
Role of Saponins in Plant Defense Against Specialist Herbivores

 ,
,  , ,
, ,  , ,
, , 

Abstract
:1. Introduction
2. Plant Defense and Evolution
3. Chemical Variety of Secondary Metabolites
4. Balance of Costs and Benefits by Formation of New Compounds
5. Larval Feeding Preference and Adult Oviposition Behavior
5.1. P. xylostella Larval Survival on Cotyledons and True Leaves within the Same Plant
5.2. Saponins Presentation in B. Vulgaris Var Arcuata (Isolation and Identification)
6. Biological Significance of Saponins
6.1. Saponins Interference with the Feeding Behavior
6.2. Saponins Effects on Protein Digestion
6.3. Enterotoxicity
7. Limits of the Use of Saponins in Pest Management Control
8. Conclusions and Recommendations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Geyter, E.; Lambert, E.; Geelen, D.; Smagghe, G. Novel advances with plant saponins as natural insecticides to control pest insects. Pest Technol. 2007, 1, 96–105. [Google Scholar]
- Mugford, S.T.; Osbourn, A. Saponin synthesis and function. In Isoprenoid Synthesis in Plants and Microorganisms; Springer: New York, NY, USA, 2012; pp. 405–424. [Google Scholar]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [PubMed] [Green Version]
- Augustin, J.M.; Drok, S.; Shinoda, T.; Sanmiya, K.; Nielsen, J.K.; Khakimov, B.; Olsen, C.E.; Hansen, E.H.; Kuzina, V.; Ekstrøm, C.T. UDP-glycosyltransferases from the UGT73C subfamily in Barbarea vulgaris catalyze sapogenin 3-O-glucosylation in saponin-mediated insect resistance. Plant Physiol. 2012, 160, 1881–1895. [Google Scholar]
- Khakimov, B.; Kuzina, V.; Erthmann, P.Ø.; Fukushima, E.O.; Augustin, J.M.; Olsen, C.E.; Scholtalbers, J.; Volpin, H.; Andersen, S.B.; Hauser, T.P. Identification and genome organization of saponin pathway genes from a wild crucifer, and their use for transient production of saponins in Nicotiana benthamiana. Plant J. 2015, 84, 478–490. [Google Scholar] [PubMed]
- Patamalai, B.; Hejtmancik, E.; Bridges, C.; Hill, D.; Camp, B. The isolation and identification of steroidal sapogenins in Kleingrass. Vet. Hum. Toxicol. 1990, 32, 314–318. [Google Scholar] [PubMed]
- Lee, S.T.; Stegelmeier, B.L.; Gardner, D.R.; Vogel, K.P. The isolation and identification of steroidal sapogenins in switchgrass. J. Nat. Toxins 2001, 10, 273–281. [Google Scholar]
- Osbourn, A.E. Saponins in cereals. Phytochemistry 2003, 62, 1–4. [Google Scholar] [CrossRef]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S. Biosynthesis of Structurally Diverse Triterpenes in Plants: The Role of Oxidosqualene Cyclases. Proc. Indian Natl. Sci. Acad. 2016, 82, 1189–1210. [Google Scholar] [CrossRef]
- Nielsen, J.K.; Nagao, T.; Okabe, H.; Shinoda, T. Resistance in the plant, Barbarea vulgaris, and counter-adaptations in flea beetles mediated by saponins. J. Chem. Ecol. 2010, 36, 277–285. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R.; Gershenzon, J.; Heckel, D.G. Insect attraction versus plant defense: Young leaves high in glucosinolates stimulate oviposition by a specialist herbivore despite poor larval survival due to high saponin content. Plos One 2014, 9, e95766. [Google Scholar]
- Talekar, N.; Shelton, A. Biology, ecology, and management of the diamondback moth. Annu. Rev. Entomol. 1993, 38, 275–301. [Google Scholar] [CrossRef]
- Li, Z.; Feng, X.; Liu, S.-S.; You, M.; Furlong, M.J. Biology, Ecology, and Management of the Diamondback Moth in China. Annu. Rev. Entomol. 2016, 61, 277–296. [Google Scholar]
- Huang, X.; Renwick, J.; Sachdev-Gupta, K. Oviposition stimulants in Barbarea vulgaris for Pieris rapae and P. napi oleracea: Isolation, identification and differential activity. J. Chem. Ecol. 1994, 20, 423–438. [Google Scholar]
- Shinoda, T.; Nagao, T.; Nakayama, M.; Serizawa, H.; Koshioka, M.; Okabe, H.; Kawai, A. Identification of a triterpenoid saponin from a crucifer, Barbarea vulgaris, as a feeding deterrent to the diamondback moth, Plutella xylostella. J. Chem. Ecol. 2002, 28, 587–599. [Google Scholar]
- Serizawa, H.; Shinoda, T.; Kawai, A. Occurrence of a feeding deterrent in Barbarea vulgaris (Brassicales: Brassicaceae), a crucifer unacceptable to the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Appl. Entomol. Zool. 2001, 36, 465–470. [Google Scholar]
- Newman, K. Feeding and oviposition preferences of the diamondback moth Plutella xylostella (Lepidoptera: Plutellidae) on six Brassicaceae host plant species. M.Sc Thesis 2014. [Google Scholar]
- Hopkins, R.J.; van Dam, N.M.; van Loon, J.J. Role of glucosinolates in insect-plant relationships and multitrophic interactions. Annu. Rev. Entomol. 2009, 54, 57–83. [Google Scholar] [CrossRef]
- Ratzka, A.; Vogel, H.; Kliebenstein, D.J.; Mitchell-Olds, T.; Kroymann, J. Disarming the mustard oil bomb. Proc. Natl. Acad. Sci. USA 2002, 99, 11223–11228. [Google Scholar] [Green Version]
- Renwick, J.A.A.; Haribal, M.; Gouinguené, S.; Städler, E. Isothiocyanates stimulating oviposition by the diamondback moth, Plutella xylostella. J. Chem. Ecol. 2006, 32, 755–766. [Google Scholar]
- Hairston, N.G.; Smith, F.E.; Slobodkin, L.B. Community structure, population control, and competition. Am. Nat. 1960, 94, 421–425. [Google Scholar] [CrossRef]
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Dicke, M.; Sabelis, M.W. Does it pay plants to advertise for bodyguards? Towards a cost-benefit analysis of induced synomone production. In Causes and Consequences of Variation in Growth Rate and Productivity of Higher Plants; SPB Academic Publishing: The Hague, The Netherlands, 1989; pp. 341–358. [Google Scholar]
- Renwick, J.A.A. The chemical world of crucivores: Lures, treats and traps. Entomol. Exp. Et Appl. 2002, 104, 35–42. [Google Scholar] [CrossRef]
- Jones, C.G.; Firn, R.D.; Malcolm, S.B. On the evolution of plant secondary chemical diversity [and discussion]. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1991, 333, 273–280. [Google Scholar]
- Stahl, E. Pflanzen und Schnecken: Eine Biologische Studie über die Schutzmittel der Pflanzen gegen Schneckenfrass; G. Fischer: Germany, 1888; Volume 1. [Google Scholar]
- Cornell, H.V.; Hawkins, B.A. Herbivore responses to plant secondary compounds: A test of phytochemical coevolution theory. Am. Nat. 2003, 161, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, W.H. Plant defense belowground and spatiotemporal processes in natural vegetation. Ecology 2003, 84, 2269–2280. [Google Scholar] [CrossRef]
- Iwao, K.; Rausher, M.D. Evolution of plant resistance to multiple herbivores: Quantifying diffuse coevolution. Am. Nat. 1997, 149, 316–335. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Vet, L.E.M.; Harvey, J.A.; Wäckers, F.L. Linking above-and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists. Trends Ecol. Evol. 2001, 16, 547–554. [Google Scholar] [CrossRef]
