Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
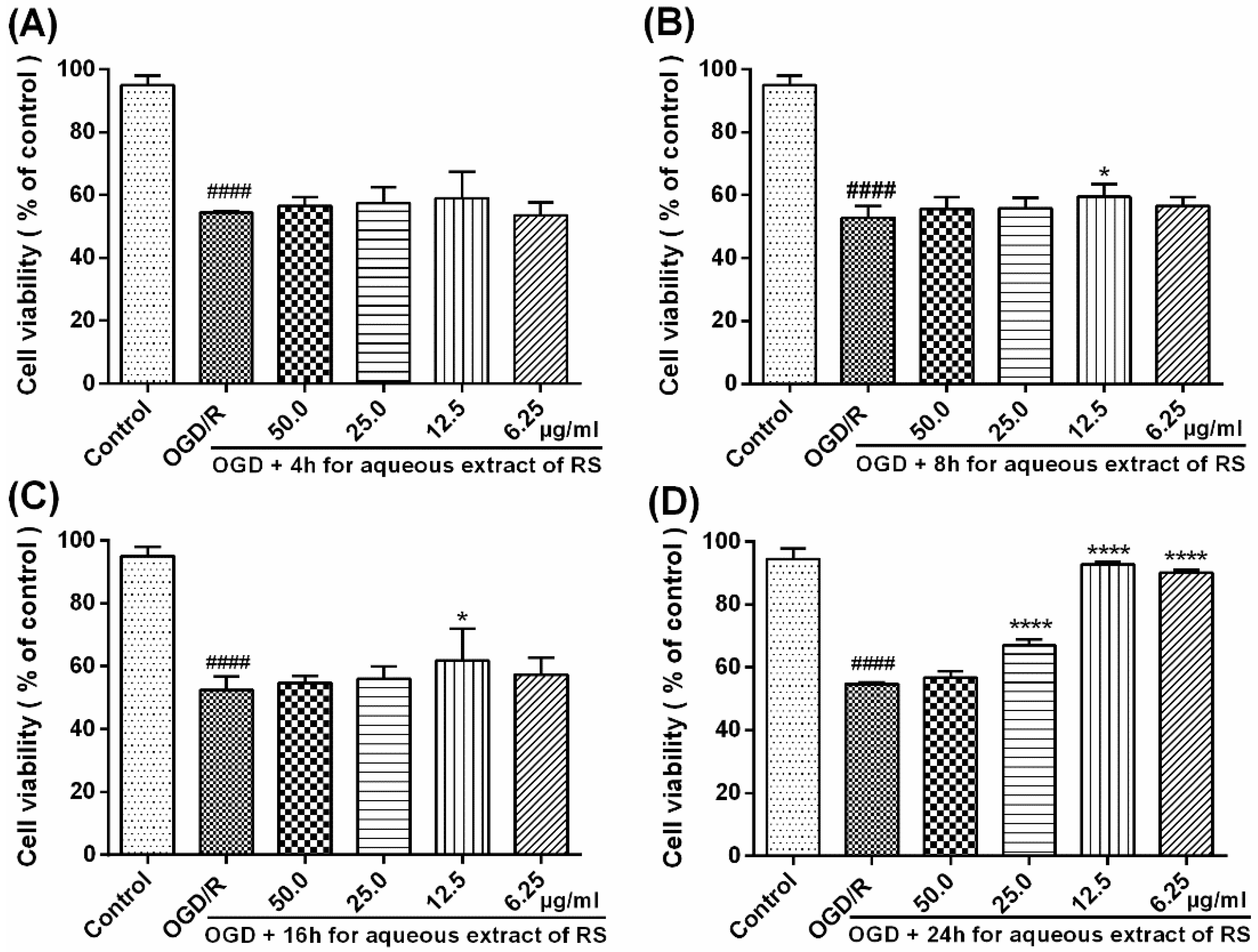
2.1. Cell Viability in PC12 Cells after OGD/R
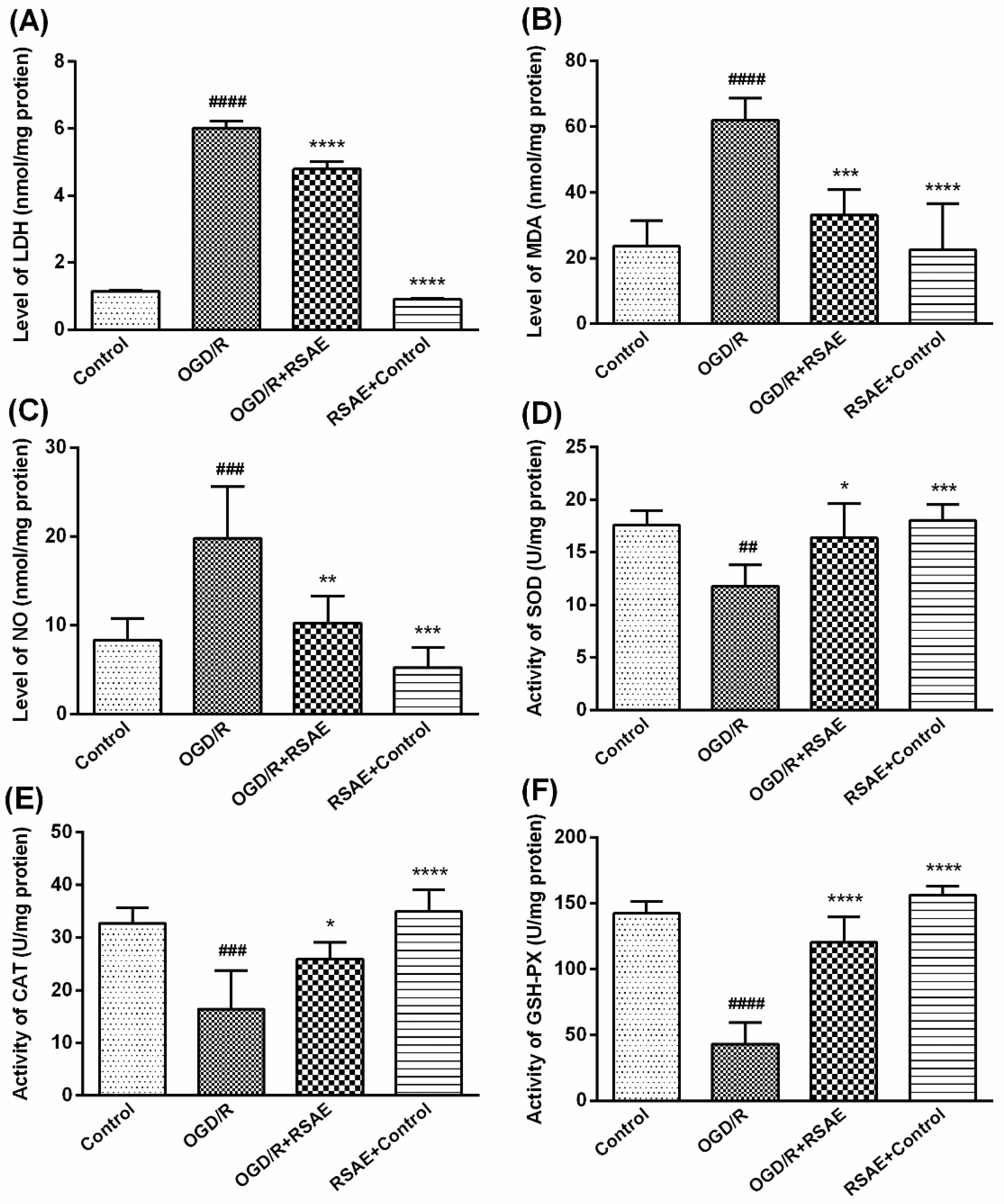
2.2. Levels of LDH Leakage, MDA, and Oxidative Stress in PC12 Cells after OGD/R
2.3. Mitochondrial Membrane Potential in PC12 Cells after OGD/R
2.4. Infarct Volume, Brain Water Content, MDA, NO Concentrations, and SOD Activity
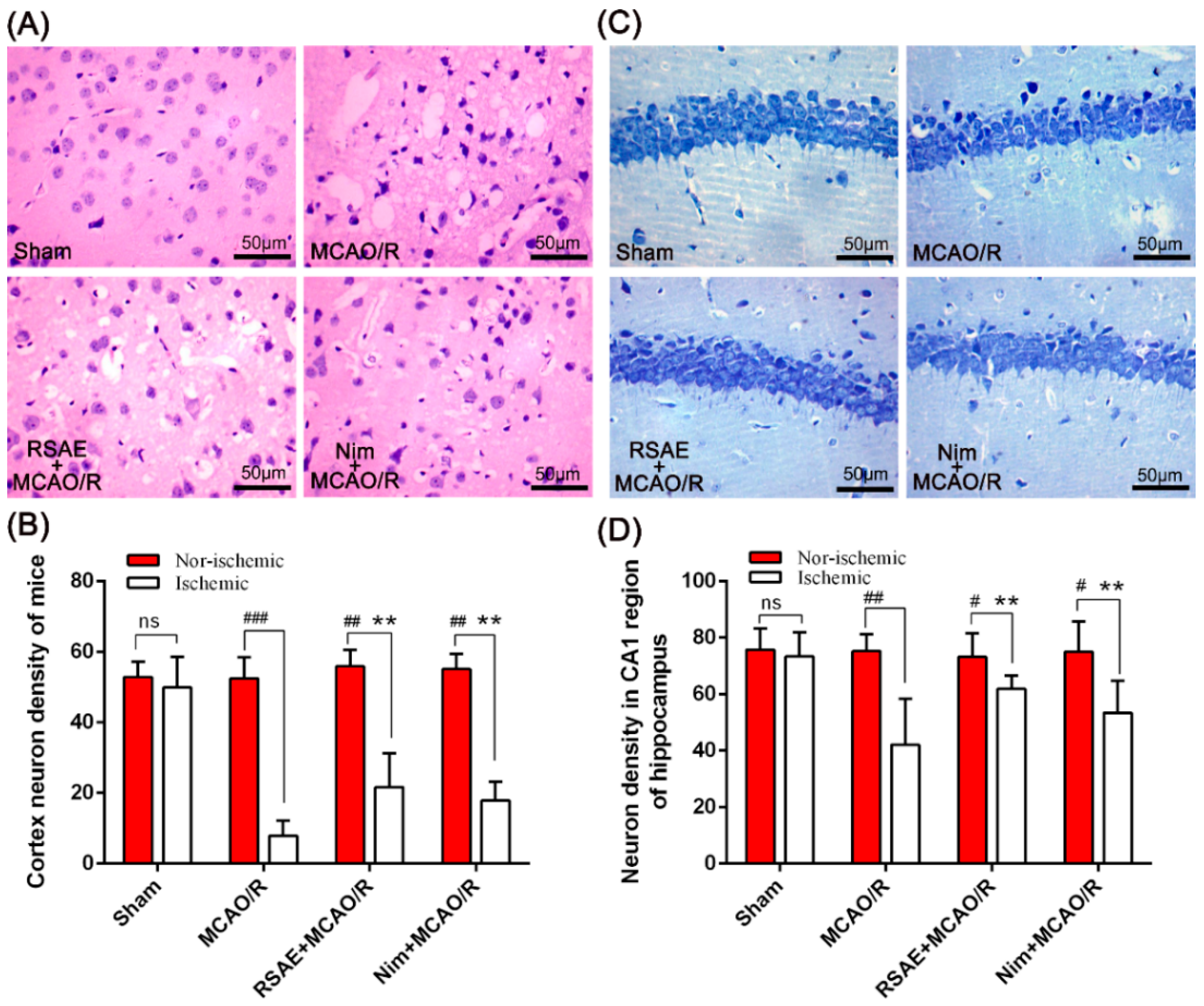
2.5. Representative Images of HE Staining and Nissl Staining
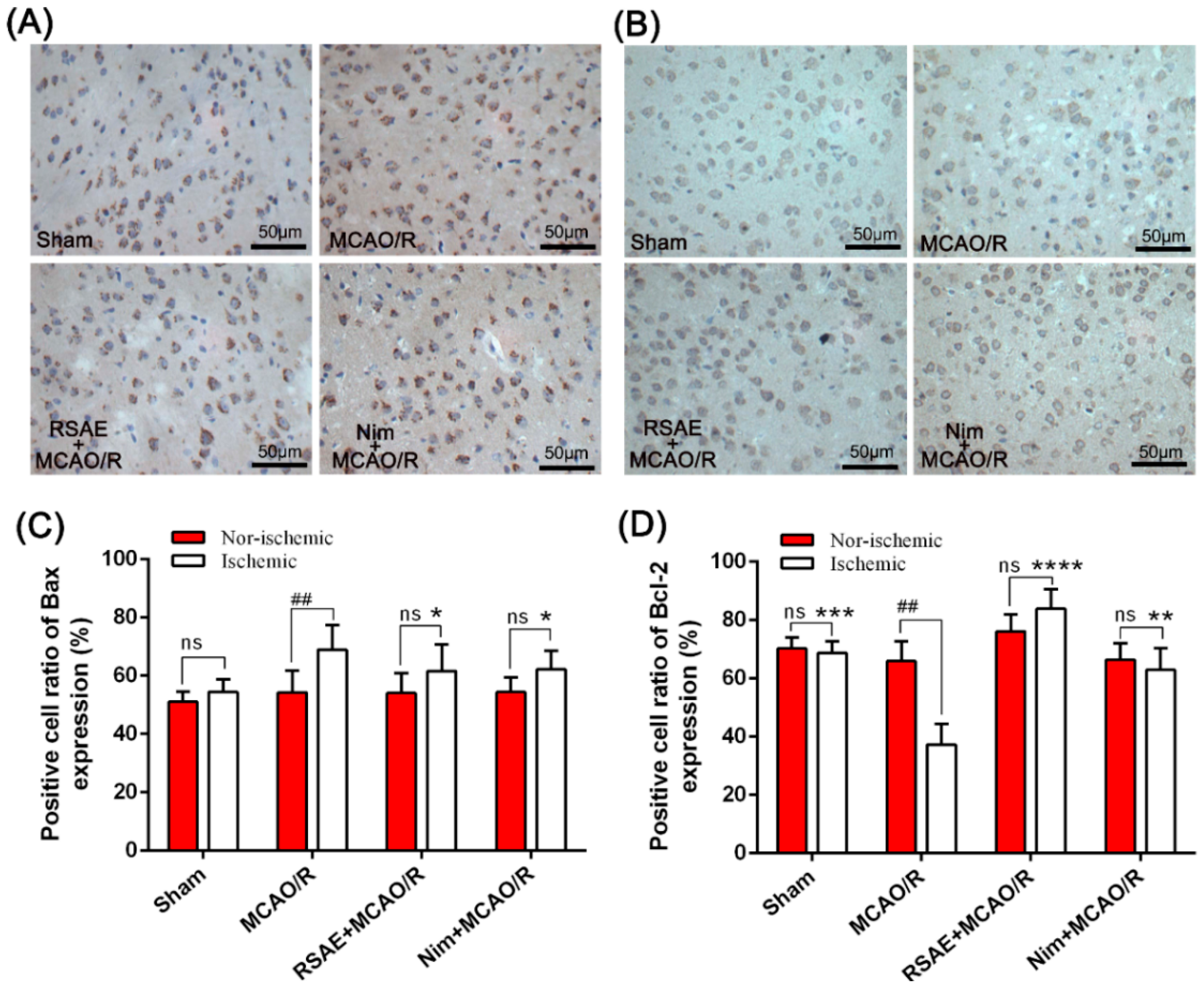
2.6. Expression of Bax and Bcl-2 in Mice with Focal Cerebral Ischemia
2.7. ERK1/2, JNK1/2 and p38 MAPK in Mice after MCAO/R
3. Discussion
4. Methods
4.1. RSAE Preparation
4.2. Cell Cultures and OGD/R Models
4.3. Cell Viability Analysis
4.4. Assessment of Cell Injury or Death
4.5. Determination of the Levels of Oxidative Stress
4.6. Measurement of the Mitochondrial Membrane Potential
4.7. Mouse MCAO/R Model and Drug Administration
4.8. Quantification of Infarct Volume and Brain Water Content
4.9. Assessment of the Levels of Superoxide Dismutase Activity and Malondialdehyde and Nitric Oxide Contents
4.10. Haematoxylin-Eosin and Nissl Staining
4.11. Immunohistochemical Evaluation
4.12. Western Blot Analysis
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wardlaw, J.M.; Zoppo, G.; Yamaguchi, T.; Berge, E. Thrombolysis for acute ischemic stroke. Cochrane Database Syst. Rev. 2003, 3. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion—From mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef]
