IL-6 in Osteoarthritis: Effects of Pine Stilbenoids

Abstract
:1. Introduction
2. Results
2.1. IL-6 Concentrations in Synovial Fluid are Higher than those in Plasma in Patients with OA
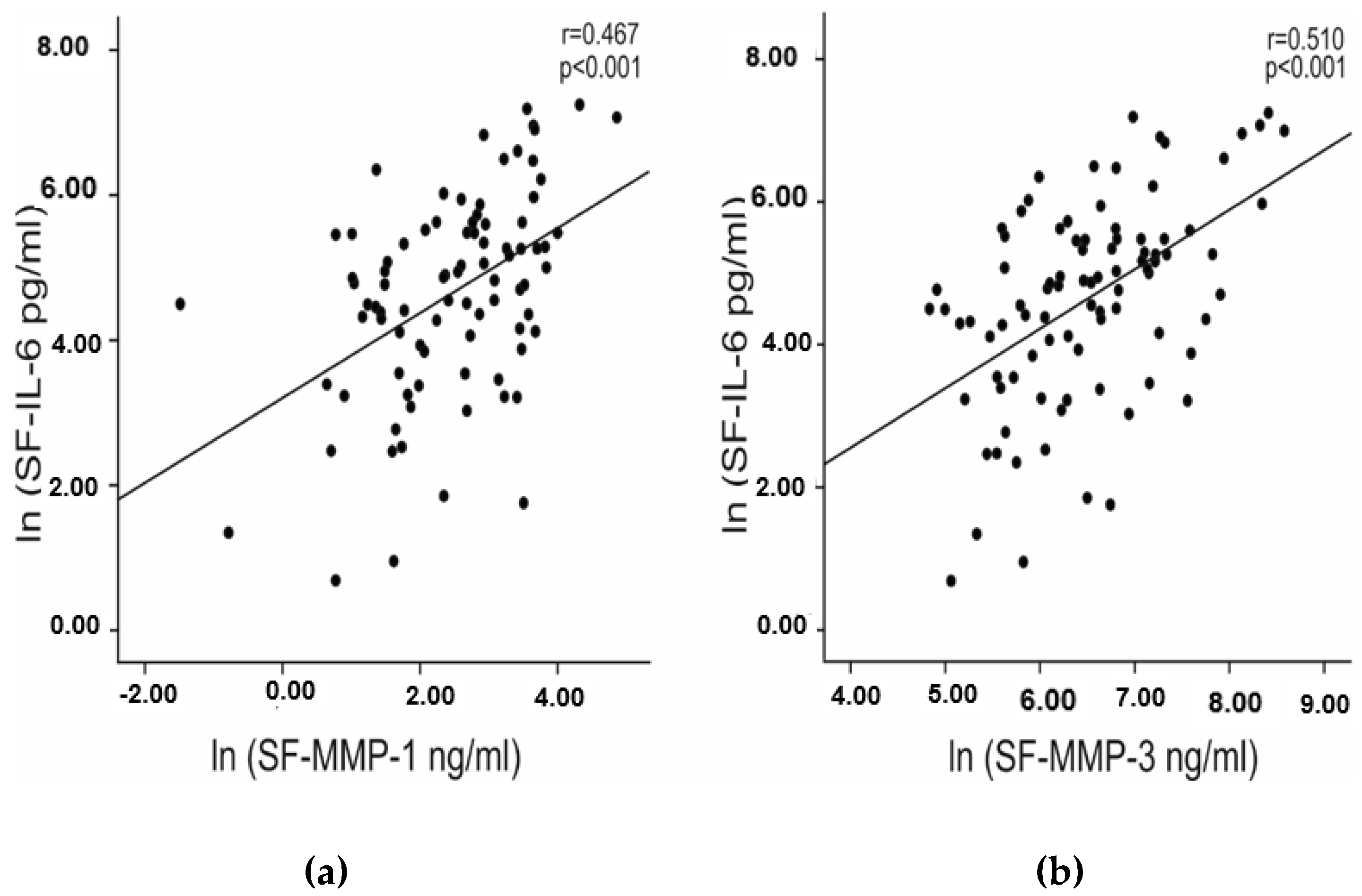
2.2. IL-6 Concentrations in Synovial Fluid Correlate with the Radiographic Severity of OA and with Matrix Metalloproteinase Concentrations
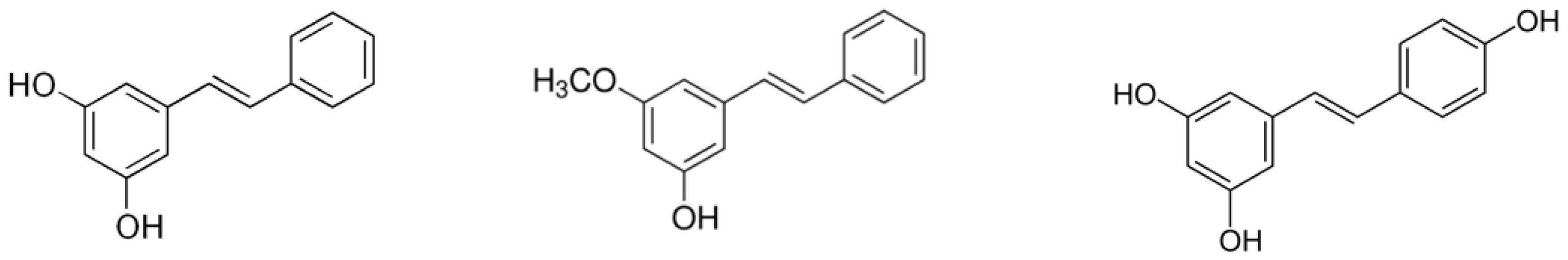
2.3. Monomethyl Pinosylvin, Pinosylvin and Resveratrol Suppress IL-6 Expression in Primary Cultures of OA Chondrocytes
2.4. Pine Stilbenoids Inhibit NF-κB Mediated Transcription in Human Chondrocytes
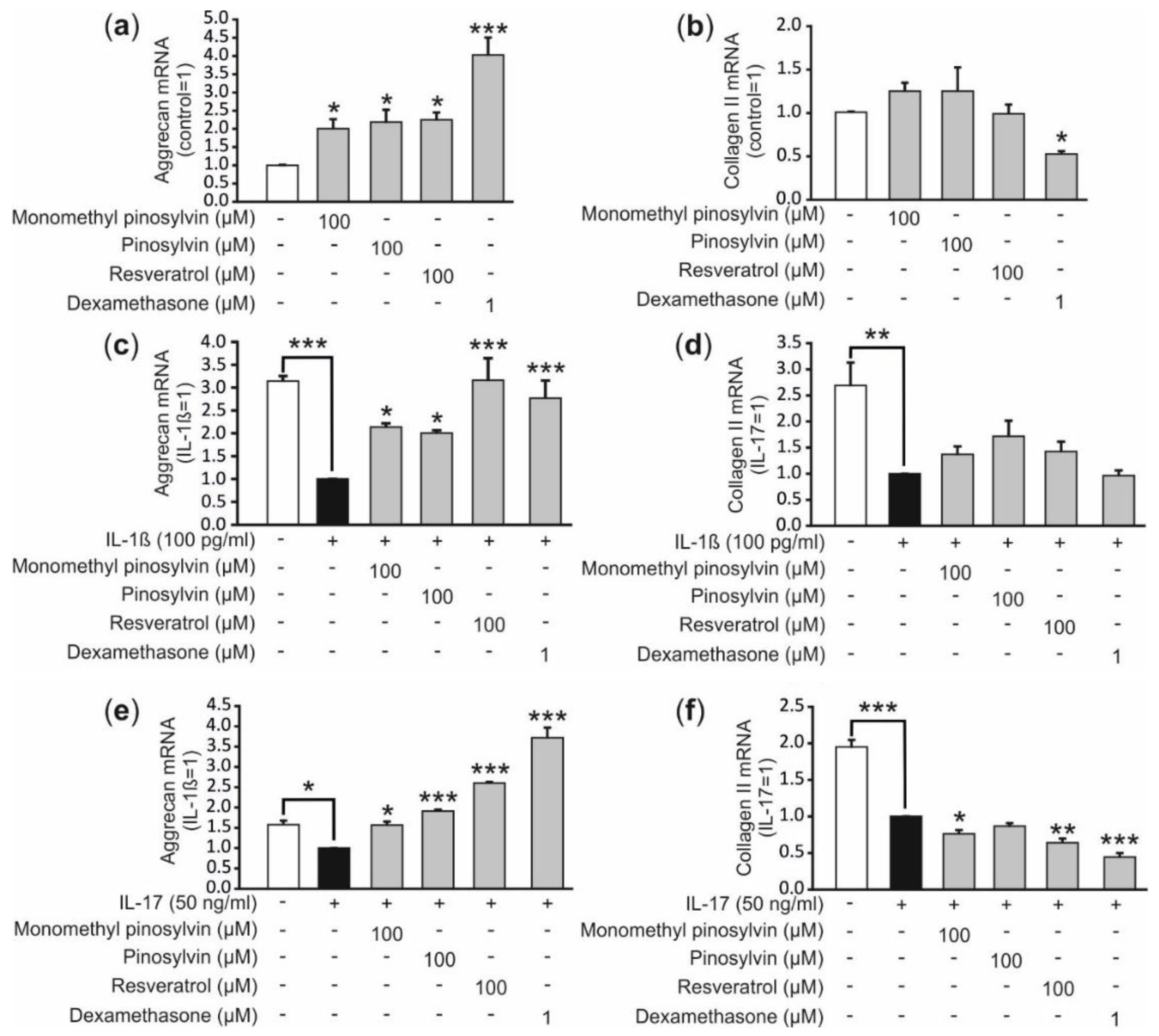
2.5. Pinosylvin, Monomethyl Pinosylvin and Resveratrol Enhance the Expression of the Anabolic Factor Aggrecan in Human Primary Chondrocytes
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Patients and Clinical Samples
4.3. Primary Chondrocyte Experiments
4.4. Preparation of the Stable T/C28a2pGL4.32NFκB Cell Line
4.5. Measurement of IL-6 and MMPs by Immunoassay
4.6. Measurement of IL-6, Collagen II and Aggrecan mRNA Levels
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Jones, S.A. IL-6 as a Keystone Cytokine in Health and Disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediators Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, X.; Eymard, F.; Richette, P. Biologic Agents in Osteoarthritis: Hopes and Disappointments. Nat. Rev. Rheumatol. 2013, 9, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Dancevic, C.M.; McCulloch, D.R. Current and Emerging Therapeutic Strategies for Preventing Inflammation and Aggrecanase-Mediated Cartilage Destruction in Arthritis. Arthritis Res. Ther. 2014, 16, 429. [Google Scholar] [CrossRef]
- Glyn-Jones, S.; Palmer, A.J.; Agricola, R.; Price, A.J.; Vincent, T.L.; Weinans, H.; Carr, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef]
