
Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide


 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Method
2.1. Compounds and Peptide
2.2. Cell Lines
2.3. MTT Cell Viability Assay for the Individual Compounds
2.4. MTT Cell Viability Assay for the Co-Administration
2.5. Dual-Fluorescence for Live/Dead Nucleated Cell Assay
2.6. Molecular Docking
2.7. Molecular Dynamics Simulation
2.8. Statistical Analysis
3. Results
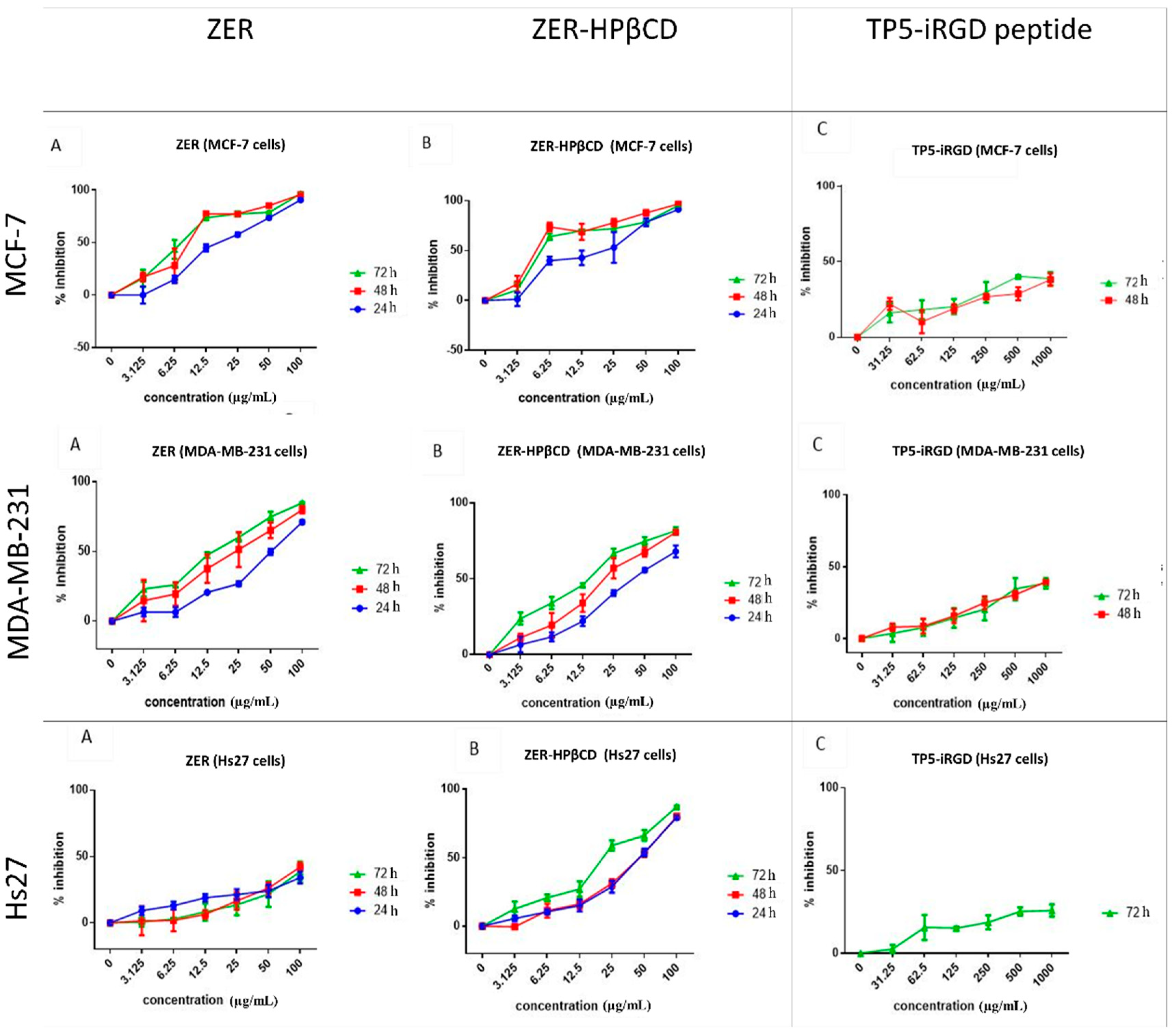
3.1. MTT Assay of Individual Compounds
3.2. MTT Assay of the Co-Administration
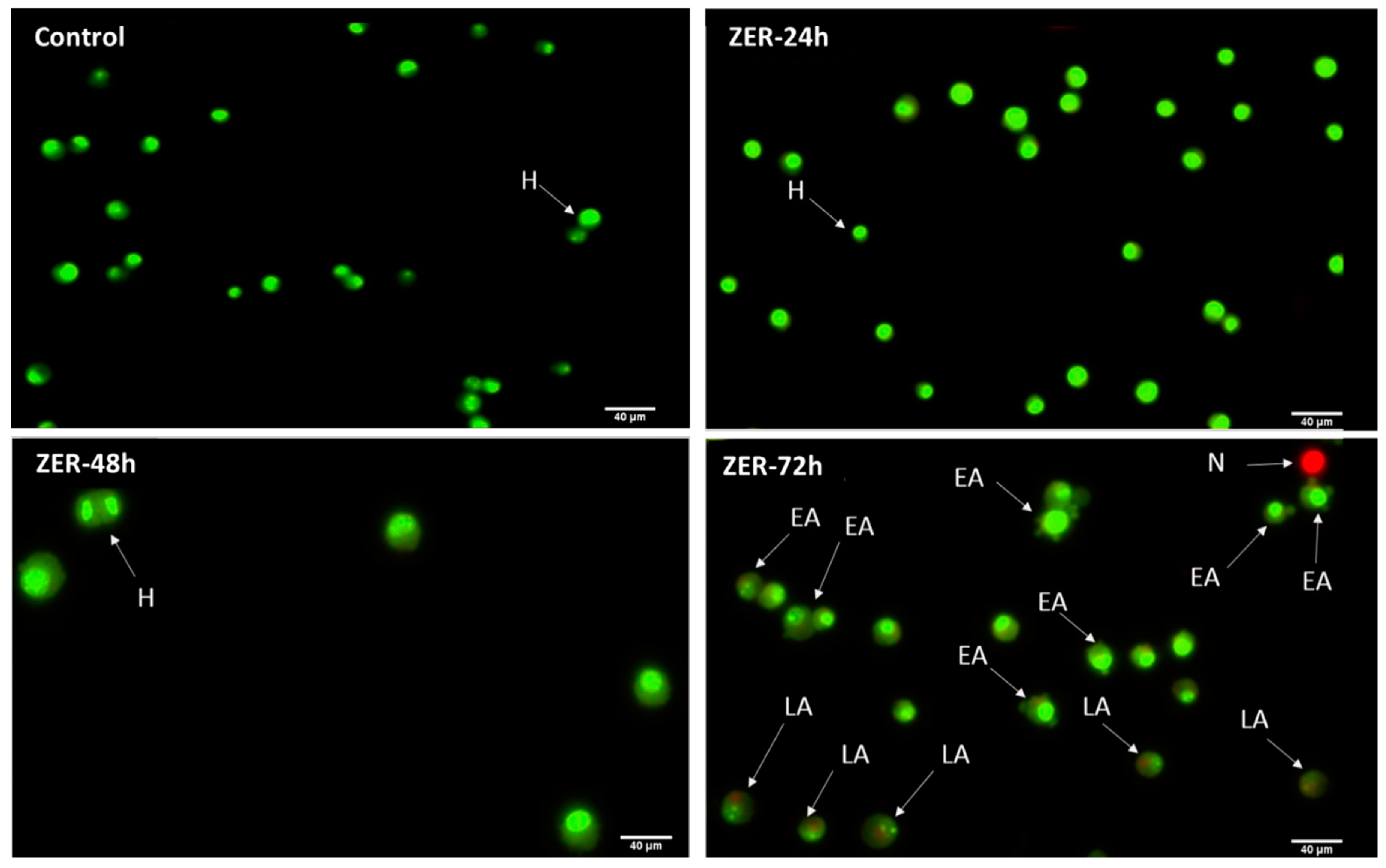
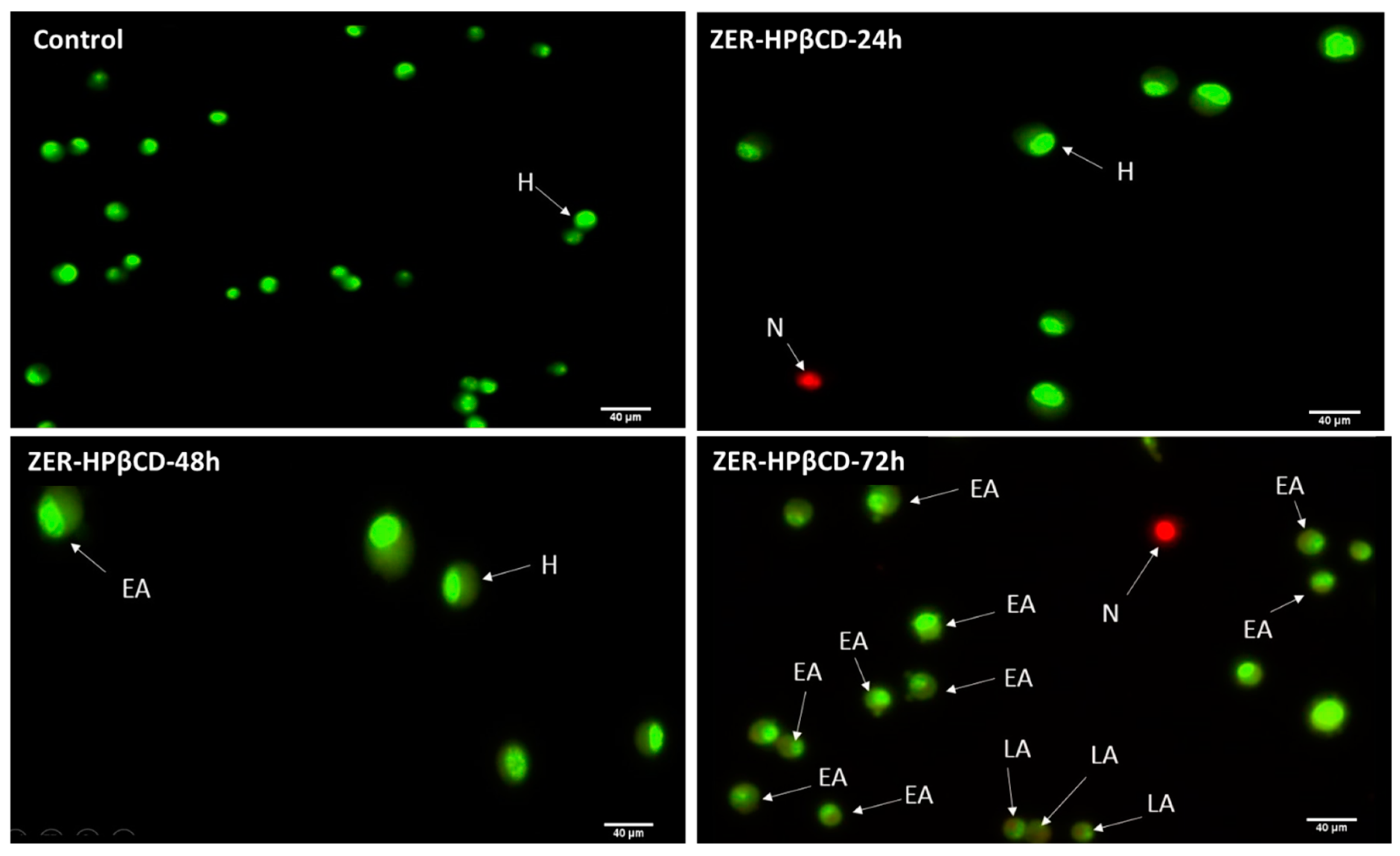
3.3. Apoptosis Detection by Dual Staining
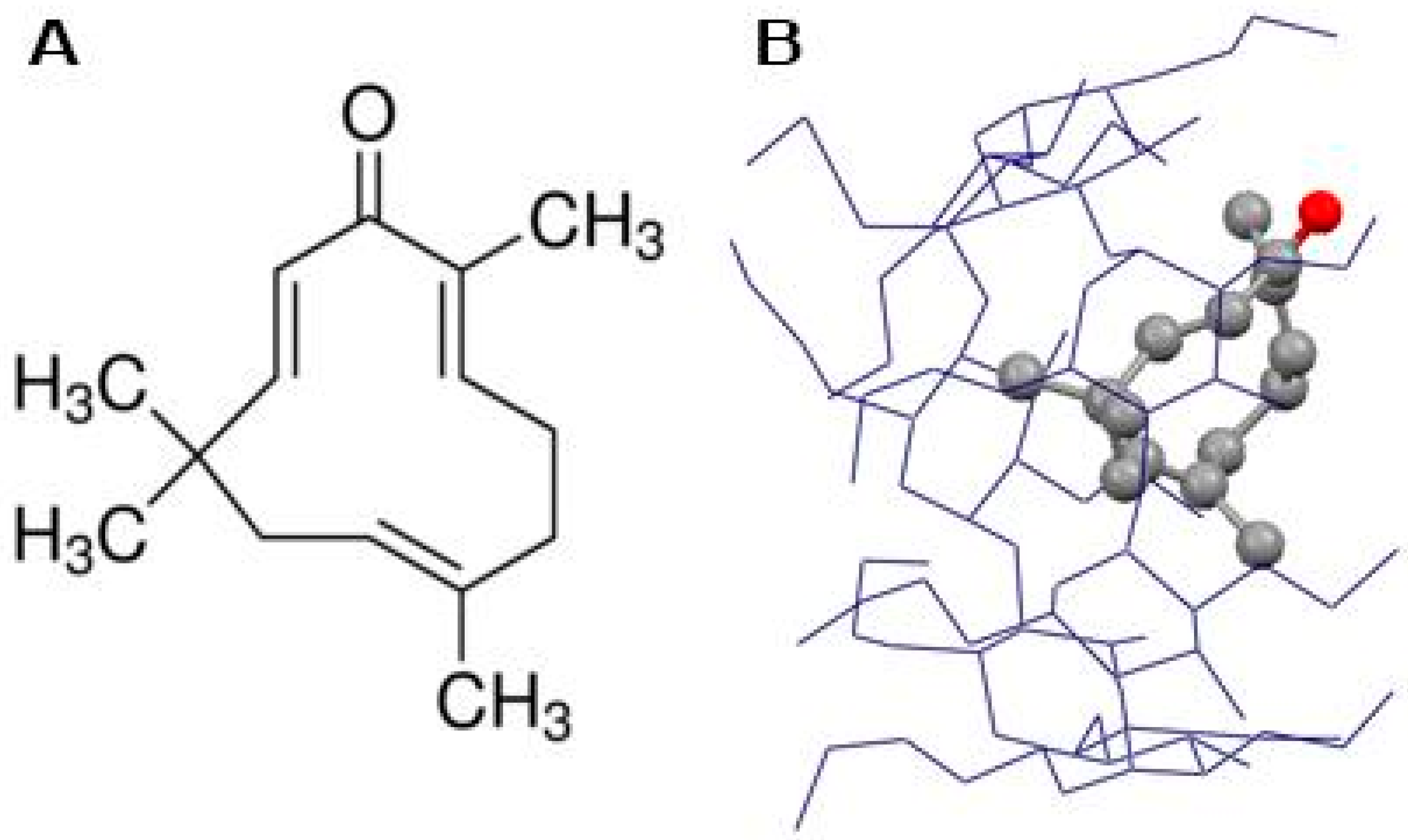
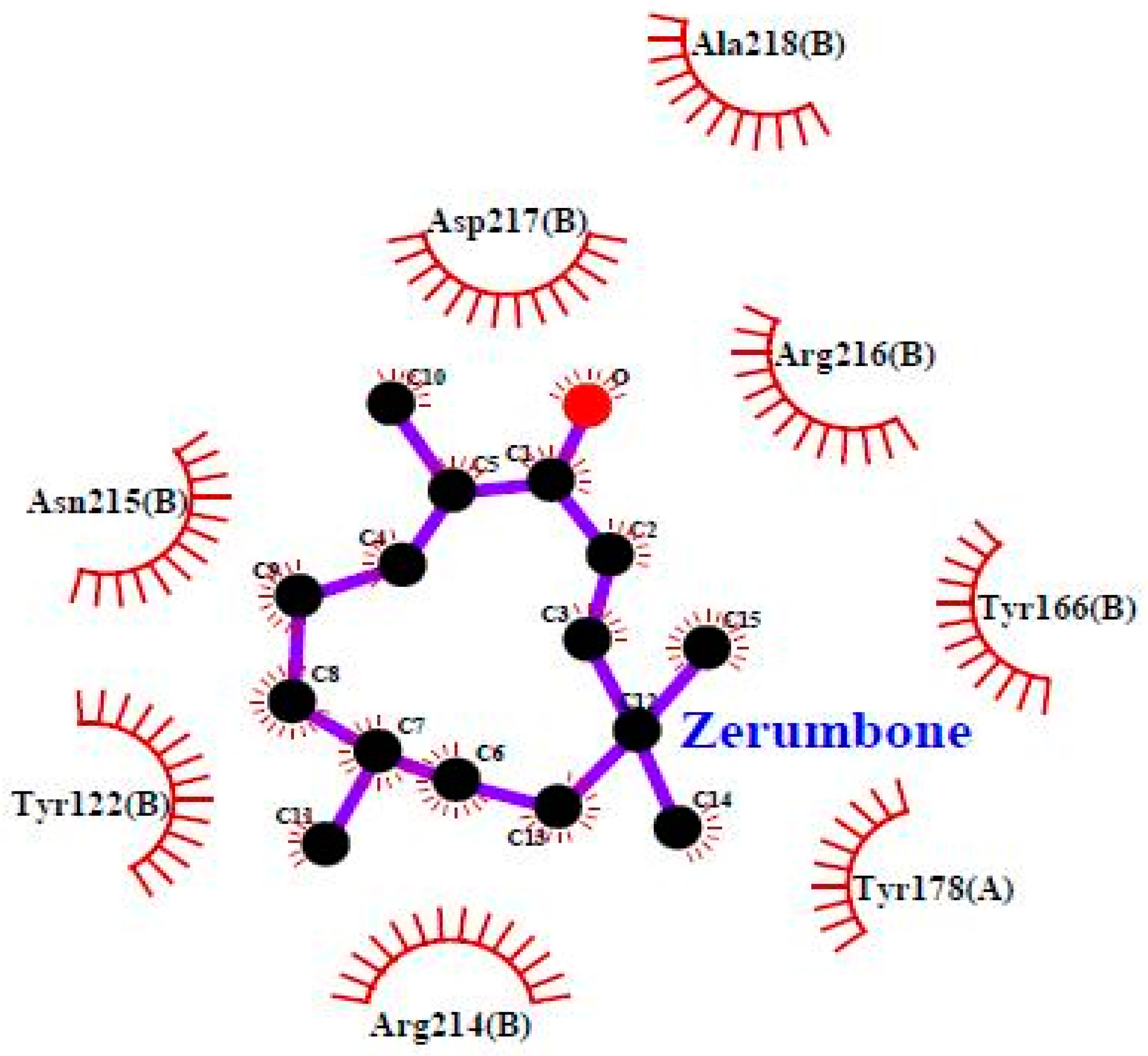
3.4. Molecular Docking
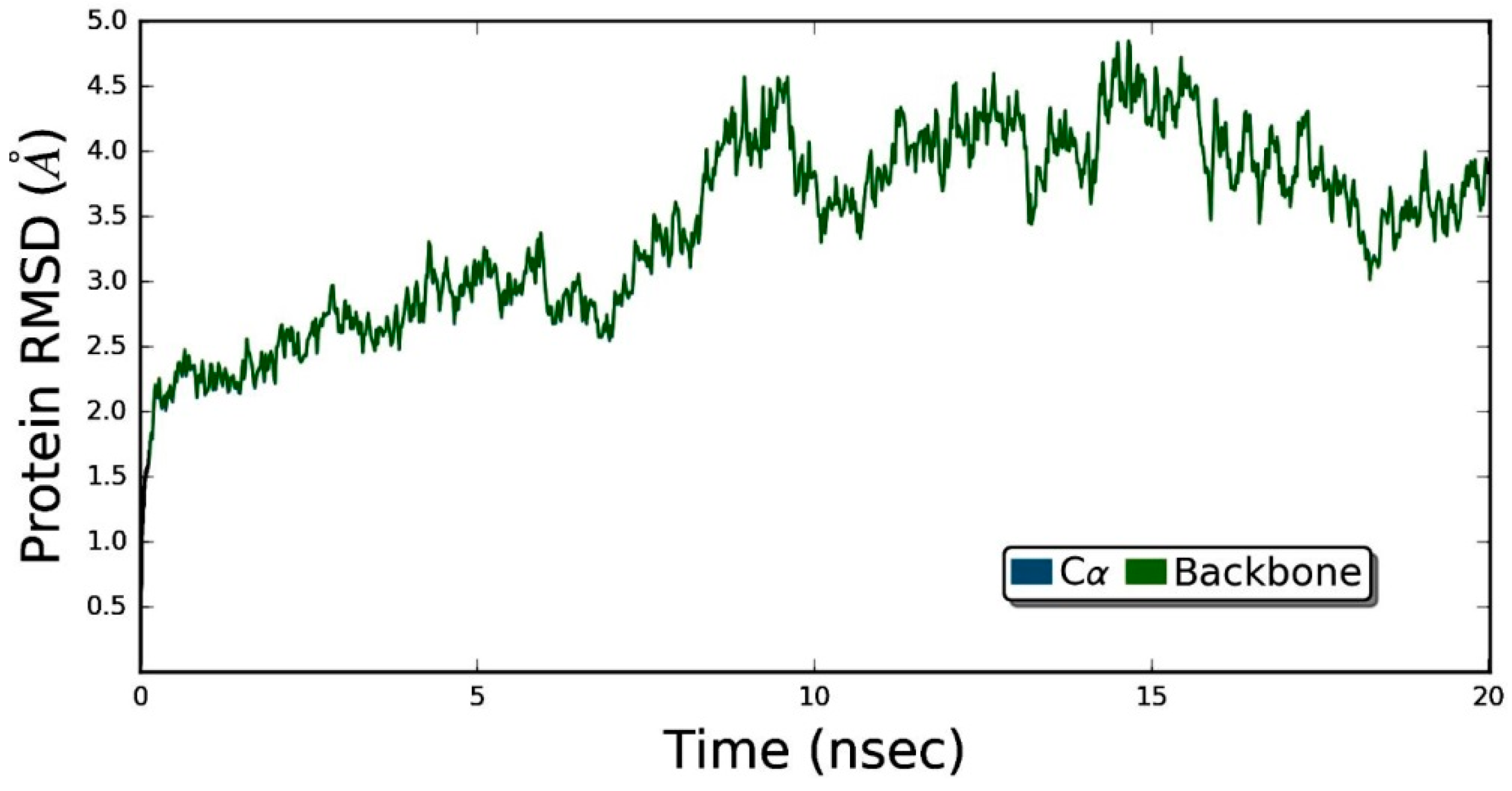
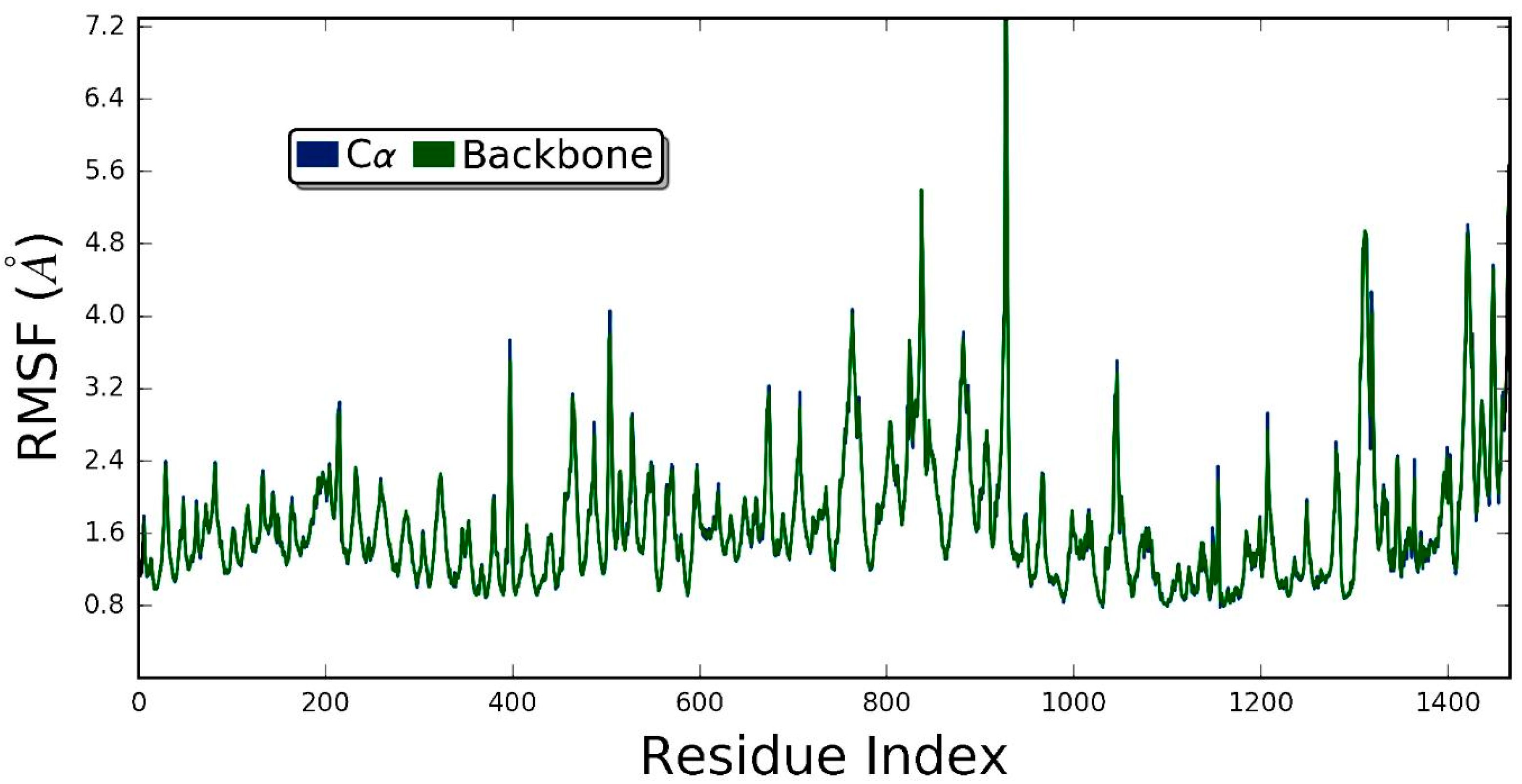
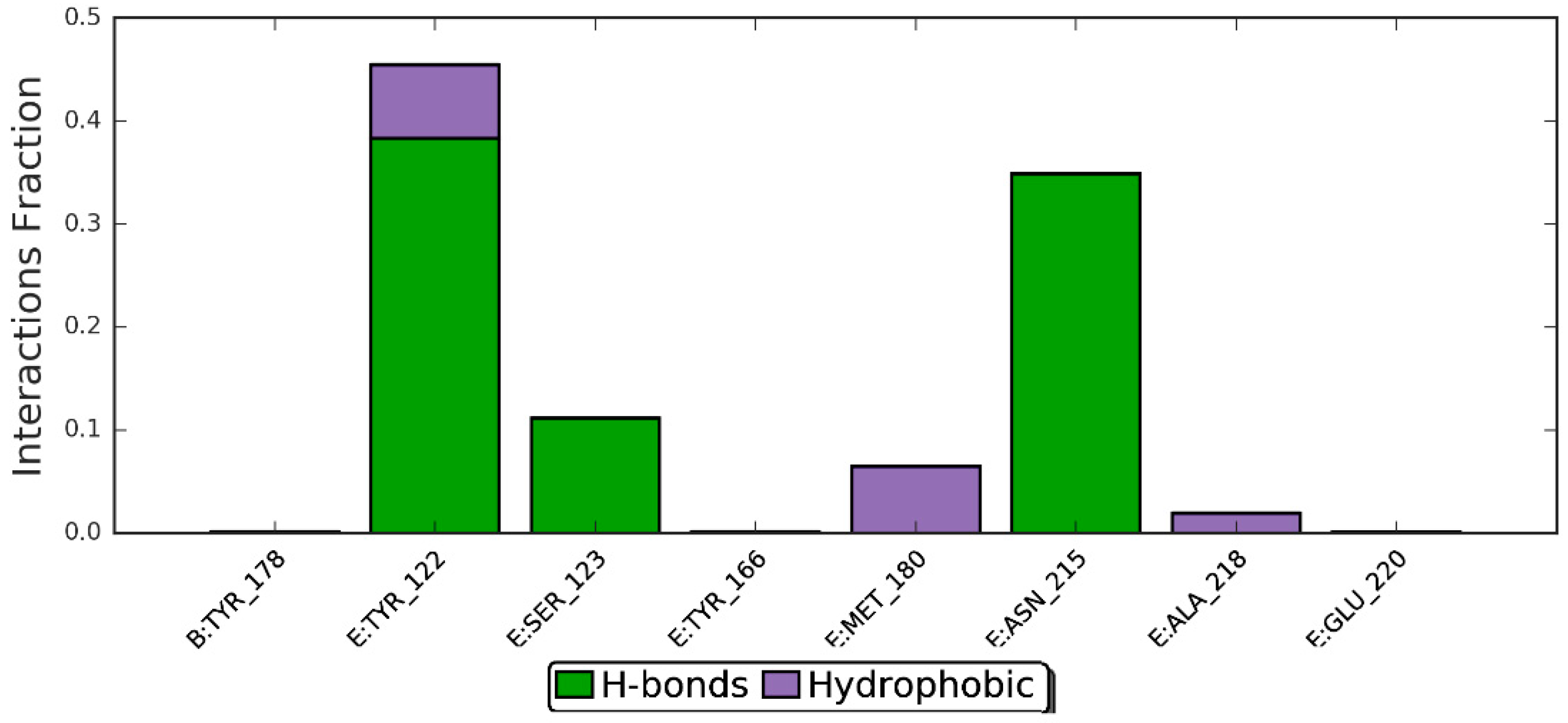
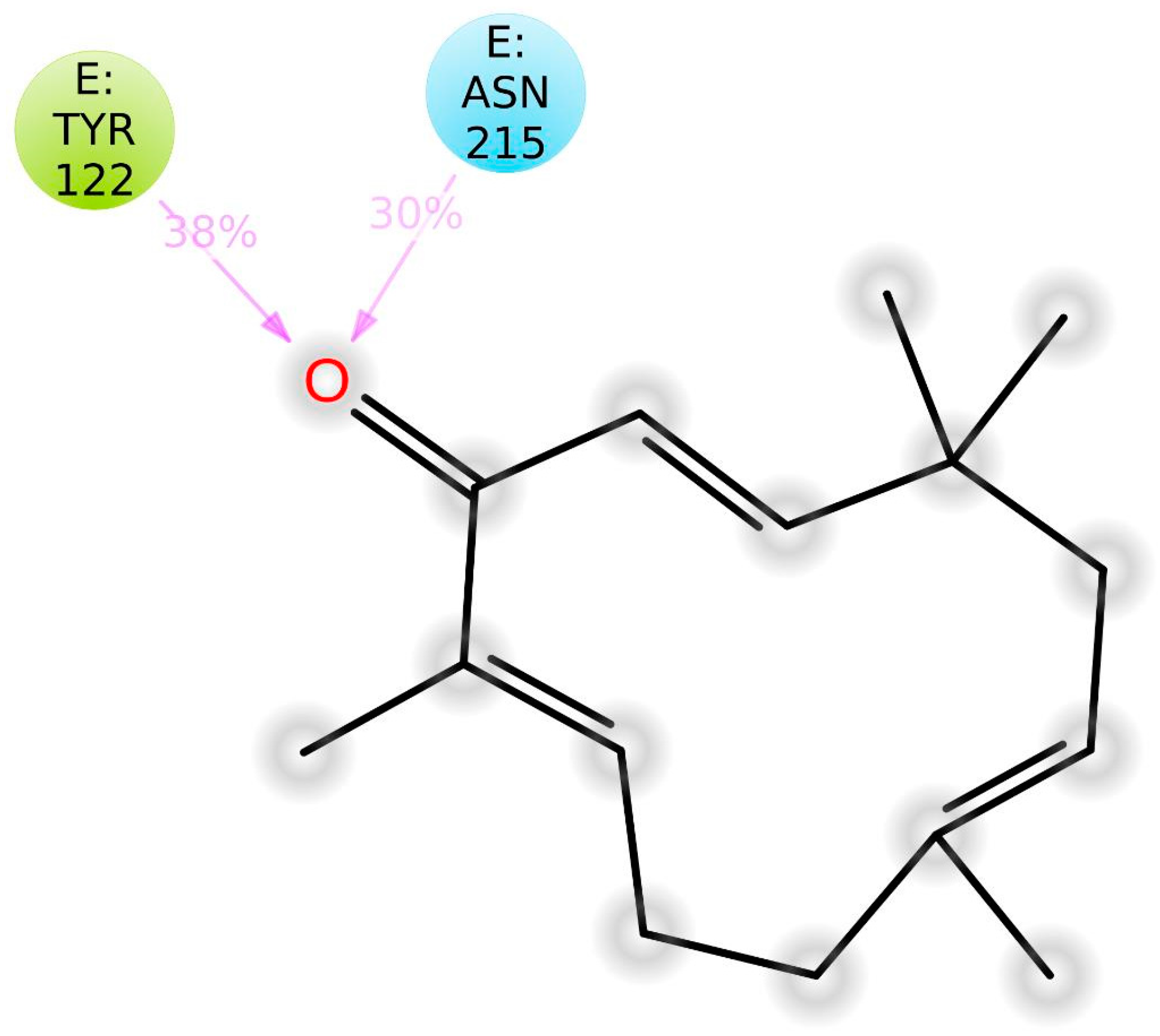
3.5. Molecular Dynamics Simulation Studies of ZER in Complex with TP5-iRGD/Integrin αvβ3
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca: A Cancer J. Clin. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Yang, J.; Zhang, Q.; Yang, J.; Wang, H.; Xu, J.; Zheng, J. iRGD as a tumorpenetrating peptide for cancer therapy (Review). Mol. Med. Rep. 2017, 15, 2925–2930. [Google Scholar] [CrossRef] [PubMed]
- Minchinton, A.I.; Tannock, I.F. Drug penetration in solid tumours. Nat. Rev. Cancer 2006, 6, 583. [Google Scholar] [CrossRef] [PubMed]