- Turley, K.R.; Toledo-Pereyra, L.H.; Kothari, R.U. Molecular mechanisms in the pathogenesis and treatment of acute ischemic stroke. J. Investig. Surg. 2005, 18, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Bereczki, D. Pharmacotherapy of stroke. Neuropsychopharmacol Hung. 2009, 11, 7–13. [Google Scholar] [PubMed]
- Ischemic stroke: Acute-phase drug therapy. Mostly aspirin and heparin. Prescrire Int. 2005, 14, 146–152.
- Del Zoppo, G.J.; Saver, J.L.; Jauch, E.C.; Adams, H.P. Expansion of the time window for treatment of acute ischemic stroke with intravenous tissue plasminogen activator: A science advisory from the American Heart Association/American Stroke Association. Stroke 2009, 40, 2945–2948. [Google Scholar] [CrossRef] [PubMed]
- Sandercock, P.; Wardlaw, J.M.; Lindley, R.I.; Dennis, M.; Cohen, G.; Murray, G.; Innes, K.; Venables, G.; Czlonkowska, A.; Kobayashi, A. The benefits and harms of intravenous thrombolysis with recombinant tissue plasminogen activator within 6 h of acute ischemic stroke (the third international stroke trial [IST-3]): A randomised controlled trial. Lancet 2012, 379, 2352–2363. [Google Scholar] [PubMed]
- Chen, C.H.; Chen, S.J.; Su, C.C.; Yen, C.C.; Tseng, T.J.; Jinn, T.R.; Tang, F.C.; Chen, K.L.; Su, Y.C.; Lee, K.I.; et al. Chloroacetic acid induced neuronal cells death through oxidative stress-mediated p38-MAPK activation pathway regulated mitochondria-dependent apoptotic signals. Toxicology 2013, 303, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Y.; Liu, F.; Wu, F.J.; Li, J.L. Effects of ginsenoside Rg1 on the expressions of p-eRK1/2 and p-JNK in local cerebral ischemia/reperfusion injury rats. China J. Chin. Mater. Med. 2013, 33, 229–334. [Google Scholar]