- Kalunian, K.C. Current Advances in Therapies for Osteoarthritis. Curr. Opin. Rheumatol. 2016, 28, 246–250. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, L.; Jiang, H.; Zhou, J.; Tang, Y. Inhibition of Interleukin-6 Function Attenuates the Central Sensitization and Pain Behavior Induced by Osteoarthritis. Eur. J. Pharmacol. 2017, 811, 260–267. [Google Scholar] [CrossRef]
- Laavola, M.; Nieminen, R.; Leppänen, T.; Eckerman, C.; Holmbom, B.; Moilanen, E. Pinosylvin and Monomethylpinosylvin, Constituents of an Extract from the Knot of Pinus Sylvestris, Reduce Inflammatory Gene Expression and Inflammatory Responses In Vivo. J. Agric. Food Chem. 2015, 63, 3445–3453. [Google Scholar] [CrossRef]
- Park, E.J.; Min, H.Y.; Chung, H.J.; Ahn, Y.H.; Pyee, J.H.; Lee, S.K. Pinosylvin Suppresses LPS-Stimulated Inducible Nitric Oxide Synthase Expression Via the MyD88-Independent, but TRIF-Dependent Downregulation of IRF-3 Signaling Pathway in Mouse Macrophage Cells. Cell Physiol. Biochem. 2011, 27, 353–362. [Google Scholar] [CrossRef]
- Gu, H.; Jiao, Y.; Yu, X.; Li, X.; Wang, W.; Ding, L.; Liu, L. Resveratrol Inhibits the IL-1beta-Induced Expression of MMP-13 and IL-6 in Human Articular Chondrocytes Via TLR4/MyD88-Dependent and -Independent Signaling Cascades. Int. J. Mol. Med. 2017, 39, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of Proinflammatory Cytokines in the Pathophysiology of Osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Troeberg, L.; Nagase, H. Proteases Involved in Cartilage Matrix Degradation in Osteoarthritis. Biochim. Biophys Acta. 2012, 1824, 133–145. [Google Scholar] [CrossRef]
- Tsuchida, A.I.; Beekhuizen, M.; ′t Hart, M.C.; Radstake, T.R.; Dhert, W.J.; Saris, D.B.; van Osch, G.J.; Creemers, L.B. Cytokine Profiles in the Joint Depend on Pathology, but are Different between Synovial Fluid, Cartilage Tissue and Cultured Chondrocytes. Arthritis Res. Ther. 2014, 16, 441. [Google Scholar] [CrossRef] [PubMed]