- Van Dam, N.M.; Harvey, J.A.; Wäckers, F.L.; Bezemer, T.M.; van der Putten, W.H.; Vet, L.E. Interactions between aboveground and belowground induced responses against phytophages. Basic Appl. Ecol. 2003, 4, 63–77. [Google Scholar] [CrossRef]
- Bezemer, T.M.; van Dam, N.M. Linking aboveground and belowground interactions via induced plant defenses. Trends Ecol. Evol. 2005, 20, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Rask, L.; Andréasson, E.; Ekbom, B.; Eriksson, S.; Pontoppidan, B.; Meijer, J. Myrosinase: Gene family evolution and herbivore defense in Brassicaceae. In Plant Molecular Evolution; Springer: The Netherlands, 2000; pp. 93–113. [Google Scholar]
- Poveda, K.; Steffan-Dewenter, I.; Scheu, S.; Tscharntke, T. Effects of below-and above-ground herbivores on plant growth, flower visitation and seed set. Oecologia 2003, 135, 601–605. [Google Scholar] [CrossRef]
- Soler, R.; Harvey, J.A.; Kamp, A.F.D.; Vet, L.E.M.; Van der Putten, W.H.; Van Dam, N.M.; Stuefer, J.F.; Gols, R.; Hordijk, C.A.; Martijn Bezemer, T. Root herbivores influence the behaviour of an aboveground parasitoid through changes in plant-volatile signals. Oikos 2007, 116, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Fritz, R.S.; Simms, E.L. Plant Resistance to Herbivores and Pathogens: Ecology, Evolution, and Genetics; University of Chicago Press: Chicago, IL, USA, 1992. [Google Scholar]
- Glénsk, M.; Włodarczyk, M.; Bassarello, C.; Pizza, C.; Stefanowicz, P.; Świtalska, M. A Major Saponin from Leaves Extract of Acer velutinum. Z. Für Nat. B 2009, 64, 1081–1086. [Google Scholar] [CrossRef]
- Ribeiro, B.D.; Alviano, D.S.; Barreto, D.W.; Coelho, M.A.Z. Functional properties of saponins from sisal (Agave sisalana) and juá (Ziziphus joazeiro): Critical micellar concentration, antioxidant and antimicrobial activities. Colloids Surf. A Physicochem. Eng. Asp. 2013, 436, 736–743. [Google Scholar] [CrossRef]
- Mitaine-Offer, A.-C.; Marouf, A.; Hanquet, B.; Birlirakis, N.; Lacaille-Dubois, M.-A. Two Triterpene Saponins from Achyranthes bidentata. Chem. Pharm. Bull. 2001, 49, 1492–1494. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Ridout, C.L.; Price, K.R.; Parkin, G.; Dijoux, M.G.; Lavaud, C. Saponins from sugar beet and the floc problem. J. Agric. Food Chem. 1994, 42, 279–282. [Google Scholar] [CrossRef]
- Massiot, G.; Dijoux, M.-G.; Lavaud, C.; Le Men-Olivier, L.; Connolly, J.D.; Sheeley, D.M. Seco-glycosides of oleanolic acid fromBeta vulgaris. Phytochemistry 1994, 37, 1667–1670. [Google Scholar] [CrossRef]
- Lavaud, C.; Massiot, G.; Le Men-Olivier, L.; Viari, A.; Vigny, P.; Delaude, C. Saponins from Steganotaenia araliacea. Phytochemistry 1992, 31, 3177–3181. [Google Scholar] [CrossRef]
- Potter, D.A.; Kimmerer, T.W. Inhibition of herbivory on young holly leaves: Evidence for the defensive role of saponins. Oecologia 1989, 78, 322–329. [Google Scholar] [CrossRef]
- Shin, B.-K.; Kwon, S.W.; Park, J.H. Chemical diversity of ginseng saponins from Panax ginseng. J. Ginseng Res. 2015, 39, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Sun, C.; Sun, Y.; Wu, Q.; Li, Y.; Song, J.; Niu, Y.; Cheng, X.; Xu, H.; Li, C.; et al. Analysis of the transcriptome of Panax notoginseng root uncovers putative triterpene saponin-biosynthetic genes and genetic markers. Bmc Genom. 2011, 12, S5. [Google Scholar] [CrossRef] [PubMed]
- Miyakoshi, M.; Tamura, Y.; Masuda, H.; Mizutani, K.; Tanaka, O.; Ikeda, T.; Ohtani, K.; Kasai, R.; Yamasaki, K. Antiyeast steroidal saponins from Yucca schidigera (Mohave Yucca), a new anti-food-deteriorating agent. J. Nat. Prod. 2000, 63, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Piacente, S.; Pizza, C.; Oleszek, W. Saponins and phenolics of Yucca schidigera Roezl: Chemistry and bioactivity. Phytochem. Rev. 2005, 4, 177–190. [Google Scholar] [CrossRef]
- Chabani, S.; Lavaud, C.; Benkhaled, M.; Harakat, D.; Long, C.; Haba, H. Three new oleanane-type triterpene saponins from Atractylis flava. Phytochem. Lett. 2016, 15, 88–93. [Google Scholar] [CrossRef]
- Khakimov, B.; Amigo, J.M.; Bak, S.; Engelsen, S.B. Plant metabolomics: Resolution and quantification of elusive peaks in liquid chromatography–mass spectrometry profiles of complex plant extracts using multi-way decomposition methods. J. Chromatogr. A 2012, 1266, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Kuzina, V.; Ekstrøm, C.T.; Andersen, S.B.; Nielsen, J.K.; Olsen, C.E.; Bak, S. Identification of defense compounds in Barbarea vulgaris against the herbivore Phyllotreta nemorum by an ecometabolomic approach. Plant Physiol. 2009, 151, 1977–1990. [Google Scholar] [CrossRef] [PubMed]
- Kuzina, V.; Nielsen, J.K.; Augustin, J.M.; Torp, A.M.; Bak, S.; Andersen, S.B. Barbarea vulgaris linkage map and quantitative trait loci for saponins, glucosinolates, hairiness and resistance to the herbivore Phyllotreta nemorum. Phytochemistry 2011, 72, 188–198. [Google Scholar] [CrossRef]
- Ma, C.-H.; Gao, Z.-J.; Zhang, J.-J.; Zhang, W.; Shao, J.-H.; Hai, M.-R.; Chen, J.-W.; Yang, S.-C.; Zhang, G.-H. Candidate Genes Involved in the Biosynthesis of Triterpenoid Saponins in Platycodon grandiflorum Identified by Transcriptome Analysis. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Hostettmann, K.; Marston, A. Saponins; Cambridge University Press: New York, NY, USA, 2005. [Google Scholar]
- Koike, K.; Jia, Z.; Nikaido, T. New triterpenoid saponins and sapogenins from Saponaria officinalis. J. Nat. Prod. 1999, 62, 1655–1659. [Google Scholar] [CrossRef]
- Jia, Z.; Koike, K.; Sahu, N.P.; Nikaido, T. Triterpenoid saponins from Caryophyllaceae family. In Studies in Natural Products Chemistry; Atta ur, R., Ed.; Elsevier: Athens, GA, USA, 2002; Volume 26, Part G; pp. 3–61. [Google Scholar]
- Houghton, P.J.; Lian, L.M. Triterpenoids from Desfontainia spinosa. Phytochemistry 1986, 25, 1939–1944. [Google Scholar] [CrossRef]
- Simon, G.; Dewelle, J.; Nacoulma, O.; Guissou, P.; Kiss, R.; Daloze, D.; Braekman, J.C. Cytotoxic pentacyclic triterpenes from Combretum nigricans. Fitoterapia 2003, 74, 339–344. [Google Scholar] [CrossRef]
- Wang, C.Z.; Yu, D.Q. Triterpenoid Saponins from Aster auriculatus. J. Asian Nat. Prod. Res. 1998, 1, 1–14. [Google Scholar] [CrossRef]
- Dongliang, C.; Yu, S. Terpenoid glycosides from the roots of Aster tataricus. Phytochemistry 1993, 35, 173–176. [Google Scholar] [CrossRef]
- Sakai, K.; Nagao, T.; Okabe, H. Triterpenoid saponins from the ground part of Aster ageratoides var. ovatus. Phytochemistry 1999, 51, 309–318. [Google Scholar] [CrossRef]
- Shao, Y.; Zhou, B.-N.; Lin, L.-Z.; Cordell, G.A. Two new triterpenoid saponins, asterbatanoside F and G, from Aster batangensis. Nat. Prod. Lett. 1994, 5, 233–240. [Google Scholar] [CrossRef]
- Shao, Y.; Zhou, B.-N.; Lin, L.-Z.; Cordell, G.A. Triterpene saponins from Aster yunnanensis. Phytochemistry 1995, 38, 1487–1492. [Google Scholar] [CrossRef]
