
High Concentrations of Polyelectrolyte Complex Nanoparticles Decrease Activity of Osteoclasts

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Live Cell Imaging of Osteoclastogenesis
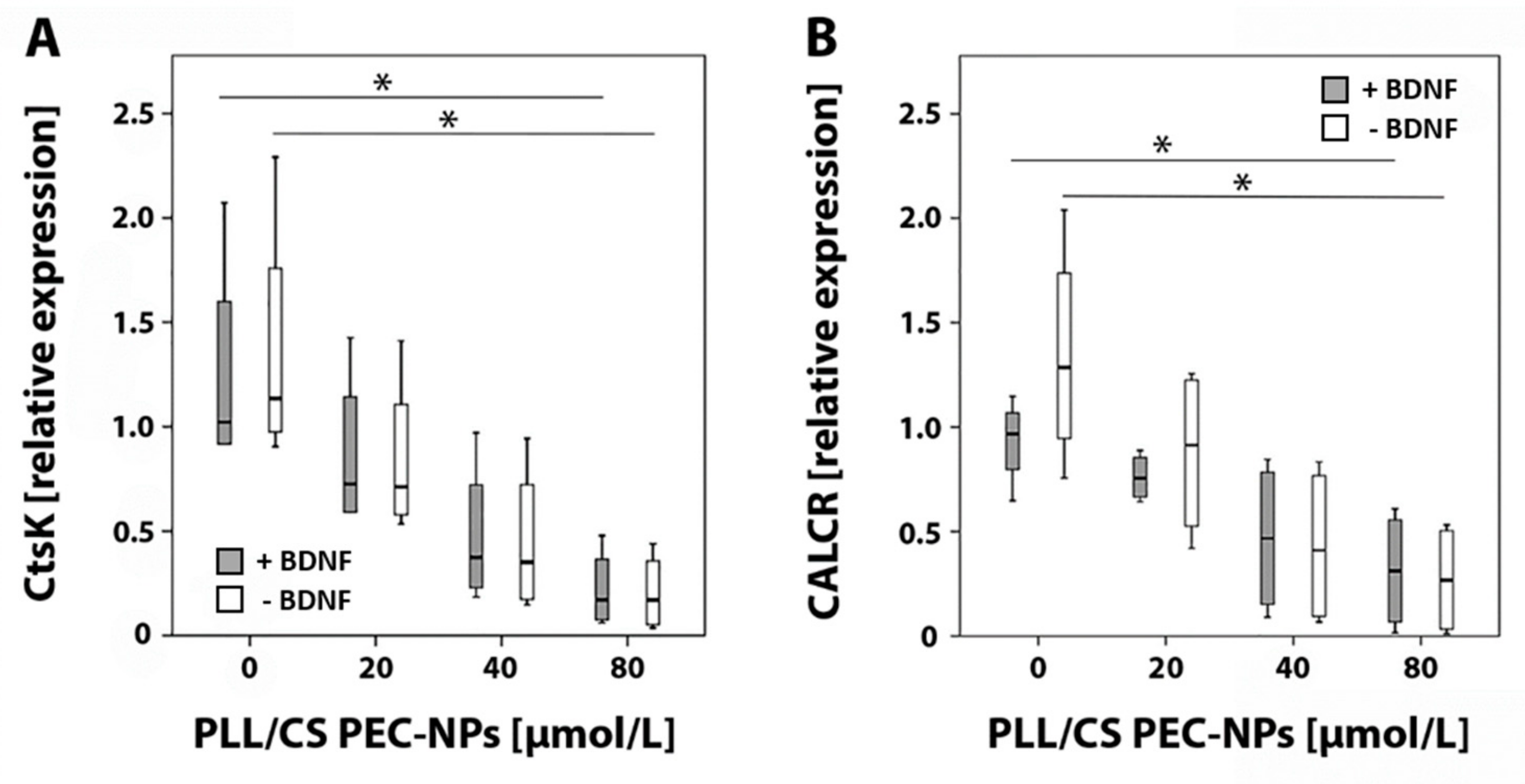
2.2. Detection of Cathepsin K and Calcitonin Receptor mRNA by Real-Time Reverse Transcriptase (RT)-Polymerase Chain Reaction (PCR)
2.3. Detection of Osteoclast Specific Tartrate-Resistant Acidic Phosphatase (TRAP) Isoform 5b
2.4. Pit Formation Assay
3. Discussion
4. Materials and Methods
4.1. Polyelectrolyte Complex Nanoparticles
4.2. In Vitro Culture of Osteoclasts
4.3. Live Cell Imaging
4.4. Real-Time Reverse Transcriptase (RT)-Polymerase Chain Reaction (PCR)
4.5. Tartrate-Resistant Acidic Phosphatase (TRAP) 5b Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Pit Formation Assay
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nesbitt, S.A.; Horton, M.A. Trafficking of matrix collagens through bone-resorbing osteoclasts. Science 1997, 276, 266–269. [Google Scholar] [CrossRef]
- Grabowski, P. Physiology of bone. Endocr. Dev. 2009, 16, 32–48. [Google Scholar] [PubMed]
- Florencio-Silva, R.; Sasso, G.R.; Sasso-Cerri, E.; Simoes, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed. Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [PubMed]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The osteocyte: An endocrine cell and more. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef] [PubMed]
- Sozen, T.; Ozisik, L.; Basaran, N.C. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Pino, A.M.; Rosen, C.J.; Rodriguez, J.P. In osteoporosis, differentiation of mesenchymal stem cells (MSCs) improves bone marrow adipogenesis. Biol. Res. 2012, 45, 279–287. [Google Scholar] [CrossRef]
- Infante, A.; Rodriguez, C.I. Osteogenesis and aging: Lessons from mesenchymal stem cells. Stem Cell Res. Ther. 2018, 9, 244. [Google Scholar] [CrossRef]
- Nikolaou, V.S.; Efstathopoulos, N.; Kontakis, G.; Kanakaris, N.K.; Giannoudis, P.V. The influence of osteoporosis in femoral fracture healing time. Injury 2009, 40, 663–668. [Google Scholar] [CrossRef]
- Cauley, J.A. Osteoporosis: Fracture epidemiology update 2016. Curr. Opin. Rheumatol. 2017, 29, 150–156. [Google Scholar] [CrossRef]
- Lorentzon, M.; Cummings, S.R. Osteoporosis: The evolution of a diagnosis. J. Intern. Med. 2015, 277, 650–661. [Google Scholar] [CrossRef]
- Agarwal, R.; Gonzalez-Garcia, C.; Torstrick, B.; Guldberg, R.E.; Salmeron-Sanchez, M.; Garcia, A.J. Simple coating with fibronectin fragment enhances stainless steel screw osseointegration in healthy and osteoporotic rats. Biomaterials 2015, 63, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginebra, M.P.; Canal, C.; Espanol, M.; Pastorino, D.; Montufar, E.B. Calcium phosphate cements as drug delivery materials. Adv. Drug Deliv. Rev. 2012, 64, 1090–1110. [Google Scholar] [CrossRef]
- Miron, R.J.; Zhang, Y.F. Osteoinduction: A review of old concepts with new standards. J. Dent. Res. 2012, 91, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, J.; Liu, J.; Wei, L.; Wu, G. BMP-functionalised coatings to promote osteogenesis for orthopaedic implants. Int. J. Mol. Sci. 2014, 15, 10150–10168. [Google Scholar] [CrossRef] [PubMed]
