
Molecular Characterization and Heterologous Production of the Bacteriocin Peocin, a DNA Starvation/Stationary Phase Protection Protein, from Paenibacillus ehimensis NPUST1

 ,
, 
Abstract
:1. Introduction
2. Results
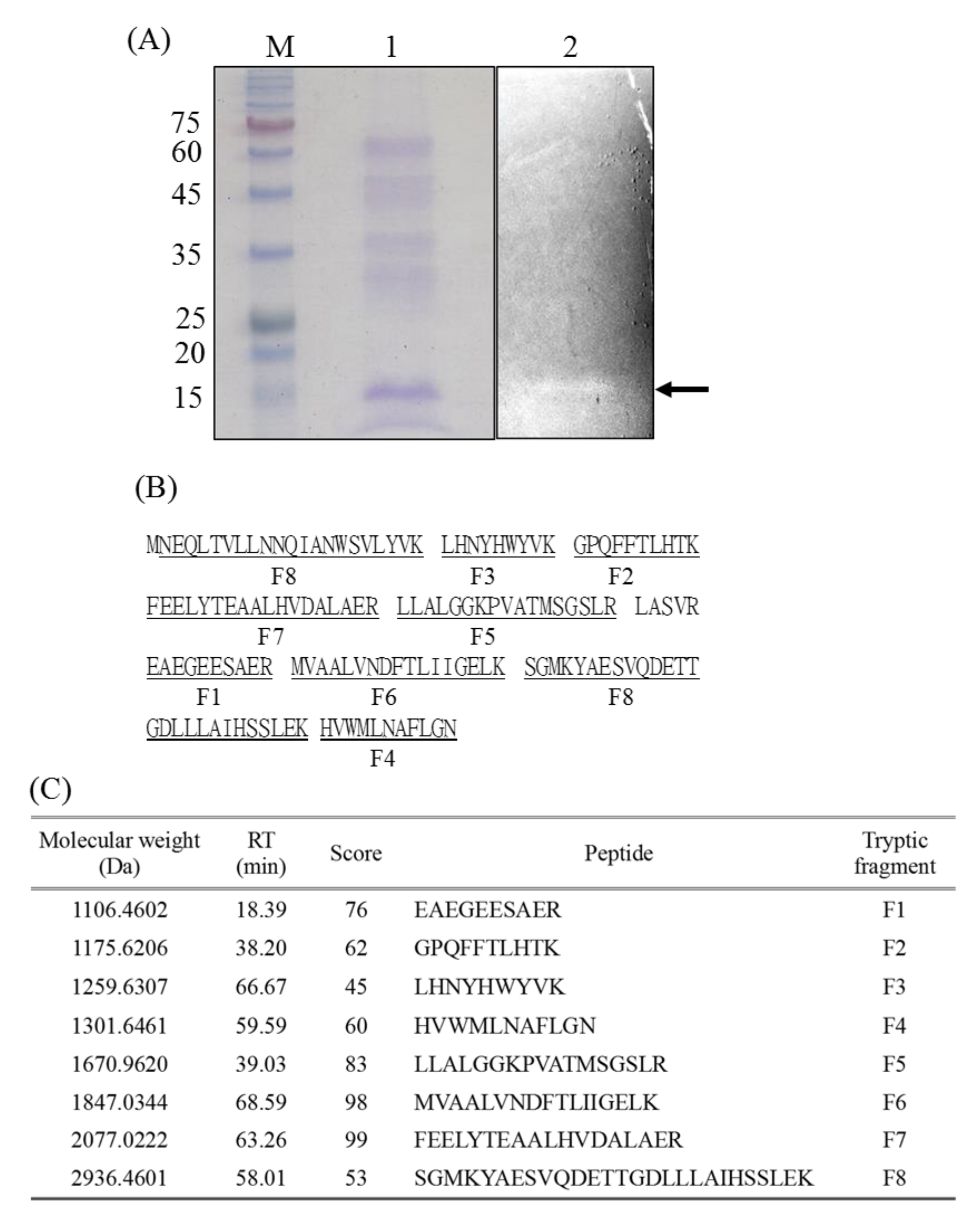
2.1. Characterization of the Bacteriocin-Like Substance from P. ehimensis NPUST1
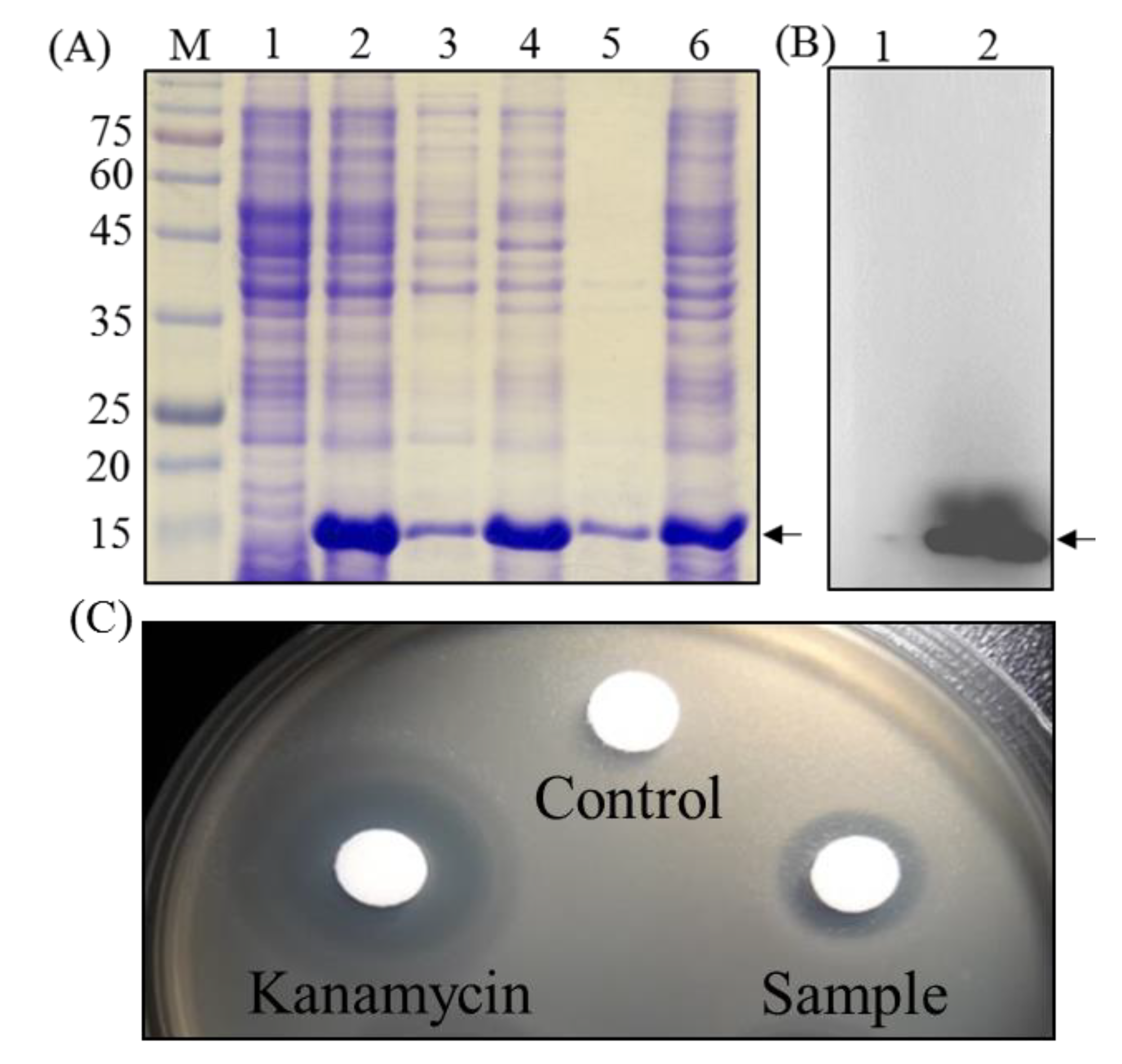
2.2. Expression and Characterization of the Recombinant Peocin by Flask Cultivation
2.3. Fermentation of Recombinant E. coli by Fed-Batch Cultivation
2.4. Purification and Characterization of Recombinant Peocin
2.5. Biological Activity of Purified Peocin
3. Discussion
4. Materials and Methods
4.1. Fish and Bacterial Strain
4.2. Gel Overlay and Mass Spectrometry Analysis
4.3. Expression Plasmid and Flask Cultivation
4.4. Bioreactor and Fed-Batch Cultivation
4.5. Purification of Recombinant Peocin Protein
4.6. Molecular Mass Determination
4.7. MIC and MBC Determination
4.8. Challenge Test
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Grenni, P.; Ancona, V.; Caracciolo, A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Habboush, Y.; Guzman, N. Antibiotic Resistance. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Watkins, R.R.; Bonomo, R.A. Overview: Global and Local Impact of Antibiotic Resistance. Infect. Dis. Clin. N. Am. 2016, 30, 313–322. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef]
- Ahmad, V.; Khan, M.S.; Jamal, Q.M.S.; Alzohairy, M.A.; Al Karaawi, M.A.; Siddiqui, M.U. Antimicrobial potential of bacteriocins: in therapy, agriculture and food preservation. Int. J. Antimicrob. Agents 2017, 49, 1–11. [Google Scholar] [CrossRef]
- Ben Lagha, A.; Haas, B.; Gottschalk, M.; Grenier, D. Antimicrobial potential of bacteriocins in poultry and swine production. Vet. Res. 2017, 48, 22. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Ross, R.P.; Stanton, C.; Silva, C.C.G. Characterization and Application of Antilisterial Enterocins on Model Fresh Cheese. J. Food Prot. 2017, 80, 1303–1316. [Google Scholar] [CrossRef]
