
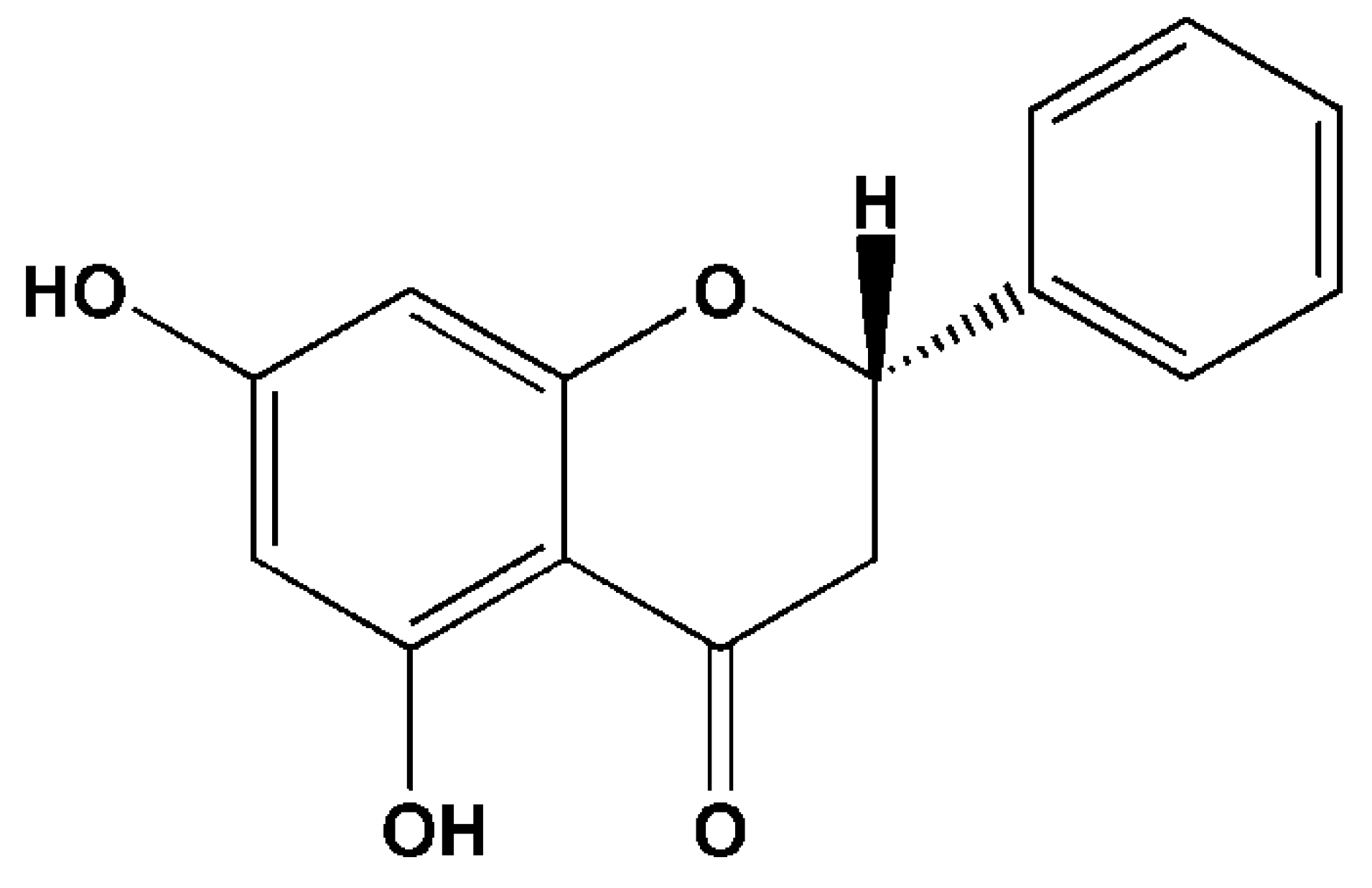
Advances in Biosynthesis, Pharmacology, and Pharmacokinetics of Pinocembrin, a Promising Natural Small-Molecule Drug

Abstract
:1. Introduction
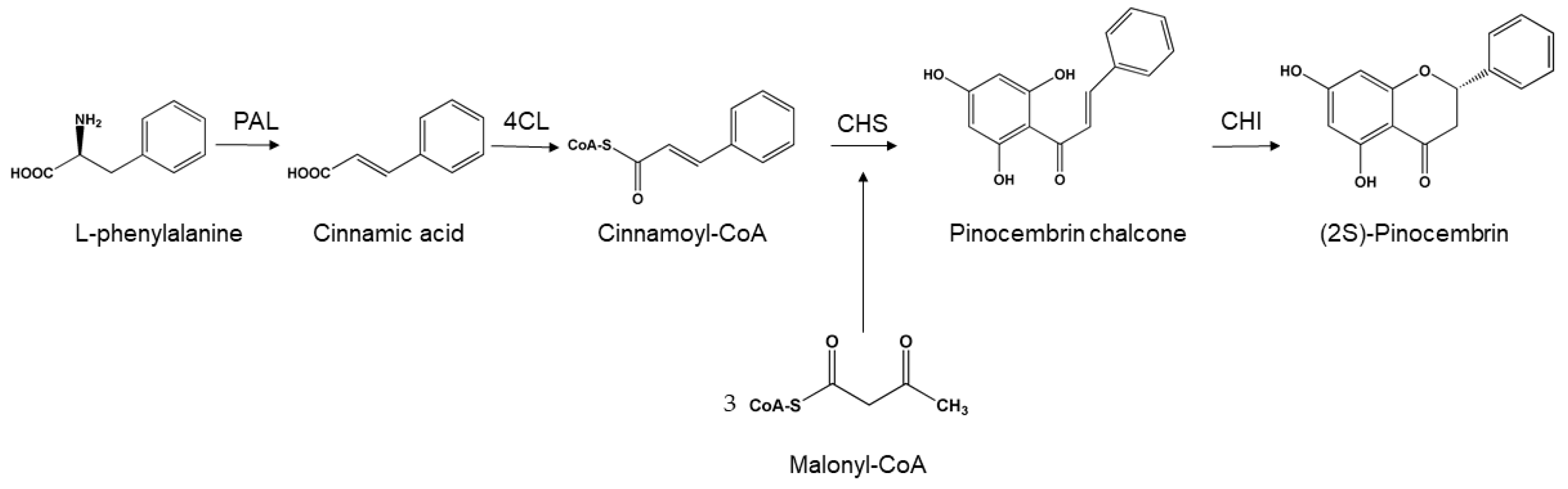
2. Microbial Biosynthesis
2.1. Synthesis of Pinocembrin from Glucose
2.2. Production Optimization Measures of Pinocembrin
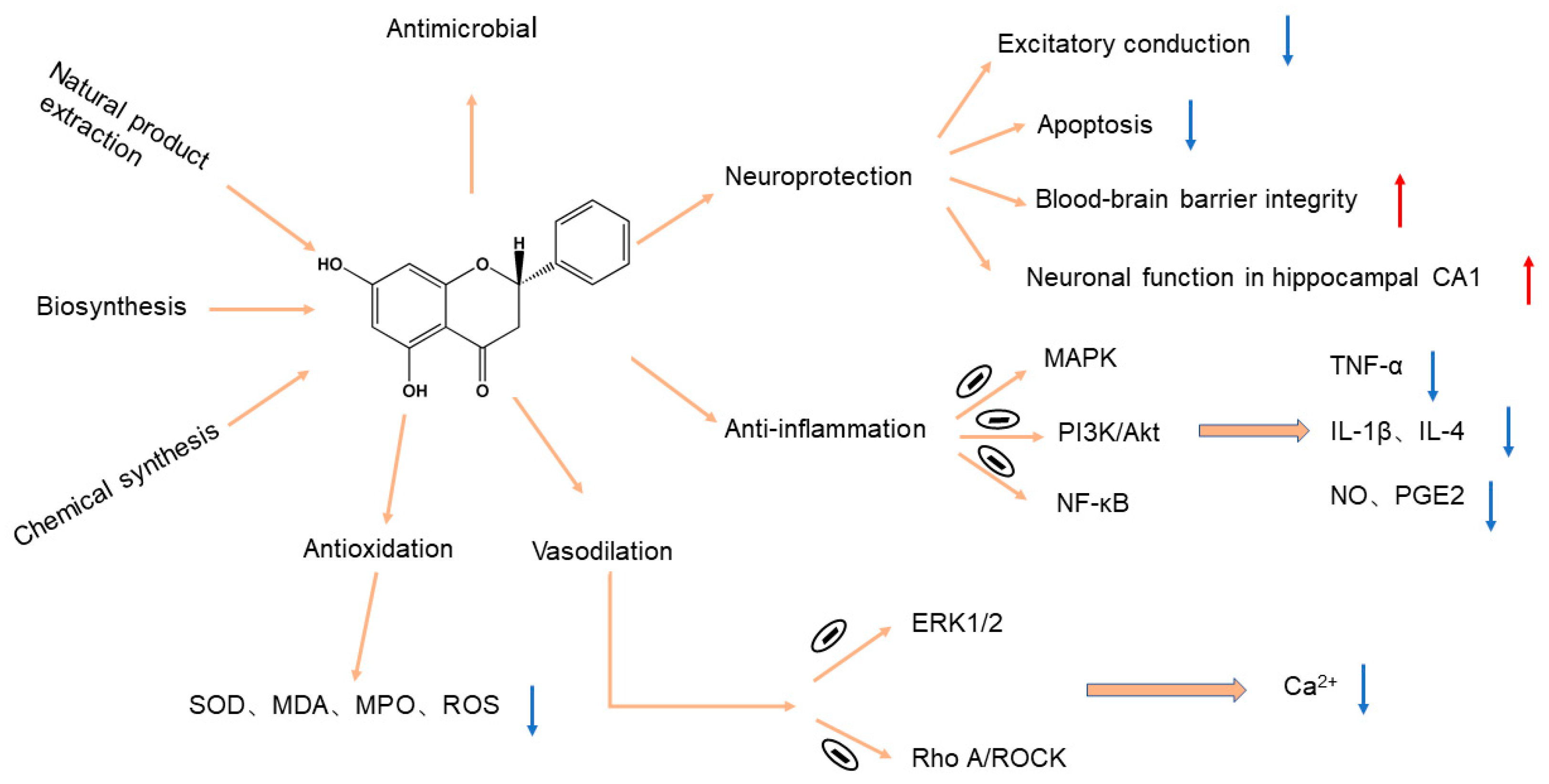
3. Pharmacological Effects of Pinocembrin
3.1. Neuroprotective Activity
3.1.1. Neuroprotective Effect in Cerebral Ischemia
3.1.2. Neuroprotective Effect of Pinocembrin in Alzheimer’s Disease and Parkinson’s Disease
3.2. Anti-Inflammation Activity
3.3. Antioxidation Activity
3.4. Antimicrobial Activity
3.5. Vasodilation Activity
3.6. Hepatoprotection Activity
3.7. Others
4. Pharmacokinetic Profiles of Pinocembrin
4.1. Transport Features Across Blood–Brain Barrier (BBB) In Vitro
4.2. Pharmacokinetic (PK) Profiles In Vivo
4.2.1. PK Profiles in Rats
4.2.2. PK Profiles in Humans
5. Conclusions and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Escriche, I.; Juan-Borrás, M. Standardizing the analysis of phenolic profile in propolis. Food Res. Int. 2018, 106, 834–841. [Google Scholar] [CrossRef] [PubMed]
- López, A.; Ming, D.S.; Towers, G.H. Antifungal activity of benzoic acid derivatives from Piper lanceaefolium. J. Nat. Prod. 2002, 65, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Danelutte, A.P.; Lago, J.H.; Young, M.C.; Kato, M.J. Antifungal flavanones and prenylated hydroquinones from Piper crassinervium Kunth. Phytochemistry 2003, 64, 555–559. [Google Scholar] [CrossRef]
- Feng, R.; Guo, Z.K.; Yan, C.M.; Li, E.G.; Tan, R.X.; Ge, H.M. Anti-inflammatory flavonoids from Cryptocarya chingii. Phytochemistry 2012, 76, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.X.; Du, G.H. Effects of pinocembrin on inflammation in rats with focal cerebral ischemia reperfusion injury. J. Shandong Med. College 2015, 37, 247–250. [Google Scholar]
- Said, M.M.; Azab, S.S.; Saeed, N.M.; El-Demerdash, E. Antifibrotic Mechanism of Pinocembrin: Impact on Oxidative Stress, Inflammation and TGF-β/Smad Inhibition in Rats. Ann. Hepatol. 2018, 17, 307–317. [Google Scholar] [CrossRef]
- Celerino de Moraes Porto, I.C.; Chaves Cardoso de Almeida, D.; Vasconcelos Calheiros de Oliveira Costa, G.; Sampaio Donato, T.S.; Moreira Nunes, L.; Gomes do Nascimento, T.; Dos Santos Oliveira, J.M.; Batista da Silva, C.; Barbosa Dos Santos, N.; de Alencar E Silva Leite, M.L.; et al. Mechanical and aesthetics compatibility of Brazilian red propolis micellar nanocomposite as a cavity cleaning agent. BMC Complement. Altern. Med. 2018, 18, 219. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Gao, M.; Yang, Z.H.; Du, G.H. Pinocembrin protects rat brain against oxidation and apoptosis induced by ischemia-reperfusion both in vivo and in vitro. Brain Res. 2008, 1216, 104–115. [Google Scholar] [CrossRef]
