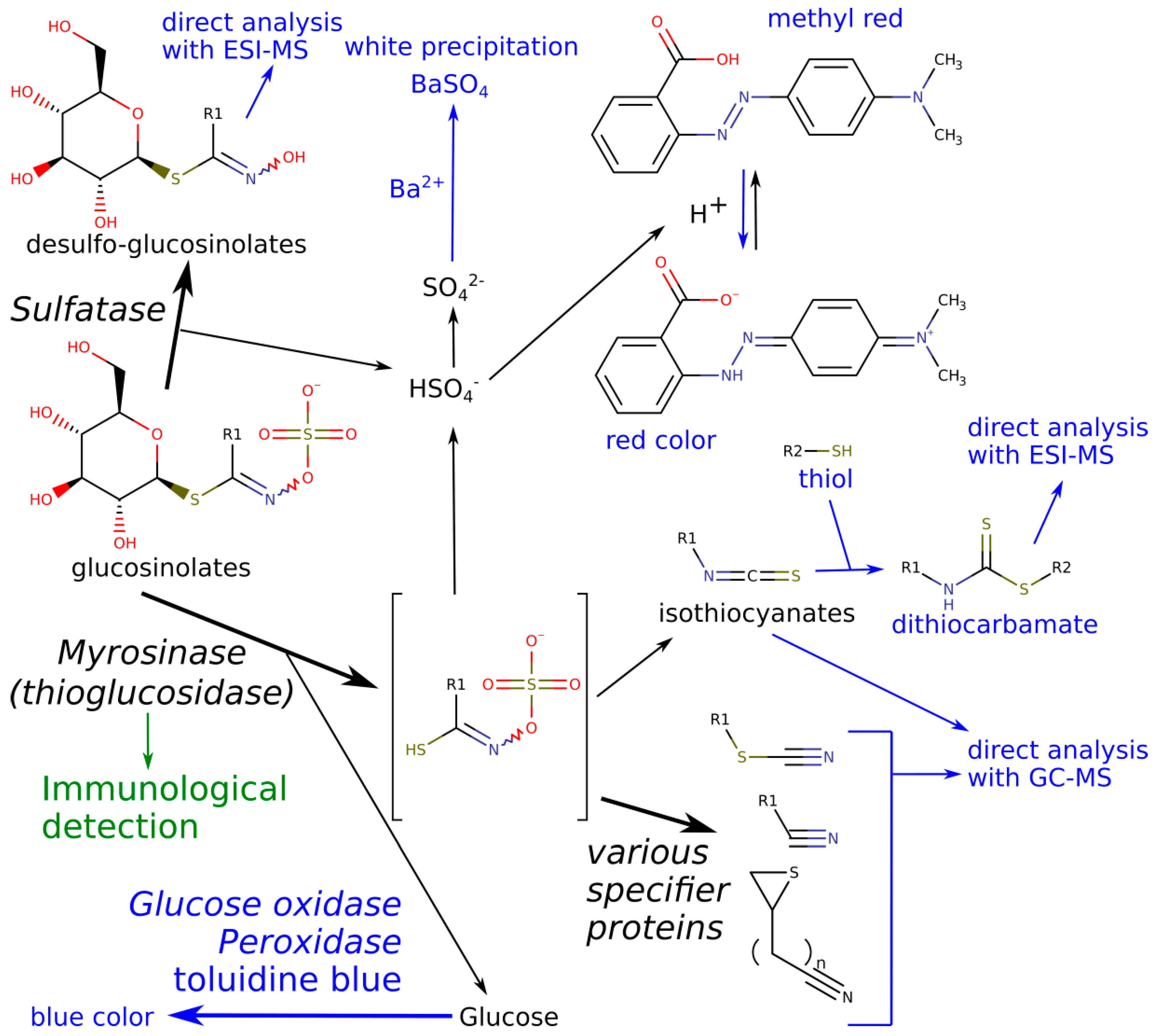
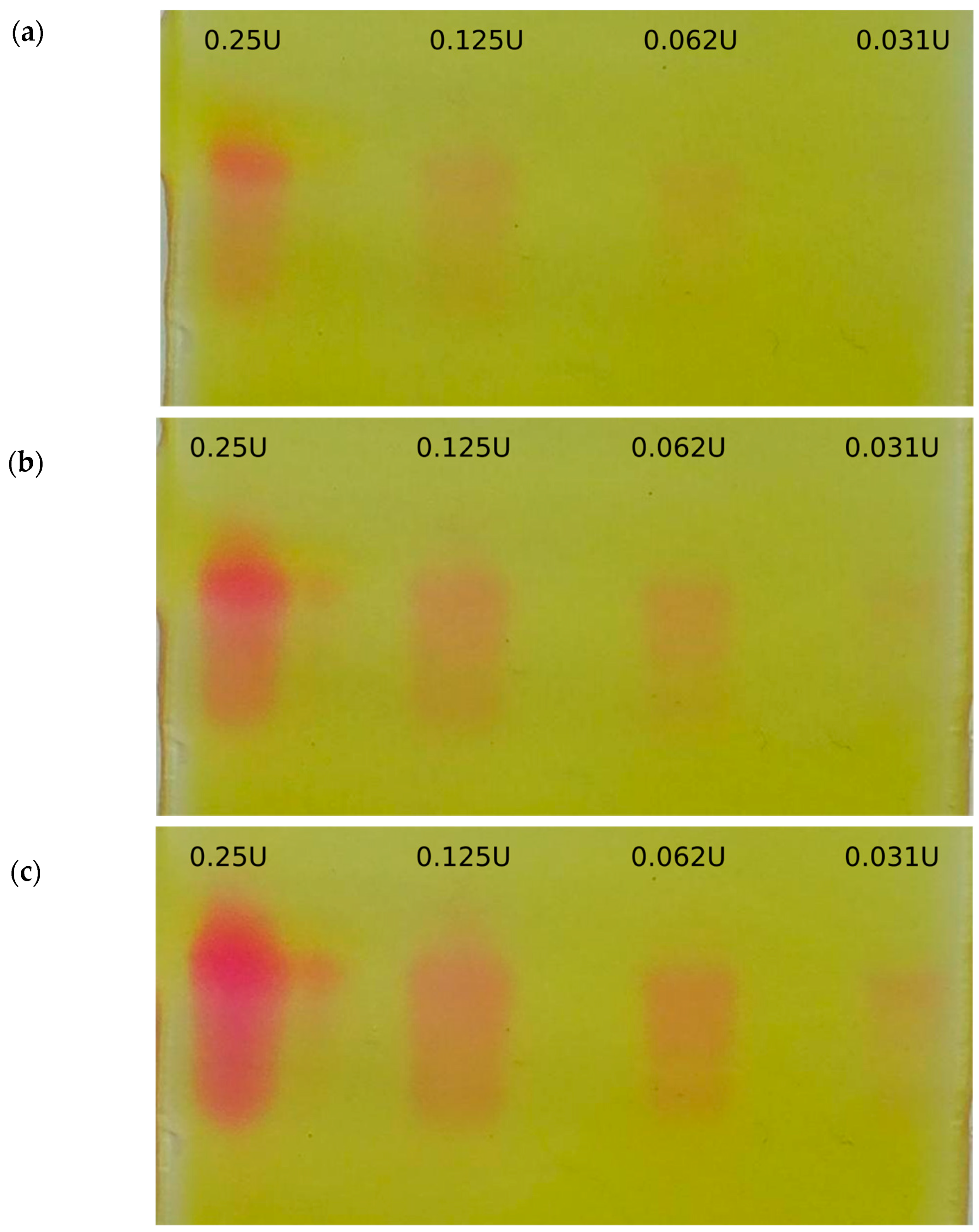
2.1. On-Gel Detection of Glucosinolate Decomposition
2.2. LC-ESI-MS of Products of Separated Sulfatase and Myrosinase Enzymes
2.3. On-Gel Detection after Separation of Crude Extracts
3. Materials and Methods
3.1. Chemicals
3.2. Pure Enzymes
3.3. Organisms
3.4. Sample Preparation
3.5. Testing of Detection Reagents
3.6. Gel-Electrophoresis
3.7. On-Gel Detection
3.8. LC-ESI-MS
3.9. Enzyme Reaction for Distinguishment of Myrosinase and Sulfatase
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kissen, R.; Rossiter, J.T.; Bones, A.M. The ‘mustard oil bomb’: Not so easy to assemble?! Localization, expression and distribution of the components of the myrosinase enzyme system. Phytochem. Rev. 2009, 8, 69–86. [Google Scholar] [CrossRef]
- Nguyen, N.M.; Gonda, S.; Vasas, G. A Review on the Phytochemical Composition and Potential Medicinal Uses of Horseradish (Armoracia rusticana) Root. Food Rev. Int. 2013, 29, 1–15. [Google Scholar] [CrossRef]
- Ludikhuyze, L.; Rodrigo, L.; Hendrickx, M. The Activity of Myrosinase from Broccoli (Brassica oleracea L. cv. Italica): Influence of Intrinsic and Extrinsic Factors. J. Food Prot. 2000, 63, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, R.; Finiguerra, M.G.; Rossi, A.A.; Palmieri, S. Isolation and Biochemical Characterization of a Basic Myrosinase from Ripe Crambe abyssinica Seeds, Highly Specific for epi-Progoitrin. J. Agric. Food Chem. 2003, 51, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Smits, J.P.; Knol, W.; Bol, J. Glucosinolate degradation by Aspergillus clavatus and Fusarium oxysporum in liquid and solid-state fermentation. Appl. Microbiol. Biotechnol. 1993, 38, 696–701. [Google Scholar] [CrossRef]
- Albaser, A.; Kazana, E.; Bennett, M.H.; Cebeci, F.; Luang-In, V.; Spanu, P.D.; Rossiter, J.T. Discovery of a Bacterial Glycoside Hydrolase Family 3 (GH3) β-Glucosidase with Myrosinase Activity from a Citrobacter Strain Isolated from Soil. J. Agric. Food Chem. 2016, 64, 1520–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tani, N.; Ohtsuru, M.; Hata, T. Purification and general characteristics of bacterial myrosinase produced by Enterobacter cloacae. Agric. Biol. Chem. 1974, 38, 1623–1630. [Google Scholar] [CrossRef]