- Kondrotiene, K.; Kasnauskyte, N.; Serniene, L.; Gölz, G.; Alter, T.; Kaskoniene, V.; Maruska, A.S.; Malakauskas, M.; Mulkyte, K. Characterization and application of newly isolated nisin producing Lactococcus lactis strains for control of Listeria monocytogenes growth in fresh cheese. LWT 2018, 87, 507–514. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.; Rehman, M.U.; Mehmood, K.; Jiang, X.; Iqbal, M.; Tong, X.; Gao, X.; Li, J. Antibacterial activity of Lactobacillus plantarum isolated from Tibetan yaks. Microb. Pathog. 2018, 115, 293–298. [Google Scholar] [CrossRef]
- Caly, D.L.; D’Inca, R.; Auclair, E.; Drider, D. Alternatives to antibiotics to prevent necrotic enteritis in broiler chickens: A microbiologist’s perspective. Front. Microbiol. 2015, 6, 1336. [Google Scholar] [CrossRef]
- Azhar, N.S.; Zin, N.H.M.; Hamid, T.H.T.A. Lactococcus lactis Strain A5 Producing Nisin-like Bacteriocin Active against Gram Positive and Negative Bacteria. Trop. Life Sci. Res. 2017, 28, 107–118. [Google Scholar] [CrossRef]
- Shokri, D.; Zaghian, S.; Khodabakhsh, F.; Fazeli, H.; Mobasherizadeh, S.; Ataei, B. Antimicrobial activity of a UV-stable bacteriocin-like inhibitory substance (BLIS) produced by Enterococcus faecium strain DSH20 against vancomycin-resistant Enterococcus (VRE) strains. J. Microbiol. Immunol. Infect. 2014, 47, 371–376. [Google Scholar] [CrossRef]
- Sahoo, T.K.; Jena, P.K.; Patel, A.K.; Seshadri, S. Purification and Molecular Characterization of the Novel Highly Potent Bacteriocin TSU4 Produced by Lactobacillus animalis TSU4. Appl. Biochem. Biotechnol. 2015, 177, 90–104. [Google Scholar] [CrossRef]
- Wannun, P.; Piwat, S.; Teanpaisan, R. Purification and characterization of bacteriocin produced by oral Lactobacillus paracasei SD1. Anaerobe 2014, 27, 17–21. [Google Scholar] [CrossRef]
- Mesa-Pereira, B.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. Heterologous Expression of Biopreservative Bacteriocins With a View to Low Cost Production. Front. Microbiol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Jin, X.; Yao, J.; Fan, H.; Che, Y.; Pan, J.; Zhang, L.; Pan, X.; Gelbič, I.; Huang, T.; Guan, X. Heterologous expression and purification of BtCspB, a novel cold-shock protein-like bacteriocin from Bacillus thuringiensis BRC-ZYR2. World J. Microbiol. Biotechnol. 2019, 35, 23. [Google Scholar] [CrossRef]
- Chen, S.-W.; Liu, C.-H.; Hu, S.-Y. Dietary administration of probiotic Paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 695–703. [Google Scholar] [CrossRef]