- Meng, F.R.; Liu, R.; Gao, M.; Wang, Y.H.; Yu, X.T.; Xuan, Z.H.; Sun, J.L.; Yang, F.; Wu, C.F.; Du, G.H. Pinocembrin attenuates blood-brain barrier injury induced by global cerebral ischemia-reperfusion in rats. Brain Res. 2011, 1391, 93–101. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, K.; Wu, Y.; Chen, Y.; Chen, X.; Hu, C.W.; Hu, F. Pinocembrin induces ER stress mediated apoptosis and suppresses autophagy in melanoma cells. Cancer Lett. 2018, 431, 31–42. [Google Scholar] [CrossRef]
- Zhou, F.Y.; Wang, A.Q.; Li, D.; Wang, Y.T.; Lin, L.G. Pinocembrin from Penthorumchinense Pursh suppresses hepatic stellate cells activation through a unified SIRT3-TGF-β-Smad signaling pathway. Toxicol. Appl. Pharmacol. 2018, 341, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Sun, X.; Qi, Y.; Mei, C.; Sun, X.B.; Du, G.H. Uptake characteristics of pinocembrin and its effect on p-glycoprotein at the blood-brain barrier in in vitro cell experiments. J. Asian Nat. Prod. Res. 2012, 14, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Du, G.C.; Zhou, J.W.; Chen, J. Metabolic engineering of Escherichia coli for (2S)-pinocembrin production from glucose by a modular metabolic strategy. Metab. Eng. 2013, 16, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Lee, H.; Ahn, J.H. Biosynthesis of pinocembrin from glucose using engineered escherichia coli. J. Microbiol. Biotechnol. 2014, 24, 1536–1541. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Kong, J.Q. Progress in synthetic biology of pinocembrin. Chin. J. Biotechnol. 2015, 31, 451–460. [Google Scholar]
- Cao, W.J.; Ma, W.C.; Zhang, B.; Wang, X.; Chen, K.Q.; Li, Y.; Ouyang, P.K. Improved pinocembrin production in Escherichia coli by engineering fatty acid synthesis. J. Ind. Microbiol. Biotechnol. 2016, 43, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.J.; Ma, W.C.; Wang, X.; Zhang, B.W.; Cao, X.; Chen, K.Q.; Li, Y.; Ouyang, P.Y. Enhanced pinocembrin production in Escherichia coli by regulating cinnamic acid metabolism. Sci. Rep. 2016, 6, 32640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Qian, Y.; Wang, X.; Cao, W.J.; Ma, W.C.; Chen, K.Q.; Ouyang, P.K. Regulation of ATP levels in Escherichia coli using CRISPR interference for enhanced pinocembrin production. Microb. Cell Fact. 2018, 17, 147–159. [Google Scholar] [CrossRef]