- Beekhuizen, M.; Gierman, L.M.; van Spil, W.E.; Van Osch, G.J.; Huizinga, T.W.; Saris, D.B.; Creemers, L.B.; Zuurmond, A.M. An Explorative Study Comparing Levels of Soluble Mediators in Control and Osteoarthritic Synovial Fluid. Osteoarthritis Cartilage 2013, 21, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Livshits, G.; Zhai, G.; Hart, D.J.; Kato, B.S.; Wang, H.; Williams, F.M.; Spector, T.D. Interleukin-6 is a Significant Predictor of Radiographic Knee Osteoarthritis: The Chingford Study. Arthritis Rheum. 2009, 60, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cai, L.; Zhang, Y.; Cui, L.; Shen, G. Intra-Articular Resveratrol Injection Prevents Osteoarthritis Progression in a Mouse Model by Activating SIRT1 and Thereby Silencing HIF-2alpha. J. Orthop. Res. 2015, 33, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, J.S.; Chen, J.W.; Li, F.; Tian, J. Effect of Resveratrol on Cartilage Protection and Apoptosis Inhibition in Experimental Osteoarthritis of Rabbit. Rheumatol. Int. 2012, 32, 1541–1548. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.; Yang, Y.; Xia, F.; Huang, A.; Gao, X.; Fang, D.; Xiong, S.; Zhang, J. Resveratrol Inhibits CD4+ T Cell Activation by Enhancing the Expression and Activity of Sirt1. PLoS ONE 2013, 8, e75139. [Google Scholar] [CrossRef]
- Schwager, J.; Richard, N.; Riegger, C.; Salem, N., Jr. Omega-3 PUFAs and Resveratrol Differently Modulate Acute and Chronic Inflammatory Processes. Biomed. Res. Int. 2015, 2015, 535189. [Google Scholar] [CrossRef]
- Lee, A.S.; Ellman, M.B.; Yan, D.; Kroin, J.S.; Cole, B.J.; van Wijnen, A.J.; Im, H.J. A Current Review of Molecular Mechanisms regarding Osteoarthritis and Pain. Gene 2013, 527, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Shen, G.; Deng, X.; Lou, D.; Sun, B.; Wu, H.; Long, L.; Ding, T.; Zhao, J. The Role of IL-6 in Bone Marrow (BM)-Derived Mesenchymal Stem Cells (MSCs) Proliferation and Chondrogenesis. Cell Tissue Bank. 2013, 14, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Mathy-Hartert, M.; Hogge, L.; Sanchez, C.; Deby-Dupont, G.; Crielaard, J.M.; Henrotin, Y. Interleukin-1beta