- Szűcs, Z.; Plaszkó, T.; Cziáky, Z.; Kiss-Szikszai, A.; Emri, T.; Bertóti, R.; Sinka, L.T.; Vasas, G.; Gonda, S. Endophytic fungi from the roots of horseradish (Armoracia rusticana) and their interactions with the defensive metabolites of the glucosinolate-myrosinase-isothiocyanate system. BMC Plant Biol. 2018, 18, 85. [Google Scholar] [CrossRef] [PubMed]
- Beran, F.; Pauchet, Y.; Kunert, G.; Reichelt, M.; Wielsch, N.; Vogel, H.; Reinecke, A.; Svatoš, A.; Mewis, I.; Schmid, D.; et al. Phyllotreta striolata flea beetles use host plant defense compounds to create their own glucosinolate-myrosinase system. Proc. Natl. Acad. Sci. USA 2014, 111, 7349–7354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.E.; Bridges, M.; Bones, A.M.; Cole, R.; Rossiter, J.T. Purification and characterisation of a non-plant myrosinase from the cabbage aphid Brevicoryne brassicae (L.). Insect Biochem. Mol. Biol. 2001, 31, 1–5. [Google Scholar] [CrossRef]
- Pontoppidan, B.; Ekbom, B.; Eriksson, S.; Meijer, J. Purification and characterization of myrosinase from the cabbage aphid (Brevicoryne brassicae), a brassica herbivore. Eur. J. Biochem. 2001, 268, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Jander, G. Arabidopsis myrosinases TGG1 and TGG2 have redundant function in glucosinolate breakdown and insect defense. Plant J. 2006, 46, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Kushad, M.M. Purification and characterization of myrosinase from horseradish (Armoracia rusticana) roots. Plant Physiol. Biochem. 2005, 43, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Ek, B.; Xue, J.; Rask, L.; Meijer, J. Identification and characterization of soluble and insoluble myrosinase isoenzymes in different organs of Sinapis alba. Physiol. Plant. 2001, 111, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Lenman, M.; Falk, A.; Rodin, J.; Hoglund, A.S.; Ek, B.; Rask, L. Differential Expression of Myrosinase Gene Families. Plant Physiol. 