- Wu, J.J.; Zhang, X.; Zhou, J.W.; Dong, M.S. Efficient biosynthesis of (2S)-pinocembrin from d-glucose by integrating engineering central metabolic pathways with a pH-shift control strategy. Bioresour. Technol. 2016, 218, 999–1007. [Google Scholar] [CrossRef]
- Guo, L.; Chen, X.; Li, L.N.; Tang, W.; Pan, Y.T.; Kong, J.Q. Transcriptome-enabled discovery and functional characterization of enzymes related to (2S)-pinocembrin biosynthesis from Ornithogalum caudatum and their application for metabolic engineering. Microb. Cell Fact. 2016, 15, 27–45. [Google Scholar] [CrossRef]
- De Oliveira, M.R.; Peres, A.; Gama, C.S.; Bosco, S.M.D. Pinocembrin Provides Mitochondrial Protection by the Activation of the Erk1/2-Nrf2 Signaling Pathway in SH-SY5Y Neuroblastoma Cells Exposed to Paraquat. Mol. Neurobiol. 2017, 54, 6018–6031. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Zhang, W.C.; Liu, Q.S.; Hu, J.J.; Liu, G.T.; Du, G.H. Pinocembrin prevents glutamate-induced apoptosis in SH-SY5Y neuronal cells via decrease of bax/bcl-2 ratio. Eur. J. Pharmacol. 2008, 91, 73–79. [Google Scholar] [CrossRef]
- Feigin, V.L.; Mensah, G.A.; Norrving, B.; Murray, C.J.; Roth, G.A.; GBD 2013 Stroke Panel Experts Group. Atlas of the Global Burden of Stroke (1990-2013): The GBD 2013 Study. Neuroepidemiology 2015, 45, 230–236. [Google Scholar] [CrossRef]
- Ying, J.; Jiang, Y.D.; Chen, Y.; Samuel, S.; Du, G.H. Electrophysiological effects of pinocembrin on Aplysia SN/L7 co-cultures. Chin. Pharm. Bull. 2011, 27, 755–759. [Google Scholar]
- Gao, M.; Liu, R.; Zhu, S.Y.; Du, G.H. Acute neurovascular unit protective action of pinocembrin against permanent cerebral ischemia in rats. J. Asian Nat. Prod. Res. 2008, 10, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.X.; Du, G.H. Pinocembrin prevented brain acute injury induced by focal cerebral ischemia-reperfusion. Chin. J. Clin. Pharmacol. Ther. 2015, 20, 1208–1211, 1220. [Google Scholar]
- Ma, Y.Z.; Li, L.; Kong, L.L.; Zhu, Z.M.; Zhang, W.; Song, J.K.; Chang, J.L.; Du, G.H. Pinocembrin Protects Blood-Brain Barrier Function and Expands the Therapeutic Time Window for Tissue-Type Plasminogen Activator Treatment in a Rat Thromboembolic Stroke Model. Biomed. Res. Int. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Gao, M.; Zhu, S.Y.; Tan, C.B.; Xu, B.; Zhang, W.C.; Du, G.H. Pinocembrin protects the neurovascular unit by reducing inflammation and extracellular proteolysis in MCAO rats. J. Asian Nat. Prod. Res. 2010, 12, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.A.; Abdel Salam, R.M.; Kenawy, S.A.; Attia, A.S. Pinocembrin attenuates hippocampal inflammation, oxidative perturbations and apoptosis in a rat model of global cerebral ischemia reperfusion. Pharmacol. Rep. 2015, 67, 115–122. [Google Scholar] [CrossRef]