- Lu, T.H.; Tseng, T.J.; Su, C.C.; Tang, F.C.; Yen, C.C.; Liu, Y.Y.; Yang, C.Y.; Wu, C.C.; Chen, K.L.; Hung, D.Z. Arsenic induces reactive oxygen species-caused neuronal cell apoptosis through JNK/ERK-mediated mitochondria-dependent and GRP 78/CHOP-regulated pathways. Toxicol. Lett. 2014, 224, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.H.; Hsieh, S.Y.; Yen, C.C.; Wu, H.C.; Chen, K.L.; Hung, D.Z.; Chen, C.H.; Wu, C.C.; Su, Y.C.; Chen, Y.W. Involvement of oxidative stress-mediated ERK1/2 and p38 activation regulated mitochondria-dependent apoptotic signals in methylmercury-induced neuronal cell injury. Toxicol. Lett. 2011, 204, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Wang, N.; Fan, X.; Zhang, L.; Luo, Y.; Huang, R.; Zhang, L.; Li, Y.; Zhao, G.; Li, L. Protective effects of cornel iridoid glycoside in rats after traumatic brain injury. Neurochem. Res. 2018, 43, 959–971. [Google Scholar] [CrossRef]
- Pietri, M.; Caprini, A.; Mouillet-Richard, S.; Pradines, E.; Ermonval, M.; Grassi, J.; Kellermann, O.; Schneider, B. Overstimulation of PrPC signaling pathways by prion peptide 106-126 causes oxidative injury of bioaminergic neuronal cells. J. Biol. Chem. 2006, 281, 28470–28479. [Google Scholar] [CrossRef] [PubMed]
- Khoshnam, S.E.; Winlow, W.; Farzaneh, M.; Farbood, Y.; Moghaddam, H.F. Pathogenic mechanisms following ischemic stroke. Neurol. Sci. 2017, 38, 1167–1186. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, W.; Yang, F.; Wang, Y.; Yue, S.; Wang, T.; Wang, X. Shengmai injection reduces apoptosis and enhances angiogenesis after myocardial ischemia and reperfusion injury in rats. Biomed. Pharmacother. 