- Hambley, T.W.; Hait, W.N. Is anticancer drug development heading in the right direction? Cancer Res. 2009, 69, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
- Hai-Tao, Z.; Hui-Cheng, L.; Zheng-Wu, L.; Chang-Hong, G. A tumor-penetrating peptide modification enhances the antitumor activity of endostatin in vivo. Anti-Cancer Drugs 2011, 22, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Rabbani-Chadegani, A.; Paydar, P.; Amirshenava, M.; Aramvash, A. An in vitro study on the effect of vinca alkaloid, vinorelbine, on chromatin histone, HMGB proteins and induction of apoptosis in mice non-adherent bone marrow cells. Drug Chem. Toxicol. 2015, 38, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Böhme, D.; Beck-Sickinger, A.G. Drug delivery and release systems for targeted tumor therapy. J. Pept. Sci. 2015, 21, 186–200. [Google Scholar]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Guo, W.; Giancotti, F.G. Integrin signalling during tumour progression. Nat. Rev. Mol. Cell Biol. 2004, 5, 816. [Google Scholar] [CrossRef]
- Xiong, J.-P.; Stehle, T.; Zhang, R.; Joachimiak, A.; Frech, M.; Goodman, S.L.; Arnaout, M.A. Crystal structure of the extracellular segment of integrin αVβ3 in complex with an Arg-Gly-Asp ligand. Science 2002, 296, 151–155. [Google Scholar] [CrossRef]
- Dutta, S.; Mahalanobish, S.; Saha, S.; Ghosh, S.; Sil, P.C. Natural products: An upcoming therapeutic approach to cancer. Food Chem. Toxicol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Khazir, J.; Mir, B A.; Mir, S.A.; Cowan, D. Natural products as lead compounds in drug discovery. Taylor Fr. 2013. [Google Scholar]