- Shao, Y.; Zhou, B.; Ma, K.; Wu, H.; Lin, L.; Cordell, G.A. Medicagenic acid saponins from Aster batangensis. Phytochemistry 1995, 39, 875–881. [Google Scholar] [CrossRef]
- Schöpke, T.; Al-Tawaha, C.; Wray, V.; Nimtz, M.; Meyer, A.; Hiller, K. Triterpenoid saponins from the aerial parts of Aster bellidiastrum. Phytochemistry 1995, 40, 1489–1492. [Google Scholar] [CrossRef]
- Schöpke, T.; Al-Tawaha, C.; Wray, V.; Nimtz, M.; Hiller, K. Triterpenoid saponins from Aster bellidiastrum. Phytochemistry 1997, 45, 125–132. [Google Scholar] [CrossRef]
- Shao, Y.; Ho, C.-T.; Chin, C.-K.; Poobrasert, O.; Yang, S.-W.; Cordell, G.A. Asterlingulatosides C and D, cytotoxic triterpenoid saponins from Aster lingulatus. J. Nat. Prod. 1997, 60, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Ho, C.-T.; Chin, C.-K.; Rosen, R.T.; Hu, B.; Qin, G.-W. Triterpenoid saponins from Aster lingulatus. Phytochemistry 1997, 44, 337–340. [Google Scholar] [CrossRef]
- Nagao, T.; Tanaka, R.; Okabe, H. Saponins from the compositae plants: Structures of the saponins from Aster scaber Thunb. In Saponins Used in Traditional and Modern Medicine; Springer: Boston, MA, USA, 1996; pp. 297–307. [Google Scholar]
- Corea, G.; Iorizzi, M.; Lanzotti, V.; Cammareri, M.; Conicella, C.; Laezza, C.; Bifulco, M. Astersedifolioside A–C, three new oleane-type saponins with antiproliferative activity. Bioorganic Med. Chem. 2004, 12, 4909–4915. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Zhou, B.-N.; Lin, L.-Z.; Cordell, G.A. Asteryunnanosides F and G: Two new triterpenoid saponins from Aster yunnanensis. Planta Med. 1995, 61, 446–449. [Google Scholar] [CrossRef]
- Shao, Y.; Zhou, B.-N.; Ma, K.; Wu, H.-M. Echinocystic acid saponins from Aster yunnanensis. J. Nat. Prod. 1995, 58, 837–842. [Google Scholar] [CrossRef]
- Shi, L.; Cao, J.-Q.; Shi, S.-M.; Zhao, Y.-Q. Triterpenoid saponins from Gynostemma pentaphyllum. J. Asian Nat. Prod. Res. 2011, 13, 168–177. [Google Scholar] [CrossRef]
- Yang, F.; Shi, H.; Zhang, X.; Yang, H.; Zhou, Q.; Yu, L.L. Two new saponins from tetraploid jiaogulan (Gynostemma pentaphyllum), and their anti-inflammatory and α-glucosidase inhibitory activities. Food Chem. 2013, 141, 3606–3613. [Google Scholar] [CrossRef]
- Keller, A.C.; Ma, J.; Kavalier, A.; He, K.; Brillantes, A.-M.B.; Kennelly, E.J. Saponins from the traditional medicinal plant Momordica charantia stimulate insulin secretion in vitro. Phytomedicine 2011, 19, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Waller, G.R.; Yamasaki, K. Saponins Used in Traditional and Modern Medicine; Springer Science & Business Media: Chicago, IL, USA, 2013; Volume 404. [Google Scholar]
- Qin, Y.; Wu, X.; Huang, W.; Gong, G.; Li, D.; He, Y.; Zhao, Y. Acute toxicity and sub-chronic toxicity of steroidal saponins from Dioscorea zingiberensis CH Wright in rodents. J. Ethnopharmacol. 2009, 126, 543–550. [Google Scholar] [CrossRef]
- Xu, R.; Zhao, W.; Xu, J.; Shao, B.; Qin, G. Studies on Bioactive Saponins from Chinese Medicinal Plants. In Saponins Used in Traditional and Modern Medicine; Waller, G.R., Yamasaki, K., Eds.; Springer US: Boston, MA, USA, 1996; pp. 371–382. [Google Scholar]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef]
- Massiot, G.; Lavaud, C.; Guillaume, D.; Le Men-Olivier, L. Reinvestigation of the sapogenins and prosapogenins from alfalfa (Medicago sativa). J. Agric. Food Chem. 1988, 36, 902–909. [Google Scholar] [CrossRef]
- Massiot, G.; Lavaud, C.; Besson, V.; Le Men-Olivier, L.; Van Binst, G. Saponins from aerial parts of alfalfa (Medicago sativa). J. Agric. Food Chem. 1991, 39, 78–82. [Google Scholar] [CrossRef]
- Oleszek, W.; Price, K.R.; Colquhoun, I.J.; Jurzysta, M.; Ploszynski, M.; Fenwick, G.R. Isolation and identification of alfalfa (Medicago sativa L.) root saponins: Their activity in relation to a fungal bioassay. J. Agric. Food Chem. 1990, 38, 1810–1817. [Google Scholar] [CrossRef]
- Rastogi, S.; Pandey, M.M.; Rawat, A.K.S. An ethnomedicinal, phytochemical and pharmacological profile of Desmodium gangeticum (L.) DC. and Desmodium adscendens (Sw.) DC. J. Ethnopharmacol. 2011, 136, 283–296. [Google Scholar] [CrossRef] [PubMed]
- McManus, O.; Harris, G.; Giangiacomo, K.; Feigenbaum, P.; Reuben, J.; Addy, M.; Burka, J.F.; Kaczorowski, G.; Garcia, M. An activator of calcium-dependent potassium channels isolated from a medicinal herb. Biochemistry 1993, 32, 6128–6133. [Google Scholar] [CrossRef] [PubMed]
- Dijoux, M.-G.; Lavaud, C.; Massiot, G.; Le Men-Olivier, L.; Sheeley, D.M. A saponin from leaves of Aphloia madagascariensis. Phytochemistry 1993, 34, 497–499. [Google Scholar] [CrossRef]
- Gopalsamy, N.; Vargas, D.; Guého, J.; Ricaud, C.; Hostettmann, K. Saponins from leaves of Aphloia theiformis. Phytochemistry 1988, 27, 3593–3595. [Google Scholar] [CrossRef]
- Kimura, H.; Ogawa, S.; Jisaka, M.; Kimura, Y.; Katsube, T.; Yokota, K. Identification of novel saponins from edible seeds of Japanese horse chestnut (Aesculus turbinata Blume) after treatment with wooden ashes and their nutraceutical activity. J. Pharm. Biomed. Anal. 2006, 41, 1657–1665. [Google Scholar] [CrossRef]
- Kimura, H.; Ogawa, S.; Katsube, T.; Jisaka, M.; Yokota, K. Antiobese effects of novel saponins from edible seeds of Japanese horse chestnut (Aesculus turbinata Blume) after treatment with wood ashes. J. Agric. Food Chem. 2008, 56, 4783–4788. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, S. Cytotoxic triterpenoid saponins from the fruits of Aesculus pavia L. Phytochemistry 2007, 68, 2075–2086. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, S.; Zhang, S.; Gorenstein, D. Triterpenoid saponins from the fruits of Aesculus pavia. Phytochemistry 2006, 67, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Murakami, T.; Matsuda, H.; Yamahara, J.; Murakami, N.; Kitagawa, I. Bioactive saponins and glycosides. III. Horse chestnut.(1): The structures, inhibitory effects on ethanol absorption, and hypoglycemic activity of escins Ia, Ib, IIa, IIb, and IIIa from the seeds of Aesculus hippocastanum L. Chem. Pharm. Bull. 1996, 44, 1454–1464. [Google Scholar] [CrossRef]
- Topcu, G.; Altiner, E.N.; Gozcu, S.; Halfon, B.; Aydogmus, Z.; Pezzuto, J.; Zhou, B.-N.; Kingston, D.G. Studies on di-and triterpenoids from Salvia staminea with cytotoxic activity. Planta Med. 2003, 69, 464–467. [Google Scholar]
- Massiot, G.; Xiang-Fei, C.; Lavaud, C.; Le Men-Olivier, L.; Delaude, C.; Viari, A.; Vigny, P.; Duval, J. Saponins from stem bark of Petersianthus macrocarpus. Phytochemistry 1992, 31, 3571–3576. [Google Scholar] [CrossRef]
- El Izzi, A.; Benie, T.; Thieulant, M.-L.; Le Men-Olivier, L.; Duval, J. Stimulation of LH release from cultured pituitary cells by saponins of Petersianthus macrocarpus: A permeabilizing effect. Planta Med. 1992, 58, 229–233. [Google Scholar] [CrossRef]