- Peres, J.A.; Lamano, T. Strategies for stimulation of new bone formation: A critical review. Braz. Dent. J. 2011, 22, 443–448. [Google Scholar] [CrossRef] [PubMed]
- De Witte, T.M.; Fratila-Apachitei, L.E.; Zadpoor, A.A.; Peppas, N.A. Bone tissue engineering via growth factor delivery: From scaffolds to complex matrices. Regen. Biomater. 2018, 5, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Roohani-Esfahani, S.I.; Zreiqat, H. Nanoparticles: A promising new therapeutic platform for bone regeneration? Nanomedicine 2017, 12, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Vehlow, D.; Schmidt, R.; Gebert, A.; Siebert, M.; Lips, K.S.; Muller, M. Polyelectrolyte Complex Based Interfacial Drug Delivery System with Controlled Loading and Improved Release Performance for Bone Therapeutics. Nanomaterials 2016, 6, 53. [Google Scholar] [CrossRef]
- Yavropoulou, M.P.; Yovos, J.G. Osteoclastogenesis--current knowledge and future perspectives. J. Musculoskelet Neuronal Int. 2008, 8, 204–216. [Google Scholar]
- Terreni, A.; Pezzati, P. Biochemical markers in the follow-up of the osteoporotic patients. Clin. Cases Miner Bone Metab. 2012, 9, 80–84. [Google Scholar]
- Pondel, M. Calcitonin and calcitonin receptors: Bone and beyond. Int. J. Exp. Pathol. 2000, 81, 405–422. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Goldring, S.R.; Lorenzo, J.A. Expression of the calcitonin receptor in bone marrow cell cultures and in bone: A specific marker of the differentiated osteoclast that is regulated by calcitonin. Endocrinology 1995, 136, 4572–4581. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.G.G.; Croucher, P.I.; Rogers, M.J. Bisphosphonates: Pharmacology, Mechanisms of Action and Clinical Uses. Osteoporos. Int. 1999, 9, S66–S80. [Google Scholar] [CrossRef] [PubMed]
- Binkley, N.; Bolognese, M.; Sidorowicz-Bialynicka, A.; Vally, T.; Trout, R.; Miller, C.; Buben, C.E.; Gilligan, J.P.; Krause, D.S. A phase 3 trial of the efficacy and safety of oral recombinant calcitonin: The oral calcitonin in postmenopausal osteoporosis (ORACAL) trial. J. Bone Miner. Res. 2012, 27, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Burkiewicz, J.S.; Scarpace, S.L.; Bruce, S.P. Denosumab in Osteoporosis and Oncology. Ann. Pharmacother. 2009, 43, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Kermani, P.; Hempstead, B. Brain-derived neurotrophic factor: A newly described mediator of angiogenesis. Trends Cardiovasc. Med. 2007, 17, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, T.; Fukunaga, T.; Yamashita, K.; Kobashi, N.; Takano-Yamamoto, T. Gene and protein expression of brain-derived neurotrophic factor and TrkB in bone and cartilage. Bone 2001, 28, 404–409. [Google Scholar] [CrossRef]
- Asaumi, K.; Nakanishi, T.; Asahara, H.; Inoue, H.; Takigawa, M. Expression of neurotrophins and their receptors (TRK) during fracture healing. Bone 2000, 26, 625–633. [Google Scholar] [CrossRef]
- Kilian, O.; Hartmann, S.; Dongowski, N.; Karnati, S.; Baumgart-Vogt, E.; Hartel, F.V.; Noll, T.; Schnettler, R.; Lips, K.S. BDNF and its TrkB receptor in human fracture healing. Ann. Anat. 2014, 196, 286–295. [Google Scholar] [CrossRef]