- Zhao, G.; Ceci, P.; Ilari, A.; Giangiacomo, L.; Laue, T.M.; Chiancone, E.; Chasteen, N.D. Iron and Hydrogen Peroxide Detoxification Properties of DNA-binding Protein from Starved Cells. A Ferritin-Like DNA-Binding Protein of Escherichia coli. J. Boil. Chem. 2002, 277, 27689–27696. [Google Scholar] [CrossRef]
- Chiancone, E.; Ceci, P. The multifaceted capacity of Dps proteins to combat bacterial stress conditions: Detoxification of iron and hydrogen peroxide and DNA binding. Biochim. Biophys. Acta (BBA) Gen. Subj. 2010, 1800, 798–805. [Google Scholar] [CrossRef]
- Karas, V.O.; Westerlaken, I.; Meyer, A.S. The DNA-Binding Protein from Starved Cells (Dps) Utilizes Dual Functions To Defend Cells against Multiple Stresses. J. Bacteriol. 2015, 197, 3206–3215. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.T.; Koo, B.-K.; Vu, T.T.T.; Song, J.-A.; Chong, S.-H.; Jeong, B.; Ryu, H.-B.; Moh, S.-H.; Choe, H. Prokaryotic Soluble Overexpression and Purification of Bioactive Human Growth Hormone by Fusion to Thioredoxin, Maltose Binding Protein, and Protein Disulfide Isomerase. PLoS ONE 2014, 9, e89038. [Google Scholar] [CrossRef]
- Kim, M.-J.; Park, H.S.; Seo, K.H.; Yang, H.-J.; Kim, S.-K.; Choi, J.-H. Complete Solubilization and Purification of Recombinant Human Growth Hormone Produced in Escherichia coli. PLoS ONE 2013, 8, e56168. [Google Scholar] [CrossRef]
- Hu, S.-Y.; Wu, J.-L.; Huang, J.-H. Production of tilapia insulin-like growth factor-2 in high cell density cultures of recombinant Escherichia coli. J. Biotechnol. 2004, 107, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Shiloach, J.; Fass, R. Growing E. coli to high cell density—A historical perspective on method development. Biotechnol. Adv. 2005, 23, 345–357. [Google Scholar] [CrossRef]
- Tanhaeian, A.; Damavandi, M.S.; Mansury, D.; Ghaznini, K. Expression in eukaryotic cells and purification of synthetic gene encoding enterocin P: a bacteriocin with broad antimicrobial spectrum. AMB Express 2019, 9, 6. [Google Scholar] [CrossRef]
- Jiang, H.; Li, P.; Gu, Q. Heterologous expression and purification of plantaricin NC8, a two-peptide bacteriocin against Salmonella spp. from Lactobacillus plantarum ZJ316. Protein Expr. Purif. 2016, 127, 28–34. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.; Sood, S.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Arthur, T.D.; Cavera, V.L.; Chikindas, M.L. On bacteriocin delivery systems and potential. Future Microbial. 2014, 9, 235–248. [Google Scholar] [CrossRef]