- Pal, B.C.; Chaudhuri, T.; Yoshikawa, K.; Arihara, S. Saponins from Barringtonia acutangula. Phytochemistry 1994, 35, 1315–1318. [Google Scholar] [CrossRef]
- Kawashima, K.; Mimaki, Y.; Sashida, Y. Steroidal saponins from Allium giganteum and A. aflatunense. Phytochemistry 1991, 30, 3063–3067. [Google Scholar] [CrossRef]
- Mimaki, Y.; Kuroda, M.; Sashida, Y. Steroidal saponins from the bulbs of Allium aflatunense. Natural Med. 1999, 53, 88–93. [Google Scholar]
- Ismailov, A.; Tagiev, S.; Rasulov, E. Steroid saponins and sapogenins from Allium rubellum and Allium albanum. Khimiia Prir. Soedin. Chem. Natural Compound. 1976, 12, 495. [Google Scholar] [CrossRef]
- Ismaĭlov, A.; Aliev, A. Determination of the steroid saponins in the onion (Allium albiflorus) that grows in Azerbaijan. Farmatsiia 1976, 25, 17. [Google Scholar]
- Mimaki, Y.; Kawashima, K.; Kanmoto, T.; Sashida, Y. Steroidal glycosides from Allium albopilosum and A. ostrowskianum. Phytochemistry 1993, 34, 799–805. [Google Scholar] [CrossRef]
- MoRITA, T.; USHIROGUCHI, T.; HAYASHI, N.; MATSUURA, H.; ITAKURA, Y.; Fuwa, T. Steroidal saponins from elephant garlic, bulbs of Allium ampeloprasum L. Chem. Pharm. Bull. 1988, 36, 3480–3486. [Google Scholar] [CrossRef]
- Matsunaga, S.; Fusetani, N.; Nishikawa, H.; Takamura, S.; Saito, T. New antifungal and cytotoxic steroidal saponins from the bulbs of an elephant garlic mutant. Biosci. Biotechnol. Biochem. 1998, 62, 1904–1911. [Google Scholar]
- Mimaki, Y.; Kuroda, M.; Sashida, Y. Steroidal saponins from the bulbs of Allium ampeloprasum. Natural Med. 1999, 53, 134–137. [Google Scholar]
- Fattorusso, E.; Iorizzi, M.; Lanzotti, V.; Taglialatela-Scafati, O. Chemical composition of shallot (Allium ascalonicum Hort.). J. Agric. Food Chem. 2002, 50, 5686–5690. [Google Scholar] [CrossRef]
- Corea, G.; Fattorusso, E.; Lanzotti, V.; Capasso, R.; Izzo, A.A. Antispasmodic saponins from bulbs of red onion, Allium cepa L. var. Tropea. J. Agric. Food Chem. 2005, 53, 935–940. [Google Scholar] [CrossRef]
- Dini, I.; Tenore, G.C.; Trimarco, E.; Dini, A. Furostanol saponins in Allium caepa L. Var. tropeana seeds. Food Chem. 2005, 93, 205–214. [Google Scholar] [CrossRef]
- Li, C.-J.; Yuan, L.; Ji, T.-F.; Yang, J.-B.; Wang, A.-G.; Su, Y.-L. Furostanol saponins from the seeds of Allium cepa L. Fitoterapia 2014, 99, 56–63. [Google Scholar] [CrossRef]
- Lanzotti, V.; Romano, A.; Lanzuise, S.; Bonanomi, G.; Scala, F. Antifungal saponins from bulbs of white onion, Allium cepa L. Phytochemistry 2012, 74, 133–139. [Google Scholar] [CrossRef]
- Kuroda, M.; Mimaki, Y.; Kameyama, A.; Sashida, Y.; Nikaido, T. Steroidal saponins from Allium chinense and their inhibitory activities on cyclic AMP phosphodiesterase and Na+ K+ ATPase. Phytochemistry 1995, 40, 1071–1076. [Google Scholar] [CrossRef]
- Peng, J.-P.; Yao, X.-S. 19 new steroidal saponins from Allium plants: Isolation, structural elucidation and effect on blood coagulability. In Saponins Used in Traditional and Modern Medicine; Springer: Boston, MA, USA, 1996; pp. 511–526. [Google Scholar]
- Jiang, Y.; Wang, N.-L.; Yao, X.-S.; Kitanaka, S. Steroidal saponins from the bulbs of Allium chinense. Stud. Plant Sci. 1999, 6, 212–219. [Google Scholar]
- Barile, E.; Zolfaghari, B.; Sajjadi, S.E.; Lanzotti, V. Saponins of Allium e lburzense. J. Nat. Prod. 2004, 67, 2037–2042. [Google Scholar] [CrossRef]
- Chincharadze, D.; Kel’Ginbaev, A.; Gorovits, M.; Eristavi, L.; Gorovits, T.; Abubakirov, N. Steroid saponins and sapogenins of Allium. XV. Eruboside B from Allium erubescens. Chem. Nat. Compd. 1979, 15, 442–446. [Google Scholar] [CrossRef]
- Kravets, S.; Vollerner, Y.S.; Gorovits, M.; Abubakirov, N. Steriods of the spirostan and furostan series from plants of the genus Allium. Chem. Nat. Compd. 1990, 26, 359–373. [Google Scholar] [CrossRef]
- Do, J.C.; Jung, K.Y.; Son, K.H. Steroidal saponins from the subterranean part of Allium fistulosum. J. Nat. Prod. 1992, 55, 168–173. [Google Scholar] [CrossRef]
- KELGINBAEV, A.; Gorovits, M.; Gorovits, T.; Abubakirov, N. Steroidal saponins and sapogenins of Allium. 9. Aginoside structure. Khimiya Prir. Soedin. 1976, 4, 480–486. [Google Scholar]
- SAsHiDA, Y.; KAwASHIMA, K.; MIMAKI, Y. Novel polyhydroxylated steroidal saponins from Allium giganteum. Chem. Pharm. Bull. 1991, 39, 698–703. [Google Scholar] [CrossRef]
- MIMAKI, Y.; NIKAIDO, T.; MATSUMOTO, K.; SASHIDA, Y.; OHMOTO, T. New steroidal saponins from the bulbs of Allium giganteum exhibiting potent inhibition of cAMP phosphodiesterase activity. Chem. Pharm. Bull. 1994, 42, 710–714. [Google Scholar] [CrossRef]
- Mimaki, Y.; Kuroda, M.; Fukasawa, T.; Sashida, Y. Steroidal glycosides from the bulbs of Allium jesdianum. J. Nat. Prod. 1999, 62, 194–197. [Google Scholar] [CrossRef]
- Mimaki, Y.; Kuroda, M.; Fukasawa, T.; SASHIDA, Y. Steroidal saponins from the bulbs of Allium karataviense. Chem. Pharm. Bull. 1999, 47, 738–743. [Google Scholar] [CrossRef]
- Vollerner, Y.S.; Abdullaev, N.; Gorovits, M.; Abubakirov, N. Steroid saponins and sapogenins of Allium. XX. Structure of karatavisoides E and F. Chem. Nat. Compd. 1984, 20, 64–68. [Google Scholar] [CrossRef]
- Inoue, T.; Mimaki, Y.; Sashida, Y.; Nishino, A.; Satomi, Y.; Nishino, H. Steroidal glycosides from Allium macleanii and A. senescens, and their inhibitory activity on tumour promoter-induced phospholipid metabolism of HeLa cells. Phytochemistry 1995, 40, 521–525. [Google Scholar] [CrossRef]
- Peng, J.; Yao, X.; Kobayashi, H.; Ma, C. Novel furostanol glycosides from Allium macrostemon. Planta Med. 1995, 61, 58–61. [Google Scholar] [CrossRef]
- Lazur’Evskii, G.; Krokhmalyuk, V.; Kintya, P. Structure of steroid glycosides of Allium narcissiflorum Wills. Dokl. Biochem. 1975, 221, 151. [Google Scholar]
- Krokhmalyuk, V.; Kintya, P. Steroid saponins. XIII. The structure of alliumosides D and E from Allium narcissiflorum. Chem. Nat. Compd. 1976, 12, 165–168. [Google Scholar] [CrossRef]
- Mimaki, Y.; Satou, T.; Ohmura, M.; Sashida, Y. Steroidal saponins from the bulbs of Allium narcissiflorum. Nat. Med. 1996, 50, 308. [Google Scholar]
- Akhov, L.; Musienko, M.; Piacente, S.; Pizza, C.; Oleszek, W. Structure of steroidal saponins from underground parts of Allium nutans L. J. Agric. Food Chem. 1999, 47, 3193–3196. [Google Scholar] [CrossRef]
- Akhov, L.; Musienko, M.; Shishova, Y.; Polishuk, V.; Oleszek, W. Biological Activity of Deltoside from Allium Nutans L. In Saponins in Food, Feedstuffs and Medicinal Plants; Springer: Dordrecht, The Netherlands, 2000; pp. 227–231. [Google Scholar]
- Harmatha, J.; Mauchamp, B.; Arnault, C.; Sláma, K. Identification of a spirostane-type saponin in the flowers of leek with inhibitory effects on growth of leek-moth larvae. Biochem. Syst. Ecol. 1987, 15, 113–116. [Google Scholar] [CrossRef]