1993, 103, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Loebers, A.; Müller-Uri, F.; Kreis, W. A young root-specific gene (ArMY2) from horseradish encoding a MYR II myrosinase with kinetic preference for the root-specific glucosinolate gluconasturtiin. Phytochemistry 2014, 99, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Durham, P.L.; Poulton, J.E. Enzymic Properties of Purified Myrosinase from Lepidium sativum Seedlings. Z. Naturforsch. C 2014, 45, 173–178. [Google Scholar] [CrossRef]
- Palmieri, S.; Iori, R.; Leoni, O. Myrosinase from Sinapis alba L.: A new method of purification for glucosinolate analyses. J. Agric. Food Chem. 1986, 34, 138–140. [Google Scholar] [CrossRef]
- Francis, F.; Lognay, G.; Wathelet, J.-P.; Haubruge, E. Characterisation of aphid myrosinase and degradation studies of glucosinolates. Arch. Insect Biochem. Physiol. 2002, 50, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Lambrix, V.; Reichelt, M.; Mitchell-Olds, T.; Kliebenstein, D.J.; Gershenzon, J. The Arabidopsis Epithiospecifier Protein Promotes the Hydrolysis of Glucosinolates to Nitriles and Influences Trichoplusia ni Herbivory. Plant Cell 2001, 13, 2793–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittstock, U.; Agerbirk, N.; Stauber, E.J.; Olsen, C.E.; Hippler, M.; Mitchell-Olds, T.; Gershenzon, J.; Vogel, H. Successful herbivore attack due to metabolic diversion of a plant chemical defense. Proc. Natl. Acad. Sci. USA 2004, 101, 4859–4864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piekarska, A.; Kusznierewicz, B.; Meller, M.; Dziedziul, K.; Namiesnik, J.; Bartoszek, A. Myrosinase activity in different plant samples; optimisation of measurement conditions for spectrophotometric and pH-stat methods. Ind. Crop. Prod. 2013, 50, 58–67. [Google Scholar] [CrossRef]
- Vastenhout, K.J.; Tornberg, R.H.; Johnson, A.L.; Amolins, M.W.; Mays, J.R. High-performance liquid chromatography-based method to evaluate kinetics of glucosinolate hydrolysis by Sinapis alba myrosinase. Anal. Biochem. 