- Eid, E.E.; Abdul, A.B.; Suliman, F.E.O.; Sukari, M.A.; Rasedee, A.; Fatah, S.S. Characterization of the inclusion complex of zerumbone with hydroxypropyl-β-cyclodextrin. Carbohydr. Polym. 2011, 83, 1707–1714. [Google Scholar] [CrossRef]
- Sung, B.; Murakami, A.; Oyajobi, B.O.; Aggarwal, B.B. Zerumbone abolishes RANKL-induced NF-κB activation, inhibits osteoclastogenesis, and suppresses human breast cancer–induced bone loss in athymic nude mice. Cancer Res. 2009, 69, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, A.; Arlotti, J.A.; Murakami, A.; Singh, S.V. Zerumbone causes Bax-and Bak-mediated apoptosis in human breast cancer cells and inhibits orthotopic xenograft growth in vivo. Breast Cancer Res. Treat. 2012, 136, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Bae, S.Y.; Oh, S.J.; Lee, J.; Lee, J.H.; Lee, H.C.; Lee, S.K.; Kil, W.H.; Kim, S.W.; Nam, S.J. Zerumbone Suppresses IL-1β-induced Cell Migration and Invasion by Inhibiting IL-8 and MMP-3 Expression in Human Triple-negative Breast Cancer Cells. Phytother. Res. 2014, 28, 1654–1660. [Google Scholar] [CrossRef]
- Kim, S.; Kil, W.H.; Lee, J.; Oh, S.-J.; Han, J.; Jeon, M.; Jung, T.; Lee, S.K.; Bae, S.Y.; Lee, H.C. Zerumbone suppresses EGF-induced CD44 expression through the inhibition of STAT3 in breast cancer cells. Oncol. Rep. 2014, 32, 2666–2672. [Google Scholar] [CrossRef] [Green Version]
- Eid, E.E.; Bustamam Abdul, A.; Rasedee, A.; Suliman, F.E.O.; Sukari, M.A.; Fatah, S.A. Liquid chromatography–tandem mass spectroscopic method for the determination of zerumbone in human plasma and its application to pharmacokinetics. J. Mass Spectrom. 2011, 46, 772–781. [Google Scholar] [CrossRef]