- Wiley, D.; Redfield, S.; Zon, L. Chemical screening in zebrafish for novel biological and therapeutic discovery. Methods Cell Boil. 2017, 138, 651–679. [Google Scholar] [CrossRef]
- Wang, L.; Chan, J.Y.; Rego, J.V.; Chong, C.-M.; Ai, N.; Falcão, C.B.; Rádis-Baptista, G.; Lee, S.M. Rhodamine B-conjugated encrypted vipericidin nonapeptide is a potent toxin to zebrafish and associated with in vitro cytotoxicity. Biochim. Biophys. Acta (BBA) Gen. Subj. 2015, 1850, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, S.; Konrad, D.; Calvez, S.; Drider, D.; Prévost, H.; Lacroix, C. Production of recombinant bacteriocin divercin V41 by high cell density Escherichia coli batch and fed-batch cultures. Appl. Microbiol. Biotechnol. 2007, 77, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; Criado, R.; Citti, R.; Martin, M.; Herranz, C.; Nes, I.; Cintas, L.; Hernandez, P.E. Cloning, production and functional expression of enterocin P, a sec-dependent bacteriocin produced by Enterococcus faecium P13, in Escherichia coli. Int. J. Food Microbiol. 2005, 103, 239–250. [Google Scholar] [CrossRef]
- Beaulieu, L.; Tolkatchev, D.; Jetté, J.-F.; Groleau, D.; Subirade, M. Production of active pediocin PA-1 in Escherichia coli using a thioredoxin gene fusion expression approach: Cloning, expression, purification, and characterization. Can. J. Microbiol. 2007, 53, 1246–1258. [Google Scholar] [CrossRef]
- Gibbs, G.M.; Davidson, B.E.; Hillier, A.J. Novel Expression System for Large-Scale Production and Purification of Recombinant Class IIa Bacteriocins and Its Application to Piscicolin 126. Appl. Environ. Microbiol. 2004, 70, 3292–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, G.; Srivastava, S. Scaling Up the Production of Recombinant Antimicrobial Plantaricin E from a Heterologous Host, Escherichia coli. Probiotics Antimicrob. Proteins 2015, 7, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Nigutová, K.; Serenčová, L.; Piknova, M.; Javorský, P.; Pristaš, P. Heterologous expression of functionally active enterolysin A, class III bacteriocin from Enterococcus faecalis, in Escherichia coli. Protein Expr. Purif. 2008, 60, 20–24. [Google Scholar] [CrossRef]