- Carotenuto, A.; Fattorusso, E.; Lanzotti, V.; Magno, S. Spirostanol saponins of Allium porrum L. Phytochemistry 1999, 51, 1077–1082. [Google Scholar] [CrossRef]
- Fattorusso, E.; Lanzotti, V.; Taglialatela-Scafati, O.; Di Rosa, M.; Ianaro, A. Cytotoxic saponins from bulbs of Allium porrum L. J. Agric. Food Chem. 2000, 48, 3455–3462. [Google Scholar] [CrossRef]
- MATSUURA, H.; USHIROGUCHI, T.; ITAKURA, Y. Further studies on steroidal glycosides from bulbs, roots and leaves of Allium sativum L. Chem. Pharm. Bull. 1989, 37, 2741–2743. [Google Scholar] [CrossRef]
- Matsuura, H. Saponins in garlic as modifiers of the risk of cardiovascular disease. J. Nutr. 2001, 131, 1000S–1005S. [Google Scholar] [CrossRef]
- Lanzotti, V. Bioactive Saponins from Allium and Aster Plants. Phytochem. Rev. 2005, 4, 95–110. [Google Scholar] [CrossRef]
- Kawashima, K.; Mimaki, Y.; Sashida, Y. Steroidal saponins from the bulbs of Allium schubertii. Phytochemistry 1993, 32, 1267–1272. [Google Scholar] [CrossRef]
- Mimaki, Y.; Satou, T.; Kuroda, M.; Kameyama, A.; Sashida, Y.; Li, H.-y.; Harada, N. A new furostanol saponin with six sugars from the bulbs of Allium sphaerosephalon structural elucidation by modern NMR techniques. Chem. Lett. 1996, 25, 431–432. [Google Scholar] [CrossRef]
- Corea, G.; Fattorusso, E.; Lanzotti, V. Saponins and Flavonoids of Allium triquetrum. J. Nat. Prod. 2003, 66, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.-M.; Yu, D.-Q.; Cong, P.-Z. A steroidal saponin from the seeds of Allium tuberosum. Phytochemistry 2001, 57, 1219–1222. [Google Scholar] [CrossRef]
- Sang, S.; Mao, S.; Lao, A.; Chen, Z.; Ho, C.-T. New steroid saponins from the seeds of Allium tuberosum L. Food Chem. 2003, 83, 499–506. [Google Scholar] [CrossRef]
- Pirtskhalava, G.; Gorovits, M.; Gorovits, T.; Abubakirov, N. Steroid saponins and sapogenins of Allium. XVI. Turoside C from Allium turcomanicum. Chem. Nat. Compd. 1979, 15, 446–452. [Google Scholar] [CrossRef]
- Chen, S.; Snyder, J.K. Diosgenin-bearing, molluscicidal saponins from Allium vineale: An NMR approach for the structural assignment of oligosaccharide units. J. Org. Chem. 1989, 54, 3679–3689. [Google Scholar] [CrossRef]
- Magid, A.A.; Bobichon, H.; Borie, N.; Lalun, N.; Long, C.; Moretti, C.; Lavaud, C. Cytotoxic triterpenoid saponins from the stem bark of Antonia ovata. Phytochemistry 2010, 71, 429–434. [Google Scholar] [CrossRef]
- Lavaud, C.; Massiot, G.; Moretti, C.; Le Men-Olivier, L. Triterpene saponins Frommyrsine pellucida. Phytochemistry 1994, 37, 1671–1677. [Google Scholar] [CrossRef]
- Kitagawa, I.; Yoshikawa, M.; Kobayashi, K.; Imakura, Y.; Im, K.; Ikenishi, Y. Saponin and sapogenol. XXVIII. Reinvestigation of the branching positions in the glucuronide moieties of three glucuronide saponins: Desacyl-jegosaponin, desacyl-boninsaponin A, and sakuraso-saponin. Chem. Pharm. Bull. 1980, 28, 296–300. [Google Scholar] [CrossRef]
- Lavaud, C.; Pichelin, O.; Massiot, G.; Le Men-Olivier, L.; Sevenet, T.; Cosson, J. Sakuraso-saponin from Tapeinosperma clethroides. Fitoterapia 1999, 70, 116–118. [Google Scholar] [CrossRef]
- Lavaud, C.; Beauvière, S.; Massiot, G.; Le Men-Olivier, L.; Bourdy, G. Saponins from Pisonia umbellifera. Phytochemistry 1996, 43, 189–194. [Google Scholar] [CrossRef]
- Nhiem, N.X.; Thu, V.K.; Van Kiem, P.; Van Minh, C.; Tai, B.H.; Quang, T.H.; Cuong, N.X.; Yen, P.H.; Boo, H.-J.; Kang, J.-I.; et al. Cytotoxic oleane-type triterpene saponins from Glochidion eriocarpum. Arch. Pharmacal Res. 2012, 35, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Montoya, G.; Arango, G.J.; Unger, M.; Holzgrabe, U. Saponins from Phytolacca bogotensis using HPLC-ESI/multi-stage tandem mass spectrometry. Phytochem. Anal. 2009, 20, 465–474. [Google Scholar] [CrossRef]
- Bankefors, J.; Nord, L.I.; Kenne, L. Structural classification of acyl-substituted Quillaja saponins by electrospray ionisation ion trap multiple-stage mass spectrometry in combination with multivariate analysis. Rapid Commun. Mass Spectrom. 2008, 22, 3851–3860. [Google Scholar] [CrossRef] [PubMed]
- Kensil, C.R.; Patel, U.; Lennick, M.; Marciani, D. Separation and characterization of saponins with adjuvant activity from Quillaja saponaria Molina cortex. J. Immunol. 1991, 146, 431–437. [Google Scholar]
- Zhan, C.; Li, X.; Zhao, Z.; Yang, T.; Wang, X.; Luo, B.; Zhang, Q.; Hu, Y.; Hu, X. Comprehensive Analysis of the Triterpenoid Saponins Biosynthetic Pathway in Anemone flaccida by Transcriptome and Proteome Profiling. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, W.-M.; Fang, Q.-C. Triterpenoid saponins from Anemone flaccida. Planta Med. 1990, 56, 92–93. [Google Scholar] [CrossRef]
- Yuan, J.-Q.; Yang, X.-Z.; Miao, J.-H.; Tang, C.-P.; Ke, C.-Q.; Zhang, J.-B.; Ma, X.-J.; Ye, Y. New triterpene glucosides from the roots of Rosa laevigata Michx. Molecules 2008, 13, 2229–2237. [Google Scholar] [CrossRef]
- Lavaud, C.; Voutquenne, L.; Massiot, G.; Le Men-Olivier, L.; Delaude, C. Saponines triperpéniques de Smelophyllum capense (Sapindaceae). Bull. De La Société R. Des Sci. De Liège 1994, 63, 455–463. [Google Scholar]
- Lavaud, C.; Voutquenne, L.; Massiot, G.; Le Men-Olivier, L.; Das, B.C.; Laprévote, O.; Serani, L.; Delaude, C.; Becchi, M. Saponins from the stem bark of Filicium decipiens. Phytochemistry 1998, 47, 441–449. [Google Scholar] [CrossRef]
- Dimbi, M.Z.; Warin, R.; Delaude, C.; Huls, R.; Mpuza, K. Triterpenoides de Harpullia cupanioides. Bull. Des Sociétés Chim. Belg. 1983, 92, 473–484. [Google Scholar] [CrossRef]
- Voutquenne, L.; Lavaud, C.; Massiot, G.; Delaude, C. Saponins from Harpullia cupanioides. Phytochemistry 1998, 49, 2081–2085. [Google Scholar] [CrossRef]
- Shiau, I.-L.; Shih, T.-L.; Wang, Y.-N.; Chen, H.-T.; Lan, H.-F.; Lin, H.C.; Yang, B.-Y.; Ko, C.-H.; Murase, Y. Quantification for saponin from a soapberry (Sapindus mukorossi Gaertn) in cleaning products by a chromatographic and two colorimetric assays. J. Fac. Agr. Kyushu Univ. 2009, 54, 215–221. [Google Scholar]
- Massiot, G.; Lavaud, C.; Delaude, C.M.; Van Binst, G.; Miller, S.P.F.; Fales, H.M. Saponins from Tridesmostemon claessenssi. Phytochemistry 1990, 29, 3291–3298. [Google Scholar] [CrossRef]
- Wandji, J.; Tillequin, F.; Mulholland, D.A.; Shirri, J.C.; Tsabang, N.; Seguin, E.; Verite, P.; Libot, F.; Fomum, Z. Pentacyclic triterpenoid and saponins from Gambeya boukokoensis. Phytochemistry 2003, 64, 845–849. [Google Scholar] [CrossRef]
- Lavaud, C.; Massiot, G.; Becchi, M.; Misra, G.; Nigam, S. Saponins from three species of Mimusops. Phytochemistry 1996, 41, 887–893. [Google Scholar] [CrossRef]