- Snyder, E.L.; Dowdy, S.F. Cell penetrating peptides in drug delivery. Pharm. Res. 2004, 21, 389–393. [Google Scholar] [CrossRef]
- Karagiannis, E.D.; Urbanska, A.M.; Sahay, G.; Pelet, J.M.; Jhunjhunwala, S.; Langer, R.; Anderson, D.G. Rational design of a biomimetic cell penetrating peptide library. Acs Nano 2013, 7, 8616–8626. [Google Scholar] [CrossRef]
- Yamashita, H.; Kato, T.; Oba, M.; Misawa, T.; Hattori, T.; Ohoka, N.; Tanaka, M.; Naito, M.; Kurihara, M.; Demizu, Y. Development of a Cell-penetrating Peptide that Exhibits Responsive Changes in its Secondary Structure in the Cellular Environment. Sci. Rep. 2016, 6, 33003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundin, P.; Johansson, H.; Guterstam, P.; Holm, T.; Hansen, M.; Langel, Ü.; El Andaloussi, S. Distinct uptake routes of cell-penetrating peptide conjugates. Bioconjugate Chem. 2008, 19, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Tünnemann, G.; Martin, R.M.; Haupt, S.; Patsch, C.; Edenhofer, F.; Cardoso, M.C. Cargo-dependent mode of uptake and bioavailability of TAT-containing proteins and peptides in living cells. Faseb J. 2006, 20, 1775–1784. [Google Scholar]
- Raucher, D.; Ryu, J.S. Cell-penetrating peptides: Strategies for anticancer treatment. Trends Mol. Med. 2015, 21, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Regberg, J.; Srimanee, A.; Langel, Ü. Applications of cell-penetrating peptides for tumor targeting and future cancer therapies. Pharmaceuticals 2012, 5, 991–1007. [Google Scholar] [CrossRef] [PubMed]