2014, 465, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Gonda, S.; Kiss-Szikszai, A.; Szűcs, Z.; Nguyen, N.M.; Vasas, G. Myrosinase Compatible Simultaneous Determination of Glucosinolates and Allyl Isothiocyanate by Capillary Electrophoresis Micellar Electrokinetic Chromatography (CE-MEKC). Phytochem. Anal. 2016, 27, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, A.; Rhodes, M.; Fenwick, G. Determination of Myrosinase (thioglucoside Glucohydrolase) Activity by a Spectrophotometric Coupled Enzyme Assay. Anal. Biochem. 1984, 139, 284–291. [Google Scholar] [CrossRef]
- Bones, A.M.; Thangstad, O.P.; Haugen, O.A.; Espevik, T. Fate of Myrosin Cells: Characterization of Monoclonal Antibodies Against Myrosinase. J. Exp. Bot. 1991, 42, 1541–1550. [Google Scholar] [CrossRef]
- Andréasson, E.; Jørgensen, L.B.; Höglund, A.-S.; Rask, L.; Meijer, J. Different Myrosinase and Idioblast Distribution in Arabidopsis and Brassica napus. Plant Physiol. 2001, 127, 1750–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenman, M.; Rödin, J.; Josefsson, L.-G.; Rask, L. Immunological characterization of rapeseed myrosinase. Eur. J. Biochem. 1990, 194, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGibbon, D.B.; Allison, R.M. A method for the separation and detection of plant glucosinolases (myrosinases). Phytochemistry 1970, 9, 541–544. [Google Scholar] [CrossRef]
- Bones, A.; Slupphaug, G. Purification, Characterization and Partial Amino Acid Sequencing of β-thioglucosidase from Brassica napus L. J. Plant Physiol. 1989, 134, 722–729. [Google Scholar] [CrossRef]
- Shikita, M.; Fahey, J.W.; Golden, T.R.; Holtzclaw, W.D.; Talalay, P. An unusual case of “uncompetitive activation” by ascorbic acid: Purification and kinetic properties of a myrosinase from Raphanus sativus seedlings. Biochem. J. 1999, 341, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Rakariyatham, N.; Butrindr, B.; Niamsup, H.; Shank, L. Screening of filamentous fungi for production of myrosinase. Braz. J. Microbiol. 2005, 36, 242–245. [Google Scholar] [CrossRef]
- Sharma, A.; Yogavel, M.; Sharma, A. Utility of anion and cation combinations for phasing of protein structures. J. Struct. Funct. Gen. 2012, 13, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.G.; Gordon, E.; Robinson, D. The identification of myrosinase after the electrophoresis of Brassica and Sinapis seed proteins. Phytochemistry 1968, 7, 1345–1348. [Google Scholar] [CrossRef]