and interleukin-6 disturb the antioxidant enzyme system in bovine chondrocytes: A possible explanation for oxidative stress generation. Osteoarthritis Cartilage 2008, 7, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.Y.; Conquer, J.A.; Grinstein, S.; Cruz, T.F. Interleukin-1 beta induction of c-fos and collagenase expression in articular chondrocytes: Involvement of reactive oxygen species. J. Cell. Biochem. 1998, 1, 19–29. [Google Scholar] [CrossRef]
- Henrotin, Y.E.; Bruckner, P.; Pujol, J.P. The role of reactive oxygen species in homeostasis and degradation of cartilage. Osteoarthritis Cartilage 2003, 10, 747–755. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, A.G. ROS/oxidative stress signaling in osteoarthritis. Biochim. Biophys. Acta. 2016, 4, 576–591. [Google Scholar] [CrossRef]
- Yu, L.; Wang, S.; Kogure, Y.; Yamamoto, S.; Noguchi, K.; Dai, Y. Modulation of TRP Channels by Resveratrol and Other Stilbenoids. Mol. Pain. 2013, 9, 3. [Google Scholar] [CrossRef]
- Moilanen, L.J.; Hämäläinen, M.; Lehtimäki, L.; Nieminen, R.M.; Muraki, K.; Moilanen, E. Pinosylvin Inhibits TRPA1-Induced Calcium Influx in Vitro and TRPA1-Mediated Acute Paw Inflammation in vivo. Basic. Clin. Pharmacol. Toxicol. 2016, 118, 238–242. [Google Scholar] [CrossRef]
- Altman, R.; Asch, E.; Bloch, D.; Bole, G.; Borenstein, D.; Brandt, K.; Christy, W.; Cooke, T.D.; Greenwald, R.; Hochberg, M. Development of Criteria for the Classification and Reporting of Osteoarthritis. Classification of Osteoarthritis of the Knee. Diagnostic and Therapeutic Criteria Committee of the American Rheumatism Association. Arthritis. Rheum. 1986, 29, 1039–1049. [Google Scholar] [CrossRef]
- Koskinen, A.; Vuolteenaho, K.; Nieminen, R.; Moilanen, T.; Moilanen, E. Leptin Enhances MMP-1, MMP-3 and MMP-13 Production in Human Osteoarthritic Cartilage and Correlates with MMP-1 and MMP-3 in Synovial Fluid from OA Patients. Clin. Exp. Rheumatol. 2011, 29, 57–64. [Google Scholar]