- Heitz, F.; Morris, M.C.; Divita, G. Twenty years of cell-penetrating peptides: From molecular mechanisms to therapeutics. Br. J. Pharmacol. 2009, 157, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Sarko, D.; Beijer, B.; Boy, R.G.; Nothelfer, E.-M.; Leotta, K.; Eisenhut, M.; Altmann, A.; Haberkorn, U.; Mier, W. The pharmacokinetics of cell-penetrating peptides. Mol. Pharm. 2010, 7, 2224–2231. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, D.M.; Langel, K.; Eriste, E.; Langel, U. Cell-penetrating peptides: Design, synthesis, and applications. Acs Nano 2014, 8, 1972–1994. [Google Scholar] [CrossRef]
- Bolhassani, A. Potential efficacy of cell-penetrating peptides for nucleic acid and drug delivery in cancer. Biochim. Et Biophys. Acta (Bba)-Rev. Cancer 2011, 1816, 232–246. [Google Scholar] [CrossRef]
- Lehto, T.; Kurrikoff, K.; Langel, Ü. Cell-penetrating peptides for the delivery of nucleic acids. Expert Opin. Drug Deliv. 2012, 9, 823–836. [Google Scholar] [CrossRef]
- Sugahara, K.N.; Teesalu, T.; Karmali, P.P.; Kotamraju, V.R.; Agemy, L.; Greenwald, D.R.; Ruoslahti, E. Coadministration of a tumor-penetrating peptide enhances the efficacy of cancer drugs. Science 2010, 328, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Bechara, C.; Sagan, S. Cell-penetrating peptides: 20 years later, where do we stand? Febs Lett. 2013, 587, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, Q.; Yu, Z.; He, Q. Cell-penetrating peptide-based intelligent liposomal systems for enhanced drug delivery. Curr. Pharm. Biotechnol. 