- Kauschke, V.; Gebert, A.; Calin, M.; Eckert, J.; Scheich, S.; Heiss, C.; Lips, K.S. Effects of new beta-type Ti-40Nb implant materials, brain-derived neurotrophic factor, acetylcholine and nicotine on human mesenchymal stem cells of osteoporotic and non osteoporotic donors. PLoS ONE 2018, 13, e0193468. [Google Scholar] [CrossRef]
- Kauschke, V.; Schneider, M.; Jauch, A.; Schumacher, M.; Kampschulte, M.; Rohnke, M.; Henss, A.; Bamberg, C.; Trinkaus, K.; Gelinsky, M.; et al. Effects of a Pasty Bone Cement Containing Brain-Derived Neurotrophic Factor-Functionalized Mesoporous Bioactive Glass Particles on Metaphyseal Healing in a New Murine Osteoporotic Fracture Model. Int. J. Mol. Sci. 2018, 19, 3531. [Google Scholar] [CrossRef] [PubMed]
- Ai, L.S.; Sun, C.Y.; Wang, Y.D.; Zhang, L.; Chu, Z.B.; Qin, Y.; Gao, F.; Yan, H.; Guo, T.; Chen, L.; et al. Gene silencing of the BDNF/TrkB axis in multiple myeloma blocks bone destruction and tumor burden in vitro and in vivo. Int. J. Cancer 2013, 133, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.Y.; Chu, Z.B.; She, X.M.; Zhang, L.; Chen, L.; Ai, L.S.; Hu, Y. Brain-derived neurotrophic factor is a potential osteoclast stimulating factor in multiple myeloma. Int. J. Cancer 2012, 130, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Ternes, S.; Trinkaus, K.; Bergen, I.; Knaack, S.; Gelinsky, M.; Kilian, O.; Heiss, C.; Lips, K.S. Impact of acetylcholine and nicotine on human osteoclastogenesis in vitro. Int. Immunopharmacol. 2015, 29, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, S.; Kaji, K.; Kudo, A. Identification and characterization of the new osteoclast progenitor with macrophage phenotypes being able to differentiate into mature osteoclasts. J. Bone Min. Res. 2000, 15, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Russmueller, G.; Winkler, L.; Lieber, R.; Seemann, R.; Pirklbauer, K.; Perisanidis, C.; Kapeller, B.; Spassova, E.; Halwax, E.; Poeschl, W.P.; et al. In vitro effects of particulate bone substitute materials on the resorption activity of human osteoclasts. Eur. Cell Mater. 2017, 34, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Sabokbar, A.; Quinn, J.M.; McGee, J.O.; Athanasou, N.A.; Fujikawa, Y. The human osteoclast precursor circulates in the monocyte fraction. Endocrinology 1996, 137, 4058–4060. [Google Scholar]
- Abdallah, D.; Jourdain, M.L.; Braux, J.; Guillaume, C.; Gangloff, S.C.; Jacquot, J.; Velard, F. An Optimized Method to Generate Human Active Osteoclasts from Peripheral Blood Monocytes. Front. Immunol. 2018, 9, 632. [Google Scholar] [CrossRef]
- Brulefert, K.; Cordova, L.A.; Brulin, B.; Faucon, A.; Hulin, P.; Nedellec, S.; Gouin, F.; Passuti, N.; Ishow, E.; Heymann, D. Pro-osteoclastic in vitro effect of Polyethylene-like nanoparticles: Involvement in the pathogenesis of implant aseptic loosening. J. Biomed. Mater Res. A. 2016, 104, 2649–2657. [Google Scholar] [CrossRef]
- Zaidi, M.; Troen, B.; Moonga, B.S.; Abe, E. Cathepsin K, osteoclastic resorption, and osteoporosis therapy. J. Bone Min. Res. 2001, 16, 1747–1749. [Google Scholar] [CrossRef]