- De Lucca, A.; Boue, S.; Palmgren, M.; Maskos, K.; Cleveland, T. Fungicidal properties of two saponins from Capsicum frutescens and the relationship of structure and fungicidal activity. Can. J. Microbiol. 2006, 52, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-H.; Shen, D.-D.; Li, N.; Yu, S.-S. Bioactive triterpenoids from Symplocos chinensis. J. Asian Nat. Prod. Res. 2003, 5, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Abliz, Z.; Tang, M.; Fu, G.; Yu, S. Rapid structural characterization of triterpenoid saponins in crude extract from Symplocos chinensis using liquid chromatography combined with electrospray ionization tandem mass spectrometry. J. Chromatogr. A 2006, 1101, 53–62. [Google Scholar] [CrossRef]
- Fu, G.-M.; Wang, Y.-H.; Gao, S.; Tang, M.-J.; Yu, S.-S. Five new cytotoxic triterpenoid saponins from the roots of Symplocos chinensis. Planta Med. 2005, 71, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Mitchell-Olds, T.; Clauss, M.J. Plant evolutionary genomics. Curr. Opin. Plant Biol. 2002, 5, 74–79. [Google Scholar] [CrossRef]
- Simms, E.L.; Rausher, M.D. Costs and benefits of plant resistance to herbivory. Am. Nat. 1987, 130, 570–581. [Google Scholar] [CrossRef]
- Vrieling, K.; Smit, W.; van der Meijden, E. Tritrophic interactions between aphids (Aphis jacobaeae Schrank), ant species, Tyria jacobaeae L. and Senecio jacobaea L. lead to maintenance of genetic variation in pyrrolizidine alkaloid concentration. Oecologia 1991, 86, 177–182. [Google Scholar] [CrossRef]
- Cipollini, D. Variation in the expression of chemical defenses in Alliaria petiolata (Brassicaceae) in the field and common garden. Am. J. Bot. 2002, 89, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- Van der Meijden, E.; Wijn, M.; Verkaar, H.J. Defence and regrowth, alternative plant strategies in the struggle against herbivores. Oikos 1988, 51, 355–363. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Gershenzon, J. Metabolic costs of terpenoid accumulation in higher plants. J. Chem. Ecol. 1994, 20, 1281–1328. [Google Scholar] [CrossRef] [PubMed]
- Strauss, S.Y.; Siemens, D.H.; Decher, M.B.; Mitchell-Olds, T. Ecological costs of plant resistance to herbivores in the currency of pollination. Evolution 1999, 53, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, G.; Janzen, D. Herbivores: Their interaction with plant secondary metabolites; Academic Press: New York, NY, USA, 1979; 718p. [Google Scholar]
- Hostettmann, K.; Marston, A.; Wolfender, J.L. In Strategy in the search for new biologically active plant constituents. In Proceedings-Phytochemical Society of Europe; Oxford University Press Inc.: Oxford, UK, 1995; p. 17. [Google Scholar]
- Abreu, A.C.; McBain, A.J.; Simoes, M. Plants as sources of new antimicrobials and resistance-modifying agents. Nat. Prod. Rep. 2012, 29, 1007–1021. [Google Scholar] [CrossRef]
- Harborne, J.B.; Baxter, H. Dictionary of Plant Toxins; John Wiley & Sons: UK, 1996. [Google Scholar]
- Morrissey, J.P.; Osbourn, A.E. Fungal resistance to plant antibiotics as a mechanism of pathogenesis. Microbiol. Mol. Biol. Rev. 1999, 63, 708–724. [Google Scholar] [PubMed]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental abiotic factors on the content of saponins in plants. Phytochem. Rev. 2011, 10, 471–491. [Google Scholar] [CrossRef]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 2016, 6, 29265. [Google Scholar] [CrossRef]
- Applebaum, S.; Birk, Y. Herbivores: Their Interactions with Secondary Plant Metabolites; Acad. Press: New York, NY, USA, 1979; pp. 539–566. [Google Scholar]
- Hostettmann, K.; Marston, A. Chemistry and Pharmacology of Natural Products, Saponin; Cambridge University Press: London, UK, 1995. [Google Scholar]
- Purkayastha, J.; Arora, R.; Singh, L. Sustainable and Novel Eco-friendly Approaches Towards Integrated Disease and Vector Management. In Herbal Insecticides, Repellents and Biomedicines: Effectiveness and Commercialization; Springer: New Delhi, India, 2016; pp. 11–23. [Google Scholar]
- Alias, E.E.M.; Zeinelabdin, M.H.; Bashir, N.H.H.; Assad, Y.O.H.; Abdelbagi, O.M. Hypoglycemic and toxic effects of saponins from the fruit of bitter apple [Citrullus colocynthis (L.) Schrad] on the internal organs of Norway rat [Rattus norvegicus (Berkenhout)]. Gezira J. Agric. Sci. 2015, 13, 1–17. [Google Scholar]
- Fuenzalida, T. Plant natural defense against insects: Role of secondary metabolites. Santiago 2015, 1–24. [Google Scholar]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu Rev Entomol 2013, 58, 517–541. [Google Scholar] [CrossRef]
- Heckel, D.G.; Gahan, L.J.; Liu, Y.B.; Tabashnik, B.E. Genetic mapping of resistance to Bacillus thuringiensis toxins in diamondback moth using biphasic linkage analysis. Proc. Natl. Acad. Sci. USA 1999, 96, 8373–8377. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Huang, F.; Ghimire, M.N.; Leonard, B.R.; Siegfried, B.D.; Rangasamy, M.; Yang, Y.; Wu, Y.; Gahan, L.J.; Heckel, D.G.; et al. Efficacy of genetically modified Bt toxins against insects with different genetic mechanisms of resistance. Nat. Biotechnol. 2011, 29, 1128–1131. [Google Scholar] [CrossRef]
- Shelton, A.M.; Badenes-Perez, F.R. Concepts and applications of trap cropping in pest management. Annu. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R.; Shelton, A.M.; Nault, B.A. Evaluating trap crops for diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2004, 97, 1365–1372. [Google Scholar] [CrossRef]
- Lu, J.H.; Liu, S.S.; Shelton, A.M. Laboratory evaluations of a wild crucifer Barbarea vulgaris as a management tool for the diamondback moth Plutella xylostella (Lepidoptera: Plutellidae). Bull. Entomol. Res. 2004, 94, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Shelton, A.M.; Nault, B.A. Dead-end trap cropping: A technique to improve management of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Crop Prot 2004, 23, 497–503. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R.; Shelton, A.M.; Nault, B.A. Using yellow rocket as a trap crop for diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 2005, 98, 884–890. [Google Scholar] [CrossRef]
- MacDonald, M.A.; Cavers, P.B. The biology of Canadian weeds: 97. Barbarea vulgaris R. Br. Can. J. Plant Sci. 1991, 71, 149–166. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Using plant chemistry and insect preference to study the potential of Barbarea (Brassicaceae) as a dead-end trap crop for diamondback moth (Lepidoptera: Plutellidae). Phytochemistry 2014, 98, 137–144. [Google Scholar] [CrossRef]
- Newman, K.; You, M.; Vasseur, L. Diamondback Moth (Lepidoptera: Plutellidae) Exhibits Oviposition and Larval Feeding Preferences Among Crops, Wild plants, and Ornamentals as Host Plants. J. Econ. Entomol. 2016, tow002. [Google Scholar] [CrossRef]
- Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.; Chen, S.; Hansen, C.; Olsen, C.; Halkier, B. Composition and content of glucosinolates in developing Arabidopsis thaliana. Planta 2002, 214, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Wallace, S.K.; Eigenbrode, S.D. Changes in the glucosinolate–myrosinase defense system in Brassica juncea cotyledons during seedling development. J. Chem. Ecol. 2002, 28, 243–256. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E.; Bibby, B.M.; Frandsen, H.O.; Brown, L.D.; Nielsen, J.K.; Renwick, J.A.A. A Saponin Correlated with Variable Resistance of Barbarea vulgaris to the Diamondback Moth Plutella xylostella. J. Chem. Ecol. 2003, 29, 1417–1433. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.J.; Nielsen, J.; Staerk, D. New resistance-correlated saponins from the insect-resistant crucifer Barbarea vulgaris. J. Agric. Food Chem. 2010, 58, 5509–5514. [Google Scholar] [CrossRef] [PubMed]
- van Mölken, T.; Kuzina, V.; Munk, K.R.; Olsen, C.E.; Sundelin, T.; van Dam, N.M.; Hauser, T.P. Consequences of combined herbivore feeding and pathogen infection for fitness of Barbarea vulgaris plants. Oecologia 2014, 175, 589–600. [Google Scholar] [CrossRef]
- De Geyter, E.; Swevers, L.; Caccia, S.; Geelen, D.; Smagghe, G. Saponins show high entomotoxicity by cell membrane permeation in Lepidoptera. Pest Manag. Sci. 2012, 68, 1199–1205. [Google Scholar] [CrossRef] [Green Version]
- Potter, S.M.; Jimenez-Flores, R.; Pollack, J.; Lone, T.A.; Berber-Jimenez, M.D. Protein-saponin interaction and its influence on blood lipids. J. Agric. Food Chem. 1993, 41, 1287–1291. [Google Scholar] [CrossRef]
- Soetan, K.; Ajibade, T.; Akinrinde, A. Saponins; A Ubiquitous Phytochemical: A Review of its Biochemical, Physiological and Pharmacological Effects. Recent Prog. Med. Plants 2014, 43, 1–24. [Google Scholar]
- Amtul, J.S.; Shakoori, A.R. Potential of Azadirachtin and Neem (Azadirachta indica) Based Saponins as Biopesticides for In vitro Insect Pests Cellulase (Beta-1, 4-Endoglucanase) Enzyme Inhibition and In vivo Repellency on Tribolium castaneum. Br. Biotechnol. J. 2014, 4, 904–917. [Google Scholar]
- Goławska, S.; Łukasik, I.; Wójcicka, A.; Sytykiewicz, H. Relationship between saponin content in alfalfa and aphid development. Acta Biol. Crac. Ser. Bot. 2012, 54, 39–46. [Google Scholar] [CrossRef]
- Tava, A.; Odoardi, M. Saponins from Medicago spp.: Chemical characterization and biological activity against insects. In Saponins Used in food and Agriculture; Springer: Boston, MA, USA, 1996; pp. 97–109. [Google Scholar]
- Vincken, J.-P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-A.; Lee, H.-Y.; Seo, E.; Lee, J.; Kim, S.-B.; Oh, S.; Choi, E.; Choi, E.; Lee, S.E.; Choi, D. Current Understandings on Plant Nonhost Resistance. Mol. Plant-Microbe Interact. 2017, 30, 5–15. [Google Scholar] [CrossRef]
- Dowd, P.F.; Berhow, M.A.; Johnson, E.T. Differential activity of multiple saponins against omnivorous insects with varying feeding preferences. J. Chem. Ecol. 2011, 37, 443–449. [Google Scholar] [CrossRef]
- Osbourn, A. Saponins and plant defence—A soap story. Trends Plant Sci. 1996, 1, 4–9. [Google Scholar] [CrossRef]
- Osbourn, A.E.; Wubben, J.P.; Melton, R.E.; Carter, J.P.; Daniels, M.J. Saponins and plant defense. In Phytochemical Signals and Plant-Microbe Interactions; Springer: Boston, MA, USA, 1998; pp. 1–15. [Google Scholar]
- Chaieb, I. Saponins as insecticides: A review. Tunis. J. Plant Prot. 2010, 5, 39–50. [Google Scholar]
- Akhtar, Y.; Isman, M.B. Binary mixtures of feeding deterrents mitigate the decrease in feeding deterrent response to antifeedants following prolonged exposure in the cabbage looper, Trichoplusia ni (Lepidoptera: Noctuidae). Chemoecology 2003, 13, 177–182. [Google Scholar] [CrossRef]
- Akhtar, Y.; Isman, M.B. Larval exposure to oviposition deterrents alters subsequent oviposition behavior in generalist, Trichoplusia ni and specialist, Plutella xylostella moths. J. Chem. Ecol. 2003, 29, 1853–1870. [Google Scholar] [CrossRef]
- Kumar, R.; Bhardwaj, U.; Kumar, P.; Mazumdar-Leighton, S. Midgut serine proteases and alternative host plant utilization in Pieris brassicae L. Front. Physiol. 2015, 6, 95. [Google Scholar] [CrossRef]
- Frisch, T.; Agerbirk, N.; Davis, S.; Cipollini, D.; Olsen, C.E.; Motawia, M.S.; Bjarnholt, N.; Møller, B.L. Glucosinolate-related glucosides in Alliaria petiolata: Sources of variation in the plant and different metabolism in an adapted specialist herbivore, Pieris rapae. J. Chem. Ecol. 2014, 40, 1063–1079. [Google Scholar] [CrossRef] [PubMed]
- Idris, A.B.; Grafius, E. The potential of using Barbarea vulgaris in insecticide-resistant diamondback moth management. Resist. Pest Manag. (USA) 1994, 6, 7–8. [Google Scholar]
- Badenes-Pérez, F.R.; Márquez, B.P.; Petitpierre, E. Can flowering Barbarea spp. (Brassicaceae) be used simultaneously as a trap crop and in conservation biological control? J. Pest Sci. 2016, 4, 1–11. [Google Scholar] [CrossRef]
- Badenes-Pérez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Phylloplane location of glucosinolates in Barbarea spp. (Brassicaceae) and misleading assessment of host suitability by a specialist herbivore. New Phytol. 2011, 189, 549–556. [Google Scholar] [CrossRef]
- Vermeer, K.M.C.A. Genetics of Insect Resistance to Plant Defence; Wageningen University: Wageningen, The Netherlands, 2014. [Google Scholar]
- Wittstock, U.; Gershenzon, J. Constitutive plant toxins and their role in defense against herbivores and pathogens. Curr. Opin. Plant Biol. 2002, 5, 300–307. [Google Scholar] [CrossRef]
- Mithöfer, A.; Maffei, M.E. General mechanisms of plant defense and plant toxins. In Plant Toxins; Springer AG: Switzerland, 2016; pp. 1–22. [Google Scholar]
- Sparg, S.G.; Light, M.E.; Van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef]
- Dinan, L. Phytoecdysteroids: Biological aspects. Phytochemistry 2001, 57, 325–339. [Google Scholar] [CrossRef]
- Harder, M.J.; Tello, V.E.; Giliomee, J.H. The acaricidal effect of ethanolic extracts of Chenopodium quinoa Willd. on Tetranychus urticae Koch (Acari: Tetranychidae). Afr. Entomol. 2016, 24, 50–60. [Google Scholar] [CrossRef]
- Nawrot, J.; Koul, O.; Isman, M.; Harmatha, J. Naturally occurring antifeedants: Effects on two polyphagous lepidopterans. J. Appl. Entomol. 1991, 112, 194–201. [Google Scholar] [CrossRef]
- Goławska, S.; Sprawka, I.; Łukasik, I. Effect of saponins and apigenin mixtures on feeding behavior of the pea aphid, Acyrthosiphon pisum Harris. Biochem. Syst. Ecol. 2014, 55, 137–144. [Google Scholar] [CrossRef]
- Pavela, R. Extract from the roots of Saponaria officinalis as a potential acaricide against Tetranychus urticae. J. Pest Sci. 2017, 90, 683–692. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Becker, K. The impact of saponins or saponin-containing plant materials on ruminant production—A review. J. Agric. Food Chem. 2005, 53, 8093–8105. [Google Scholar] [CrossRef]
- Tamura, Y.; Miyakoshi, M.; Yamamoto, M. Application of saponin-containing plants in foods and cosmetics. In Alternative Medicine; InTech: London, UK, 2012. [Google Scholar]