2014, 15, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Kandela, I.; Chou, J.; Chow, K. Registered report: Coadministration of a tumor-penetrating peptide enhances the efficacy of cancer drugs. eLife 2015, 4, e06959. [Google Scholar] [CrossRef] [PubMed]
- Lao, X.; Li, B.; Liu, M.; Chen, J.; Gao, X.; Zheng, H. Increased antitumor activity of tumor-specific peptide modified thymopentin. Biochimie 2014, 107, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.-Q.; Gao, W.; Xiang, B.; Qi, X.-R. Enhancing cellular uptake of activable cell-penetrating peptide-doxorubicin conjugate by enzymatic cleavage. Int. J. Nanomed. 2012, 7, 1613–1621. [Google Scholar]
- Aroui, S.; Brahim, S.; De Waard, M.; Kenani, A. Cytotoxicity, intracellular distribution and uptake of doxorubicin and doxorubicin coupled to cell-penetrating peptides in different cell lines: A comparative study. Biochem. Biophys. Res. Commun. 2010, 391, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Cun, X.; Chen, J.; Ruan, S.; Zhang, L.; Wan, J.; He, Q.; Gao, H. A novel strategy through combining iRGD peptide with tumor-microenvironment-responsive and multistage nanoparticles for deep tumor penetration. Acs Appl. Mater. Interfaces 2015, 7, 27458–27466. [Google Scholar] [CrossRef]
- Hu, C.; Yang, X.; Liu, R.; Ruan, S.; Zhou, Y.; Xiao, W.; Yu, W.; Yang, C.; Gao, H. Coadministration of iRGD with multistage responsive nanoparticles enhanced tumor targeting and penetration abilities for breast cancer therapy. Acs Appl. Mater. Interfaces 2018, 10, 22571–22579. [Google Scholar] [CrossRef]