- Tao, J.H.; Shen, C.; Sun, Y.C.; Chen, W.M.; Yan, G.F. Neuroprotective effects of pinocembrin on ischemia/reperfusion-induced brain injury by inhibiting autophagy. Biomed. Pharmacother. 2018, 106, 1003–1010. [Google Scholar] [CrossRef]
- Guang, H.M.; Gao, M.; Zhu, S.Y.; He, X.L.; He, G.R.; Zhu, X.M.; Du, G.H. Effect of pinocembrin on Mitochondrial function in Rats with Acute focal Cerebral Ischemia. Chin. Pharm. Bull. 2012, 28, 24–29. [Google Scholar]
- Guang, H.M.; Du, G.H. Protections of pinocembrin on brain mitochondria contribute to cognitive improvement in chronic cerebral hypoperfused rats. Eur. J. Pharmacol. 2006, 542, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.L.; Qiang, G.F.; Gao, M.; Zhang, H.A.; Chen, B.N.; Yu, X.Y.; Xuan, Z.H.; Wang, Q.Y.; Du, G.H. Effect of pinocembrin on brain mitochondrial respiratory function. Acta Pharm. Sin. 2011, 46, 642–649. [Google Scholar]
- Zhang, P.F.; Xu, S.T.; Zhu, Z.Y.; Xu, J.Y. Multi-target design strategies for the improved treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 176, 228–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xie, X.X.; Ji, M.; Wang, S.W.; Zha, J.; Zhou, W.W.; Yu, X.L.; Wei, C.; Ma, S.; Xi, Z.Y.; et al. Naturally occurring autoantibodies against Aβ oligomers exhibited more beneficial effects in the treatment of mouse model of Alzheimer’s disease than intravenous immunoglobulin. Neuropharmacology 2016, 105, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wu, C.X.; Zhou, D.; Yang, F.; Tian, S.; Zhang, L.; Zhang, T.T.; Du, G.H. Pinocembrin protects against β-amyloid-induced toxicity in neurons through inhibiting receptor for advanced glycation end products (RAGE)-independent signaling pathways and regulating mitochondrion-mediated apoptosis. BMC Med. 2012, 10, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, J.Z.; Song, J.K.; Zhou, D.; Huang, C.; Bai, X.Y.; Xie, T.; Zhang, X.; Li, Y.J.; Wu, C.X.; et al. Pinocembrin improves cognition and protects the neurovascular unit in Alzheimer related deficits. Neurobiol. Aging 2014, 35, 1275–1285. [Google Scholar] [CrossRef]
- Liu, R.; Li, J.Z.; Song, J.K.; Sun, J.L.; Li, Y.J.; Zhou, S.B.; Zhang, T.T.; Du, G.H. Pinocembrin protects human brain microvascular endothelial cells against fibrillar amyloid-β (1-40) injury by suppressing the MAPK/NF-κB inflammatory pathways. Biomed. Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- Wang, Y.; Miao, Y.; Mir, A.Z.; Cheng, L.; Wang, L.; Zhao, L.; Cui, Q.; Zhao, W.; Wang, H. Inhibition of beta-amyloid-induced neurotoxicity by pinocembrin through Nrf2/HO-1 pathway in SH-SY5Y cells. J. Neurol. Sci. 2016, 368, 223–230. [Google Scholar] [CrossRef]