- Goldring, M.B.; Birkhead, J.R.; Suen, L.F.; Yamin, R.; Mizuno, S.; Glowacki, J.; Arbiser, J.L.; Apperley, J.F. Interleukin-1 β-Modulated Gene Expression in Immortalized Human Chondrocytes. J. Clin. Investig. 1994, 94, 2307–2316. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Oligonucleotide | Sequence 5′→3′ |
|---|---|---|
| Human | Forward primer | TACCCCCAGGAGAAGATTCCA |
| Interleukin-6 | Reverse primer | CCGTCGAGGATGTACCGAATT |
| Probe | CGCCCCACACAGACAGCCACTC | |
| Human | Forward primer | GGCAATAGCAGGTTCACGTACA |
| Collagen II | Reverse primer | CGATAACAGTCTTGCCCCACTT |
| Probe | CTGAAGGATGGCTGCACGAAACATACC | |
| Human | Forward primer | GCCTGCGCTCCAATGACT |
| Aggrecanase | Reverse primer | TAATGGAACACGATGCCTTTCA |
| Probe | CCATGCATCACCTCGCAGCGGTA | |
| Human | Forward primer | AAGGTCGGAGTCAACGGATTT |
| GAPDH * | Reverse primer | GCAACAATATCCACTTTACCAGAGTTAA |
| Probe | CGCCTGGTCACCAGGGCTGC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laavola, M.; Leppänen, T.; Hämäläinen, M.; Vuolteenaho, K.; Moilanen, T.; Nieminen, R.; Moilanen, E. IL-6 in Osteoarthritis: Effects of Pine Stilbenoids. Molecules 2019, 24, 109. https://doi.org/10.3390/molecules24010109
Laavola M, Leppänen T, Hämäläinen M, Vuolteenaho K, Moilanen T, Nieminen R, Moilanen E. IL-6 in Osteoarthritis: Effects of Pine Stilbenoids. Molecules. 2019; 24(1):109. https://doi.org/10.3390/molecules24010109
Chicago/Turabian StyleLaavola, Mirka, Tiina Leppänen, Mari Hämäläinen, Katriina Vuolteenaho, Teemu Moilanen, Riina Nieminen, and Eeva Moilanen. 2019. "IL-6 in Osteoarthritis: Effects of Pine Stilbenoids" Molecules 24, no. 1: 109. https://doi.org/10.3390/molecules24010109
APA StyleLaavola, M., Leppänen, T., Hämäläinen, M., Vuolteenaho, K., Moilanen, T., Nieminen, R., & Moilanen, E. (2019). IL-6 in Osteoarthritis: Effects of Pine Stilbenoids. Molecules, 24(1), 109. https://doi.org/10.3390/molecules24010109