2018, 104, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Sezen, S.C.; Kucuk, A.; Ozer, A.; Kilic, Y.; Mardin, B.; Alkan, M.; Erkent, F.D.; Arslan, M.; Unal, Y.; Oktar, G.L.; Tosun, M. Assessment of the effects of levosimendan and thymoquinone on lung injury after myocardial ischemia reperfusion in rats. Drug Des. Dev. Ther. 2018, 12, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Meng, Q.; Lu, D.; Liu, X.; Wang, Y.; Hao, J. Mitofusin2 induces cell autophagy of pancreatic cancer through inhibiting the PI3K/Akt/mTOR signaling pathway. Oxid. Med. Cell. Longev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nan, G. The mitogen-activated protein kinase (MAPK) signaling pathway as a discovery target in stroke. J. Mol. Neurosci. 2016, 59, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Yen, T.L.; Hsu, C.Y.; Thomas, P.A.; Sheu, J.R.; Jayakumar, T. Multi-targeting andrographolide, a novel NF-kB inhibitor, as a potential therapeutic agent for stroke. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Lebesgue, D.; Chevaleyre, V.; Zukin, R.S.; Etgen, A.M. Estradiol rescues neurons from global ischemia-induced cell death: Multiple cellular pathways of neuroprotection. Steroids 2009, 74, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.; Fan, J.; Han, J. Ameliorating effects of traditional Chinese medicine preparation, Chinese materia medica and active compounds on ischemia/reperfusion-induced cerebral microcirculatory disturbances and neuron damage. Acta Pharm. Sin. B 2015, 5, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; Xu, F.; Liu, Z.; Ma, L.M.; Shang, M.Y.; Liu, G.X.; Cai, S.Q. Identification of chemical constituents in Scrophulariae Radix by HPLC-IT-TOF-MS. China J. Chin. Mater. Med. 2016, 41, 1257–1268. [Google Scholar]