- Pieczenik, S.R.; Neustadt, J. Mitochondrial dysfunction and molecular pathways of disease. Exp. Mol. Pathol. 2007, 83, 84–92. [Google Scholar] [CrossRef]
- Wang, Y.M.; Gao, J.H.; Miao, Y.C.; Cui, Q.F.; Zhao, W.L.; Zhang, J.Y.; Wang, H.Q. Pinocembrin protects SH-SY5Y cells against MPP+-induced neurotoxicity through the mitochondrial apoptotic pathway. J. Mol. Neurosci. 2014, 53, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Wang, Y.M.; Zhao, L.N.; Cui, Q.F.; Wang, Y.H.; Du, G.H. Pinocembrin attenuates MPP+-induced neurotoxicity by the induction of heme oxygenase-1 through ERK1/2 pathway. Neurosci. Lett. 2016, 612, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.H.; Liu, Q.; Jia, L.L.; Li, M.; Wang, X. Pinocembrin attenuates 6-OHDA-induced neuronal cell death through Nrf2/ARE pathway in SH-SY5Y cells. Cell Mol. Neurobiol. 2015, 35, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Pei, B.; Sun, J. Pinocembrin alleviates cognition deficits by inhibiting inflammation in diabetic mice. J. Neuroimmunol. 2018, 314, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.Y.; Zhang, Q.; Du, Q.; Shen, H.; Zhu, Z.H. Pinocembrin attenuates allergic airway inflammation via inhibition of NF-κB pathway in mice. Int. Immuno. Pharmacol. 2017, 53, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.W.; Huang, B.; Xiong, C.J.; Yue, Z. Pinocembrin inhibits matrix metalloproteinase expression in chondrocytes. IUBMB Life 2015, 67, 36–41. [Google Scholar] [CrossRef]
- Zhou, L.T.; Wang, K.J.; Li, L.; Li, H.; Geng, M. Pinocembrin inhibits lipopolysaccharide-induced inflammatory mediators’ production in BV2 microglial cells through suppression of PI3K/Akt/NF-κB pathway. Eur. J. Pharmacol. 2015, 761, 211–216. [Google Scholar] [CrossRef]
- Soromou, L.W.; Chu, X.; Jiang, L.; Wei, M.; Huo, M.; Chen, N.; Guan, S.; Yang, X.; Chen, C.; Feng, H.; et al. In vitro and in vivo protection provided by pinocembrin against lipopolysaccharide-induced inflammatory responses. Int. Immuno. Pharmacol. 2012, 14, 66–74. [Google Scholar] [CrossRef]
- Shi, L.L.; Chen, B.N.; Gao, M.; Zhang, H.A.; Li, Y.J.; Wang, L.; Du, G.H. The characteristics of therapeutic effect of pinocembrin in transient global brain ischemia/reperfusion rats. Life Sci. 2011, 88, 521–528. [Google Scholar] [CrossRef]
- Su, Q.; Sun, Y.H.; Ye, Z.L.; Yang, H.F.; Kong, B.H.; Li, L. Pinocembrin protects endothelial cells from oxidized LDL-induced injury. Cytokine 2018, 111, 475–480. [Google Scholar] [CrossRef]