- Chung, W.-J.; Huang, C.-L.; Gong, H.-Y.; Ou, T.-Y.; Hsu, J.-L.; Hu, S.-Y. Recombinant production of biologically active giant grouper (Epinephelus lanceolatus) growth hormone from inclusion bodies of Escherichia coli by fed-batch culture. Protein Expr. Purif. 2015, 110, 79–88. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Faikoh, E.N.; Hong, Y.-H.; Hu, S.-Y. Liposome-encapsulated cinnamaldehyde enhances zebrafish (Danio rerio) immunity and survival when challenged with Vibrio vulnificus and Streptococcus agalactiae. Fish Shellfish Immunol. 2014, 38, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-S.; Saputra, F.; Chen, Y.-C.; Hu, S.-Y. Dietary administration of Bacillus amyloliquefaciens R8 reduces hepatic oxidative stress and enhances nutrient metabolism and immunity against Aeromonas hydrophila and Streptococcus agalactiae in zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 86, 410–419. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the E. coli BL21(DE3) for peocin expression are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Volume (mL) | Protein Conc. (mg/mL) | Amount of Total Protein (mg) | Amount of Peocin (mg) | Peocin Recovery (%) | Purity (%) |
|---|---|---|---|---|---|---|
| Supernatant of cell lysate a | 20 | 6.82 | 136.4 | 91.8 | 100 | 67.3 |
| Ni-NTA affinity chromatography | 15 | 4.28 | 64.3 | 59.4 | 64.8 | 92.3 b |
| Dialysis | 24 | 2.05 | 49.2 | 45.3 | 49.3 | 92.1 c |
| Pathogen | Purified Peocin (µg/mL) | Kanamycin (µg/mL) | Pathogen | Purified Peocin (µg/mL) | Kanamycin (µg/mL) | ||||
|---|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | ||
| Aeromonas hydrophila | 20 | 40 | 0.5 | 2.0 | Staphylococcus aureus | 20 | 50 | 0.5 | 2 |
| Streptococcus agalactise | 30 | 50 | 2.0 | 4.0 | Salmonella typhimurium | 30 | 60 | 1 | 2.0 |
| Streptococcus iniae | 30 | 60 | 3.0 | 6.0 | Listeria monocytogenes | 40 | 60 | 1 | 3.0 |
| Vibrio vulnificus a | 30 | 40 | Non b | Non b | Methicillin-resistant S. aureus (MRSA) a | Non b | Non b | Non b | Non b |
| Vibrio parahaemolyticus a | 30 | 40 | Non b | Non b | Pseudomonas aeruginosa | 40 | 60 | 2.0 | 4.0 |
| Vibrio alginolyticus | 30 | 50 | 3.0 | 6.0 | Porphyromonas gingivalis | 30 | 60 | 2.0 | 3.0 |
| Debaryomyces hansenii | 50 | 80 | Non b | Non b | Prevotella nigrescens | 20 | 50 | 1 | 3.0 |
| Escherichia coli | 20 | 30 | 1.0 | 2.0 | Propionbacterium acnes | 20 | 40 | 1 | 2.0 |