- You-Hua, C.; Jin, Q.; Jing, H.; Bo-Yang, Y. Structural characterization and identification of major constituents in radix scrophulariae by hplc coupled with electrospray ionization quadrupole time-of-flight tandem mass spectrometry. Chin. J. Nat. Med. 2014, 12, 47–54. [Google Scholar] [CrossRef]
- Shen, X.; Eichhorn, T.; Greten, H.J.; Efferth, T. Effects of Scrophularia ningpoensis Hemsl. on inhibition of proliferation, apoptosis induction and NF-kappaB signaling of immortalized and cancer cell lines. Pharmaceuticals 2012, 5, 189–208. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.C.; Hsu, C.C.; Liu, S.H.; Su, C.C.; Yen, C.C.; Lee, M.J.; Chen, K.L.; Ho, T.J.; Hung, D.Z.; Wu, C.C. Cadmium induces apoptosis in pancreatic beta-cells through a mitochondria-dependent pathway: The role of oxidative stress-mediated c-Jun N-terminal kinase activation. PLoS ONE 2013, 8, e54374. [Google Scholar]
- Lee, K.-I.; Su, C.-C.; Yang, C.-Y.; Hung, D.-Z.; Lin, C.-T.; Lu, T.-H.; Liu, S.-H.; Huang, C.-F. Etoposide induces pancreatic β-cells cytotoxicity via the jnk/erk/gsk-3 signaling-mediated mitochondria-dependent apoptosis pathway. Toxicol. Vitr. 2016, 36, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Eto, M.; Kozaki, K.; Akishita, M.; Okabe, T.; Ouchi, Y. Raloxifene analogue ly117018 suppresses oxidative stress-induced endothelial cell apoptosis through activation of erk1/2 signaling pathway. Eur. J. Pharmacol. 2008, 589, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.N.; He, Y.Y.; Wu, G.; Khan, M.; Hsu, C.Y. Effect of brain edema on infarct volume in a focal ischemia model in rats. Stroke 1993, 24, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xu, B.; Xiao, A.; Liu, L.; Fang, X.; Liu, R.; Turlova, E.; Barszczyk, A.; Zhong, X.; Sun, C.L.F.; Britto, L.R.G.; Feng, Z.-P.; Sun, H.