- Promsan, S.; Jaikumkao, K.; Pongchaidecha, A.; Chattipakorn, N.; Chatsudthipong, V.; Arjinajarn, P.; Pompimon, W.; Lungkaphin, A. Pinocembrin attenuates gentamicin-induced nephrotoxicity in rats. Can. J. Physiol. Pharmacol. 2016, 94, 808–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soromou, L.W.; Zhang, Y.; Cui, Y.; Wei, M.; Chen, N.; Yang, X.; Huo, M.; Baldé, A.; Guan, S.; Deng, X.M.; et al. Subinhibitory concentrations of pinocembrin exert anti-Staphylococcus aureus activity by reducing α-toxin expression. J. Appl. Microbiol. 2013, 115, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Gabaston, J.; Richard, T.; Cluzet, S.; Palos Pinto, A.; Dufour, M.C.; Corio-Costet, M.F.; Mérillon, J.M. Pinus pinaster Knot: A Source of Polyphenols against Plasmoparaviticola. J. Agric. Food Chem. 2017, 65, 8884–8891. [Google Scholar] [CrossRef] [PubMed]
- Melaku, Y.; Worku, T.; Tadesse, Y.; Mekonnen, Y.; Schmidt, J.; Arnold, N.; Dagne, E. Antiplasmodial Compounds from Leaves of Dodonaea angustifolia. Curr. Bioact. Compd. 2017, 13, 268–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tundis, R.; Frattaruolo, L.; Carullo, G.; Armentano, B.; Badolato, M.; Loizzo, M.R.; Aiello, F.; Cappello, A.R. An ancient remedial repurposing: Synthesis of new pinocembrin fatty acid acyl derivatives as potential antimicrobial/anti-inflammatory agents. Nat. Prod. Res. 2019, 33, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pang, X.B.; Chen, B.N.; Gao, L.; Wang, L.; Wang, S.B.; Wang, S.B.; Liu, D.P.; Du, G.H. Pinocembrin inhibits angiotensin II-induced vasoconstriction via suppression of the increase of Ca2+ and ERK1/2 activation through blocking AT1 R in the rat aorta. Biochem. Biophys. Res. Commun. 2013, 435, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, H.G.; Yuan, T.Y.; Zhao, Y.; Du, G.H. Rho kinase inhibition activity of pinocembrin in rat aortic rings contracted by angiotensin II. Chin. J. Nat. Med. 2013, 11, 258–263. [Google Scholar] [CrossRef]
- Zhu, X.M.; Fang, L.H.; Li, Y.J.; Du, G.H. Endothelium-dependent and -independent relaxation induced by pinocembrin in rat aortic rings. Vascul. Pharmacol. 2007, 46, 160–165. [Google Scholar] [CrossRef]
- Sun, Z.L.; Zhang, Y.Z.; Zhang, F.; Zhang, J.W.; Zheng, G.C.; Tan, L.; Wang, C.Z.; Zhou, L.D.; Zhang, Q.H.; Yuan, C.S. Quality assessment of Penthorum chinense Pursh through multicomponent qualification and fingerprint, chemometric, and antihepatocarcinoma analyses. Food Funct. 2018, 9, 3807–3814. [Google Scholar] [CrossRef]
- Wang, Y.H.; Huang, M.Z.; Xuan, H.Z.; Yi, X.S. Different biological activities of pinocembrin and luteolin. Apiculture China 2017, 68, 13–15. [Google Scholar]