| Bacteriocin | Native Host | E. coli Strain | Vector | Culture Type | Bacteriocin Yield | References |
|---|---|---|---|---|---|---|
| Divercin 41 Class(IIa) | C. divergens V41 | Origami (DE3) | pET32b | Fed-batch | 74 mg/L | [33] |
| Enterocin P (Class IIa) | Enterococcus faecium P13 | Tuner (DE3) | pETBlue-1 | Shake-flask | 0.006 mg/L | [34] |
| Pediocin PA-1 (Class IIa) | P. acidilactici PAC1.0 | Origami (DE3) | pET-32b | Shake-flask | 30 mg/L | [35] |
| Piscicolin 126 (Class IIa) | C. piscicola JG126 | AD494(DE3) | pET-32b | Fed-batch | 26 mg/L | [36] |
| Plantaricin E (Class IIb) | L. plantarum | BL21 (DE3) | pET32a | Shake-flask | 15 mg/L | [37] |
| BtCspB | B. thuringiensis BRC-ZYR2 | BL21 (DE3) | pET32a | Shake-flask | 20 mg/L | [18] |
| Enterolysin A (Class III) | E. faecalis | SG13009[pREP4] | pQE-30 UA | Shake-flask | 1.69 mg/g of wet cells | [38] |
| Peocin (Class III) | P. ehimensis | BL21 (DE3) | pET28b | Fed-batch | 2260 mg/L | This work |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, C.-C.; Murni, L.; Han, T.-W.; Arfiati, D.; Shih, H.-T.; Hu, S.-Y. Molecular Characterization and Heterologous Production of the Bacteriocin Peocin, a DNA Starvation/Stationary Phase Protection Protein, from Paenibacillus ehimensis NPUST1. Molecules 2019, 24, 2516. https://doi.org/10.3390/molecules24132516
Tseng C-C, Murni L, Han T-W, Arfiati D, Shih H-T, Hu S-Y. Molecular Characterization and Heterologous Production of the Bacteriocin Peocin, a DNA Starvation/Stationary Phase Protection Protein, from Paenibacillus ehimensis NPUST1. Molecules. 2019; 24(13):2516. https://doi.org/10.3390/molecules24132516
Chicago/Turabian StyleTseng, Chung-Chih, Lini Murni, Tai-Wei Han, Diana Arfiati, Hui-Tsu Shih, and Shao-Yang Hu. 2019. "Molecular Characterization and Heterologous Production of the Bacteriocin Peocin, a DNA Starvation/Stationary Phase Protection Protein, from Paenibacillus ehimensis NPUST1" Molecules 24, no. 13: 2516. https://doi.org/10.3390/molecules24132516
APA StyleTseng, C. -C., Murni, L., Han, T. -W., Arfiati, D., Shih, H. -T., & Hu, S. -Y. (2019). Molecular Characterization and Heterologous Production of the Bacteriocin Peocin, a DNA Starvation/Stationary Phase Protection Protein, from Paenibacillus ehimensis NPUST1. Molecules, 24(13), 2516. https://doi.org/10.3390/molecules24132516