-S. Trpm7 inhibitor carvacrol protects brain from neonatal hypoxic-ischemic injury. Mol. Brain 2015, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-L.; Qin, P.; Li, Y.; Shen, L.; Wang, S.-Q.; Dong, H.-L.; Hou, W.-G.; Xiong, L.-Z. The effects of different doses of estradiol (e2) on cerebral ischemia in an in vitro model of oxygen and glucose deprivation and reperfusion and in a rat model of middle carotid artery occlusion. BMC Neurosci. 2013, 14, 118. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Nishimura, M.; Wang, Y.; Sims, J.R.; Qiu, S.; Savitz, S.I.; Salomone, S.; Moskowitz, M. Early release of HMGB-1 from neurons after the onset of brain ischemia. J. Cereb. Blood Flow Metab. 2008, 28, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Sun, X.; Yin, L.; Han, X.; Xu, L.; Qi, Y.; Xu, Y.; Li, H.; Lin, Y.; Liu, K.; Peng, J. Dioscin ameliorates cerebral ischemia/reperfusion injury through the downregulation of tlr4 signaling via hmgb-1 inhibition. Free. Radic. Biol. Med. 2015, 84, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Procaccio, V.; Bris, C.; de la Barca, J.M.C.; Oca, F.; Chevrollier, A.; Amati-Bonneau, P.; Bonneau, D.; Reynier, P. Perspectives of drug-based neuroprotection targeting mitochondria. Rev. Neurol. 2014, 170, 390–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaux, D.L. A boom time for necrobiology. Curr. Biol. 1993, 3, 877–878. [Google Scholar] [CrossRef]
- Martinou, J.-C.; Youle, R.J. Mitochondria in apoptosis: bcl-2 family members and mitochondrial dynamics. Dev. Cell 2011, 21, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, J.; Peng, Q.; Liu, Y.; Liu, W.; Luo, C.; Peng, J.; Li, J.; Yung, K.K.L.; Mo, Z. Neuroprotective effects of bilobalide on cerebral ischemia and reperfusion injury are associated with inhibition of pro-inflammatory mediator production and down-regulation of jnk1/2 and p38 mapk activation. J. Neuroinflamm. 2014, 11. [Google Scholar] [CrossRef] [PubMed]