- Jia, Z.; Koike, K.; Nikaido, T. Major triterpenoid saponins from Saponaria officinalis. J. Nat. Prod. 1998, 61, 1368–1373. [Google Scholar] [CrossRef] [PubMed]
- Agerbirk, N.; Olsen, C.E.; Nielsen, J.K. Seasonal variation in leaf glucosinolates and insect resistance in two types of Barbarea vulgaris ssp. arcuata. Phytochemistry 2001, 58, 91–100. [Google Scholar] [CrossRef]
- Borah, B.; Phukon, P.; Hazarika, M.P.; Ahmed, R.; Sarmah, D.K.; Wann, S.B.; Das, A.; Bhau, B.S. Calamus leptospadix Griff. a high saponin yielding plant with antimicrobial property. Ind. Crop. Prod. 2016, 82, 127–132. [Google Scholar] [CrossRef]
- Lanzotti, V.; Barile, E.; Antignani, V.; Bonanomi, G.; Scala, F. Antifungal saponins from bulbs of garlic, Allium sativum L. var. Voghiera. Phytochemistry 2012, 78, 126–134. [Google Scholar] [CrossRef]


{kind=link}
| Family | Plant | Secondary Metabolite Form | Reference |
|---|---|---|---|
| Aceraceae | Acer velutinum | Td.S | [38] |
| Agavaceae | Agave sisalana | S.A | [39] |
| Amaranthaceae | Achyranthes bidentata | Te.S; Bidentatoside II and chikusetsusaponin V methyl ester. | [40] |
| Chenopodium quinoa | Td.S | [1,41] | |
| Beta vulgaris | Td.S | [42,43] | |
| Apiaceae | Steganotaenia araliacea | Td.S | [44] |
| Aquifoliaceae | Ilex opaca | [45] | |
| Araliaceae | Panax ginseng | Te.S; Ginsenosides, glycosides of triterpenoid aglycones | [46,47] |
| Asparagaceae | Yucca schidigera | S.S | [48,49] |
| Asparagus officinalis | S.S | [1,41] | |
| Asteraceae | Atractylis flava | Td.S | [50] |
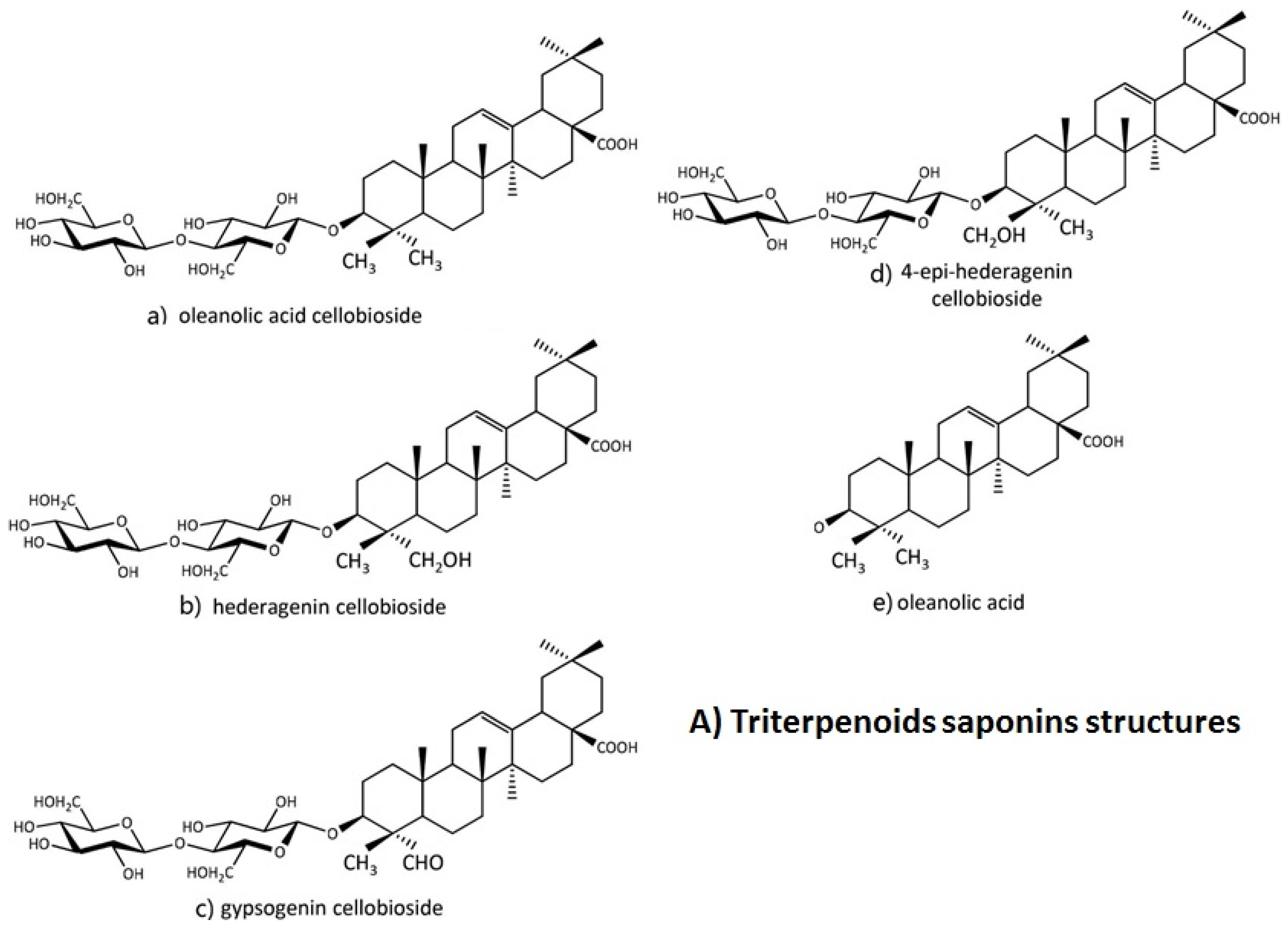
| Brassicacea | B. vulgaris | Td.S; hederagenincellobioside, oleanolic acid cellobioside, epihederagenincellobioside, and gypsogenincellobioside | [4,51,52,53] |
| Campanulaceae | Platycodon grandiflorum | Td.S | [54] |
| Caryophyllaceae | Saponaria officinalis | Td.S | [55,56,57] |
| Columelliaceae | Desfontainia spinose | Triterpenoids | [58] |
| Combretaceae | Combretum nigricans | Cytotoxic pentacyclic triterpenes | [59] |
| Compositae | Aster auriculatus | Td.S | [60] |
| As. tataricus | Triterpene glycoside | [61] | |
| As. ageratoides | Td.S | [62] | |
| As. batagensis | Td.S | [63,64,65] | |
| As. bellidiastrum | Td.S | [66,67] | |
| As. lingulatus | Td.S | [68,69] | |
| As. scaber | Td.S | [70] | |
| As. sedifolius | Oleane-type saponins; Astersedifolioside A, B and C | [71] | |
| As. yunnamensis | Td.S | [64,72,73] | |
| Cucurbitaceae | Gynostemma pentaphyllum | Gypenosides | [74,75] |
| Momordica charantia | Td.S | [76] | |
| Dioscoreae | Dioscorea spp. | Te.S, Dioscin | [77,78] |
| Fabaceae | Glycyrrhiza spp. | Glycyrrhizin; Td.S | [79,80] |
| Medicago sativa | Td.S | [81,82,83] | |
| Desmodium adscendens | Td.S | [84,85] | |
| Flacourtiaceae | Aphloia madagascariensis | Te.S | [86] |
| Flacourtiaceae | Aphloia theiformis | Te.S | [87] |
| Hippocastanaceae | Aesculus spp. | Td.S; Escins Polyhydroxyoleanene pentacyclic triterpenoid saponins; Aesculiosides | [88,89,90,91,92] |
| Lamiaceae | Salvia staminea | Td.S, salvistamineol | [93] |
| Lecythidaceae | Petersianthus macrocarpus | Td.S | [94,95] |
| Barringtonia acutangula | Monodesmosidic glucuronide saponins; Barringtosides A, B and C | [96] | |
| Liliaceae | Allium aflatunense | S.S | [97,98] |
| A. albanum | S.S | [99] | |
| A. albiflorus | S.S | [100] | |
| A. albopilosum | S.G | [101] | |
| A. ampeloprasum | S.S | [102,103,104] | |
| A. ascalonicum | S.S | [105] | |
| A. cepa | S.S; furostanol saponins, ceposide A, B, and C | [106,107,108,109] | |
| A. chinense | S.S | [110,111,112] | |
| A. elburzense | S.S | [113] | |
| A. erubescens | S.S | [114,115] | |
| A. fistulosum | S.S | [116] | |
| A. giganteum | S.S | [97,117,118,119] | |
| A. jesdianum | S.G | [120] | |
| A. karataviense | S.S | [121,122] | |
| A. macleanii | S.G | [123] | |
| A. macrostemon | Furostanol glycosides | [124] | |
| A. narcissiflorum | S.S | [125,126,127] | |
| A. nutans | S.S | [128,129] | |
| A. ostrowskianum | S.G | [101] | |
| A. porrum | Spirostane-type saponin | [130,131,132] | |
| A. sativum | S.S | [111,133,134,135] | |
| A. schubertii | S.S | [136] | |
| A. sphaerosephalon | Furostanol saponin | [137] | |
| A. senescens | S.G | [123] | |
| A. triquetrum | S.S | [138] | |
| A. tuberosum | S.S | [139,140] | |
| A. turcomanicum | S.S | [141] | |
| A. vineale | Molluscicidal saponins | [142] | |
| A. waldstenii | Steroids of spirostan and furostan series | [115] | |
| Loganiaceae | Antonia ovata | Td.S | [143] |
| Myrsinaceae | Myrsine pellucida | Te.S | [144] |
| Tapeinosperma clethroides | Glucuronide saponins: Desacyl-jegosaponin, desacylboninsaponin A, and sakuraso-saponin | [145,146] | |
| Nyctaginaceae | Pisonia umbellifera | Oleanolic acid saponins and Seco-glycopyranosyl moiety. | [147] |
| Phyllanthaceae | Glochidion eriocarpum | Cytotoxic oleane-type triterpene saponins | [148] |
| Phytolaccaceae | Phytolacca bogotensis | Te.S | [149] |
| Poaceae | Avena sativa | S.S | [1] |
| Quillajaceae | Quillaja saponaria | Te.S | [150,151] |
| Ranunculaceae | Anemone flaccida | Te.S | [152,153] |
| Rhamnaceae | Ziziphus joazeiro | Triterpenicaglycone | [39] |
| Rosaceae | Rosa laevigata | Triterpene glucosides | [154] |
| Sapindaceae | Smelophyllum capense | Te.S | [155] |
| Filicium decipiens | Te.S | [156] | |
| Harpullia cupanioides | Triterpenoïdes | [157,158] | |
| Sapindus mukorossi | [159] | ||
| Sapotaceae | Tridesmostemon claessenssi | Tridesmosaponin A and B | [160] |
| Gambeya boukokoensis | Gamboukokoensides A and B | [161] | |
| Mimusops spp. | Td.S | [162] | |
| Solanaceae | Solanum tuberosum | S.S | [1] |
| S. melongena | S.S | [1,41] | |
| Capsicum species | S.S; four glucose moieties and three glucose moieties | [1,163] | |
| Symplocaceae | Symplocos chinensis | Td.S | [164,165,166] |
| Theaceae | Camellia sinensis | Td.S | [41] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, M.; Debnath, B.; Qasim, M.; Bamisile, B.S.; Islam, W.; Hameed, M.S.; Wang, L.; Qiu, D. Role of Saponins in Plant Defense Against Specialist Herbivores. Molecules 2019, 24, 2067. https://doi.org/10.3390/molecules24112067
Hussain M, Debnath B, Qasim M, Bamisile BS, Islam W, Hameed MS, Wang L, Qiu D. Role of Saponins in Plant Defense Against Specialist Herbivores. Molecules. 2019; 24(11):2067. https://doi.org/10.3390/molecules24112067
Chicago/Turabian StyleHussain, Mubasher, Biswojit Debnath, Muhammad Qasim, Bamisope Steve Bamisile, Waqar Islam, Muhammad Salman Hameed, Liande Wang, and Dongliang Qiu. 2019. "Role of Saponins in Plant Defense Against Specialist Herbivores" Molecules 24, no. 11: 2067. https://doi.org/10.3390/molecules24112067
APA StyleHussain, M., Debnath, B., Qasim, M., Bamisile, B. S., Islam, W., Hameed, M. S., Wang, L., & Qiu, D. (2019). Role of Saponins in Plant Defense Against Specialist Herbivores. Molecules, 24(11), 2067. https://doi.org/10.3390/molecules24112067