- Zhang, P.; Xu, J.; Hu, W.; Yu, D.; Bai, X.L. Effects of Pinocembrin Pretreatment on Connexin 43 (Cx43) Protein Expression After Rat Myocardial Ischemia-Reperfusion and Cardiac Arrhythmia. Med. Sci. Monit. 2018, 24, 5008–5014. [Google Scholar] [CrossRef] [PubMed]
- Granados-Pineda, J.; Uribe-Uribe, N.; García-López, P.; Ramos-Godinez, M.D.P.; Rivero-Cruz, J.F.; Pérez-Rojas, J.M. Effect of Pinocembrin Isolated from Mexican Brown Propolis on Diabetic Nephropathy. Molecules 2018, 23, 852. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liang, X.; Zhang, G.; Kong, L.; Peng, W.; Zhang, H. Galangin and Pinocembrin from Propolis Ameliorate Insulin Resistance in HepG2 Cells via Regulating Akt/mTOR Signaling. Evid. Based Complement. Alternat. Med. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ajmala Shireen, P.; Abdul Mujeeb, V.M.; Muraleedharan, K. Theoretical insights on flavanones as antioxidants and UV filters: A TDDFT and NLMO study. J. Photochem. Photobiol. B 2017, 170, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Ying, J.; Du, G.H. A New Method for Quantitative Study of Drug Disposal and Its Application. Chin. Pharm. Bull. 2009, 25, 323. [Google Scholar]
- Sayre, C.L.; Alrushaid, S.; Martinez, S.E.; Anderson, H.D.; Davies, N.M. Pre-Clinical Pharmacokinetic and Pharmacodynamic Characterization of Selected Chiral Flavonoids: Pinocembrin and Pinostrobin. J. Pharm. Phar. Sci. 2015, 18, 368–395. [Google Scholar] [CrossRef] [Green Version]
- Sayre, C.L.; Takemoto, J.K.; Martinez, S.E.; Davies, N.M. Chiral analytical method development and application to pre-clinical pharmacokinetics of pinocembrin. Biomed. Chromatogr. 2013, 27, 681–684. [Google Scholar] [CrossRef]
- Guo, W.W.; Qiu, F.; Chen, X.Q.; Ba, Y.Y.; Wang, X.; Wu, X. In-vivo absorption of pinocembrin-7-O-β-D-glucoside in rats and its in-vitro biotransformation. Sci. Rep. 2016, 6, 29340. [Google Scholar] [CrossRef]
- Yan, B.; Cao, G.Y.; Yan, J.L.; Hu, X.; Peng, Y.Y.; Bian, Z.R.; Li, K.X. Determination of Pinocembrin in Human Urine by LC-MS / MS and Research on Urinary Excretion. Chin. Pharm. J. 2014, 49, 1540–1544. [Google Scholar]
- Wang, D.M.; Liu, H.T.; Tong, Y.F.; Wu, S. Synthesis of two metabolites of pinocembrin. Chin. J. New Drugs 2013, 22, 1130–1132, 1165. [Google Scholar]
- Yan, B.; Cao, G.Y.; Sun, T.H.; Zhao, X.; Hu, X.; Yan, J.L.; Peng, Y.Y.; Shi, A.X.; Li, Y.; Xue, W.; et al. Determination of pinocembrin in human plasma by solid-phase extraction and LC/MS/MS: Application to pharmacokinetic studies. Biomed. Chromatogr. 2014, 28, 1601–1606. [Google Scholar] [CrossRef]