- Martín-Romero, F.J.; Santiago-Josefat, B.; Correa-Bordes, J.; Gutierrez-Merino, C.; Fernandez-Salguero, P. Potassium-induced apoptosis in rat cerebellar granule cells involves cell-cycle blockade at the g1/s transition. J. Mol. Neurosci. 2000, 15, 155–166. [Google Scholar] [CrossRef]
- Ma, B.; Meng, X.; Wang, J.; Sun, J.; Ren, X.; Qin, M.; Sun, J.; Sun, G.; Sun, X. Notoginsenoside r1 attenuates amyloid-β-induced damage in neurons by inhibiting reactive oxygen species and modulating mapk activation. Int. Immunopharmacol. 2014, 22, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Lu, S.; Luo, Y.; Wang, S.; Yang, K.; Zhai, Y.; Sun, G.; Sun, X. Attenuation of TNF-α-induced inflammatory injury in endothelial cells by ginsenoside Rb1 via inhibiting NF-κB, JNK and p38 signaling pathways. Front. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, M.; Wang, M.; Tian, Y.; Sun, X.; Sun, G.; Sun, X. Bavachinin induces oxidative damage in HepaRG cells through p38/JNK MAPK pathways. Toxins 2018, 10, 154. [Google Scholar] [CrossRef] [PubMed]
- Ai, Q.; Sun, G.; Luo, Y.; Dong, X.; Hu, R.; Meng, X.; Sun, X. Ginsenoside rb1 prevents hypoxia-reoxygenation-induced apoptosis in h9c2 cardiomyocytes via an estrogen receptor-dependent crosstalk among the akt, jnk, and erk 1/2 pathways using a label-free quantitative proteomics analysis. RSC Adv. 2015, 5, 26346–26363. [Google Scholar] [CrossRef]