- Wang, H.; Lin, W.; Shen, G.; Khor, T.-O.; Nomeir, A.A.; Kong, A.-N. Development and Validation of an LC-MS-MS Method for the Simultaneous Determination of Sulforaphane and its Metabolites in Rat Plasma and its Application in Pharmacokinetic Studies. J. Chromatogr. Sci. 2011, 49, 801–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platz, S.; Kuehn, C.; Schiess, S.; Schreiner, M.; Mewis, I.; Kemper, M.; Pfeiffer, A.; Rohn, S. Determination of benzyl isothiocyanate metabolites in human plasma and urine by LC-ESI-MS/MS after ingestion of nasturtium (Tropaeolum majus L.). Anal. Bioanal. Chem. 2013, 405, 7427–7436. [Google Scholar] [CrossRef] [PubMed]
- Kusznierewicz, B.; Iori, R.; Piekarska, A.; Namieśnik, J.; Bartoszek, A. Convenient identification of desulfoglucosinolates on the basis of mass spectra obtained during liquid chromatography–diode array–electrospray ionisation mass spectrometry analysis: Method verification for sprouts of different Brassicaceae species extracts. J. Chromatogr. A 2013, 1278, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. Sampling of wild plants complied with local legislation and guidelines. |



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonda, S.; Szűcs, Z.; Plaszkó, T.; Cziáky, Z.; Kiss-Szikszai, A.; Vasas, G.; M-Hamvas, M. A Simple Method for On-Gel Detection of Myrosinase Activity. Molecules 2018, 23, 2204. https://doi.org/10.3390/molecules23092204
Gonda S, Szűcs Z, Plaszkó T, Cziáky Z, Kiss-Szikszai A, Vasas G, M-Hamvas M. A Simple Method for On-Gel Detection of Myrosinase Activity. Molecules. 2018; 23(9):2204. https://doi.org/10.3390/molecules23092204
Chicago/Turabian StyleGonda, Sándor, Zsolt Szűcs, Tamás Plaszkó, Zoltán Cziáky, Attila Kiss-Szikszai, Gábor Vasas, and Márta M-Hamvas. 2018. "A Simple Method for On-Gel Detection of Myrosinase Activity" Molecules 23, no. 9: 2204. https://doi.org/10.3390/molecules23092204
APA StyleGonda, S., Szűcs, Z., Plaszkó, T., Cziáky, Z., Kiss-Szikszai, A., Vasas, G., & M-Hamvas, M. (2018). A Simple Method for On-Gel Detection of Myrosinase Activity. Molecules, 23(9), 2204. https://doi.org/10.3390/molecules23092204