- Cao, G.Y.; Ying, P.Y.; Yan, B.; Xue, W.; Li, K.X.; Shi, A.X.; Sun, T.H.; Yan, J.L.; Hu, X. Pharmacokinetics, safety, and tolerability of single and multiple-doses of pinocembrin injection administered intravenously in healthy subjects. J. Ethnopharmacol. 2015, 168, 31–36. [Google Scholar] [CrossRef]
- ŠarićMustapić, D.; Debeljak, Ž.; Maleš, Ž.; Bojić, M. The Inhibitory Effect of Flavonoid Aglycones on the Metabolic Activity of CYP3A4 Enzyme. Molecules 2018, 23, 2553. [Google Scholar] [CrossRef]
- Navrátilová, L.; Ramos Mandíková, J.; Pávek, P.; Mladěnka, P.; Mladěnka, P.; Trejtnar, F. Honey flavonoids inhibit hOATP2B1 and hOATP1A2 transporters and hOATP-mediated rosuvastatin cell uptake in vitro. Xenobiotica 2018, 48, 745–755. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Subject | Mode of Administration | Dose (mg/kg) | AUC (h*µg/mL) | Vd (L/kg) | CLtotal (L/h/kg) | T1/2 Serum (h) | References |
|---|---|---|---|---|---|---|---|
| SD rats | iv. | 10 | S-1.821 ± 0.211; R-1.876 ± 0.427 | S-1.758 ± 1.313; R-1.793 ± 0.805 | S-5.527 ± 0.641; R-5.535 ± 1.217 | S-0.212 ± 0.140; R-0.223 ± 0.083 | [67] |
| SD rats | iv. | 10 | S-1.83 ± 0.092; R-1.876 ± 0.312 | S-1.46 ± 0.591; R-1.80 ± 0.271 | S-5.44 ± 0.287; R-5.83 ± 0.865 | S-0.262 ± 0.071; R-0.263 ± 0.027 | [66] |
| SD rats | po. | 100 | S-570 ± 21.7; R-531 ± 82.1 | S-3.80 ± 1.34; R-5.14 ± 1.81 | S-2.82 ± 0.084; R-2.83 ± 0.844 | S-20.3 ± 8.41; R-27.1 ± 18.8 | [66] |
| SD rats | iv. | 10 | 0.686.1 ± 0.0651 | 48.7 ± 19.6 | 15.5 ± 1.4 | 2.14 ± 0.68 | [68] |
| SD rats | po. | 50 | 0.518 ± 0.170 | 478 ± 213 | 110 ± 31.4 | 3.11 ± 1.21 | [68] |
| Human | iv. | 20 (mg) | 10.3381 ± 1.5394 (min μg/mL) | 136.6 ± 52.8 (L) | 2.0 ± 0.3 (L/min) | 0.79 ± 0.23 | [71] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, X.; Liu, Y.; Luo, X.; Yang, Z. Advances in Biosynthesis, Pharmacology, and Pharmacokinetics of Pinocembrin, a Promising Natural Small-Molecule Drug. Molecules 2019, 24, 2323. https://doi.org/10.3390/molecules24122323
Shen X, Liu Y, Luo X, Yang Z. Advances in Biosynthesis, Pharmacology, and Pharmacokinetics of Pinocembrin, a Promising Natural Small-Molecule Drug. Molecules. 2019; 24(12):2323. https://doi.org/10.3390/molecules24122323
Chicago/Turabian StyleShen, Xiaoling, Yeju Liu, Xiaoya Luo, and Zhihong Yang. 2019. "Advances in Biosynthesis, Pharmacology, and Pharmacokinetics of Pinocembrin, a Promising Natural Small-Molecule Drug" Molecules 24, no. 12: 2323. https://doi.org/10.3390/molecules24122323
APA StyleShen, X., Liu, Y., Luo, X., & Yang, Z. (2019). Advances in Biosynthesis, Pharmacology, and Pharmacokinetics of Pinocembrin, a Promising Natural Small-Molecule Drug. Molecules, 24(12), 2323. https://doi.org/10.3390/molecules24122323