- Drake, M.T.; Clarke, B.L.; Oursler, M.J.; Khosla, S. Cathepsin K Inhibitors for Osteoporosis: Biology, Potential Clinical Utility, and Lessons Learned. Endocr. Rev. 2017, 38, 325–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, M.; Inzerillo, A.M.; Moonga, B.S.; Bevis, P.J.; Huang, C.L. Forty years of calcitonin—Where are we now? A tribute to the work of Iain Macintyre. FRS Bone 2002, 30, 655–663. [Google Scholar]
- Kallio, D.M.; Garant, P.R.; Minkin, C. Ultrastructural effects of calcitonin on osteoclasts in tissue culture. J. Ultrastruct. Res. 1972, 39, 205–216. [Google Scholar] [CrossRef]
- Chambers, T.J.; Magnus, C.J. Calcitonin alters behaviour of isolated osteoclasts. J. Pathol. 1982, 136, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Goldring, S.; Katz, M.; Hilsenbeck, S.; Williams, R.; Roodman, G.D. Downregulation of calcitonin receptor mRNA expression by calcitonin during human osteoclast-like cell differentiation. J. Clin. Invest 1995, 95, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Overman, R.A.; Borse, M.; Gourlay, M.L. Salmon calcitonin use and associated cancer risk. Ann. Pharmacother. 2013, 47, 1675–1684. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, K.; Lu, X.; Li, M.; Liu, H.; Xie, C.; Meng, F.; Jiang, O.; Li, C.; Zhi, W. BMP-2 encapsulated polysaccharide nanoparticle modified biphasic calcium phosphate scaffolds for bone tissue regeneration. J. Biomed. Mater Res. A 2015, 103, 1520–1532. [Google Scholar] [CrossRef]
- Nguyen, V.; Meyers, C.A.; Yan, N.; Agarwal, S.; Levi, B.; James, A.W. BMP-2-induced bone formation and neural inflammation. J. Orthop. 2017, 14, 252–256. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds poly(l-lysine) (PLL) and cellulose sulfate (CS-0.5, CS-3.0) for PEC-NPs used in this report are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Length [bp] | Accession No. |
|---|---|---|---|
| Cathepsin K | for GCG ATA ATC TGA ACC ATG CG rev TTG TTT CCC CAG TTT TCT CCC | 103 | NM_000396.3 |
| Calcitonin Receptor | for TGA GTG TGG AAA CCC ATT TGC rev ATT TTG GTC ACA AGC ACC CG | 109 | NM_001164737.1 |
| Beta-2-microglubulin | for TCT CTC TTT CTG GCC TGG AG rev CAA CTT CAA TGT CGG ATG GA | 135 | NM_004048.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kauschke, V.; Hessland, F.M.; Vehlow, D.; Müller, M.; Heiss, C.; Lips, K.S. High Concentrations of Polyelectrolyte Complex Nanoparticles Decrease Activity of Osteoclasts. Molecules 2019, 24, 2346. https://doi.org/10.3390/molecules24122346
Kauschke V, Hessland FM, Vehlow D, Müller M, Heiss C, Lips KS. High Concentrations of Polyelectrolyte Complex Nanoparticles Decrease Activity of Osteoclasts. Molecules. 2019; 24(12):2346. https://doi.org/10.3390/molecules24122346
Chicago/Turabian StyleKauschke, Vivien, Felix Maximilian Hessland, David Vehlow, Martin Müller, Christian Heiss, and Katrin Susanne Lips. 2019. "High Concentrations of Polyelectrolyte Complex Nanoparticles Decrease Activity of Osteoclasts" Molecules 24, no. 12: 2346. https://doi.org/10.3390/molecules24122346
APA StyleKauschke, V., Hessland, F. M., Vehlow, D., Müller, M., Heiss, C., & Lips, K. S. (2019). High Concentrations of Polyelectrolyte Complex Nanoparticles Decrease Activity of Osteoclasts. Molecules, 24(12), 2346. https://doi.org/10.3390/molecules24122346