- Wang, M.; Xu, X.; Xu, H.; Wen, F.; Zhang, X.; Sun, H.; Yao, F.; Sun, G.; Sun, X. Effect of the total saponins of aralia elata (miq) seem on cardiac contractile function and intracellular calcium cycling regulation. J. Ethnopharmacol. 2014, 155, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Feng, G. Protective effect of salvianolic acid b on isolated heart ischemia/reperfusion injury in rats. China J. Chin. Mater. Med. 2012. [Google Scholar] [CrossRef]
- Di, Y.; He, Y.L.; Zhao, T.; Huang, X.; Wu, K.W.; Liu, S.H.; Zhao, Y.Q.; Fan, M.; Wu, L.Y.; Zhu, L.L. Methylene blue reduces acute cerebral ischemic injury via the induction of mitophagy. Mol. Med. 2015, 21, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Liang, J.; Wang, J.; Kolattukudy, P.E. Mcp-induced protein 1 mediates the minocycline-induced neuroprotection against cerebral ischemia/reperfusion injury in vitro and in vivo. J. Neuroinflamm. 2015, 12, 39. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, X.; Xie, W.; Xu, Q.; Liang, T.; Xu, X.; Sun, G.; Sun, X. Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways. Molecules 2018, 23, 2401. https://doi.org/10.3390/molecules23092401
Meng X, Xie W, Xu Q, Liang T, Xu X, Sun G, Sun X. Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways. Molecules. 2018; 23(9):2401. https://doi.org/10.3390/molecules23092401
Chicago/Turabian StyleMeng, Xiangbao, Weijie Xie, Quanfu Xu, Tian Liang, Xudong Xu, Guibo Sun, and Xiaobo Sun. 2018. "Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways" Molecules 23, no. 9: 2401. https://doi.org/10.3390/molecules23092401
APA StyleMeng, X., Xie, W., Xu, Q., Liang, T., Xu, X., Sun, G., & Sun, X. (2018). Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways. Molecules, 23(9), 2401. https://doi.org/10.3390/molecules23092401