- Tang, E.L.; Rajarajeswaran, J.; Fung, S.Y.; Kanthimathi, M. Antioxidant activity of Coriandrum sativum and protection against DNA damage and cancer cell migration. BMC Complement. Altern. Med. 2013, 13, 347. [Google Scholar] [CrossRef]
- Ciapetti, G.; Granchi, D.; Savarino, L.; Cenni, E.; Magrini, E.; Baldini, N.; Giunti, A. In vitro testing of the potential for orthopedic bone cements to cause apoptosis of osteoblast-like cells. Biomaterials 2002, 23, 617–627. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Huey, R.; Olson, A.J. Distributed automated docking of flexible ligands to proteins: Parallel applications of AutoDock 2.4. J. Comput.-Aided Mol. Des. 1996, 10, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D. Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006. [Google Scholar]
- Desmond Molecular Dynamics System. Maestro-Desmond Interoperability Tools; Schrödinger: New York, NY, USA, 2016. [Google Scholar]
- Harder, E.; Damm, W.; Maple, J.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L. OPLS3: A force field providing broad coverage of drug-like small molecules and proteins. J. Chem. Theory Comput. 2015, 12, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1695. [Google Scholar] [CrossRef]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Tuckerman, M.; Berne, B.J.; Martyna, G.J. Reversible multiple time scale molecular dynamics. J. Chem. Phys. 1992, 97, 1990–2001. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Ji, Y. RGD-modified polymer and liposome nanovehicles: Recent research progress for drug delivery in cancer therapeutics. Eur. J. Pharm. Sci. 2018, 128, 8–17. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Mao, X.; Liu, J.; Gong, Z.; Zhang, H.; Lu, Y.; Zou, H.; Yu, Y.; Chen, Y.; Sun, Z.; Li, W. iRGD-conjugated DSPE-PEG2000 nanomicelles for targeted delivery of salinomycin for treatment of both liver cancer cells and cancer stem cells. Nanomedicine 2015, 10, 2677–2695. [Google Scholar] [CrossRef]
- Eid, E.E.; Azam, F.; Hassan, M.; Taban, I.M.; Halim, M.A. Zerumbone binding to estrogen receptors: An in-silico investigation. J. Recept. Signal. Transduct. 2018, 38, 342–351. [Google Scholar] [CrossRef]
- Azam, F.; Alabdullah, N.H.; Ehmedat, H.M.; Abulifa, A.R.; Taban, I.; Upadhyayula, S. NSAIDs as potential treatment option for preventing amyloid β toxicity in Alzheimer’s disease: An investigation by docking, molecular dynamics, and DFT studies. J. Biomol. Struct. Dyn. 2018, 36, 2099–2117. [Google Scholar] [CrossRef] [PubMed]
- Maiorov, V.N.; Crippen, G.M. Significance of root-mean-square deviation in comparing three-dimensional structures of globular proteins. J. Mol. Biol. 1994, 235, 625–634. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | ZER | ZER-HPβCD | TP5-iRGD Peptide | |||||
|---|---|---|---|---|---|---|---|---|
| 24 h IC50 (µg/mL) | 48 h IC50 (µg/mL) | 72 h IC50 (µg/mL) | 24 h IC50 (µg/mL) | 48 h IC50 (µg/mL) | 72 h IC50 (µg/mL) | 48 h IC50 (µg/mL) | 72 h IC50 (µg/mL) | |
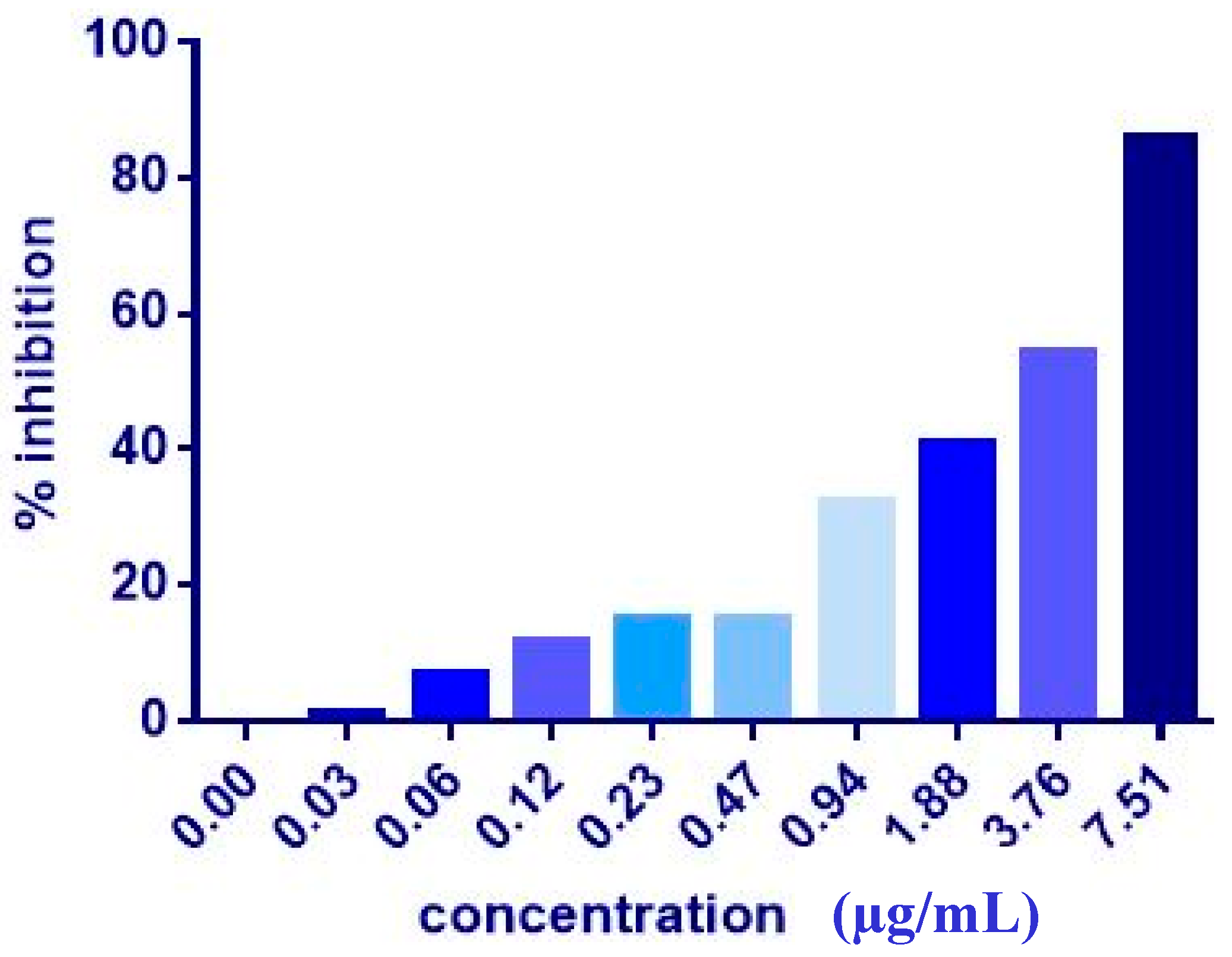
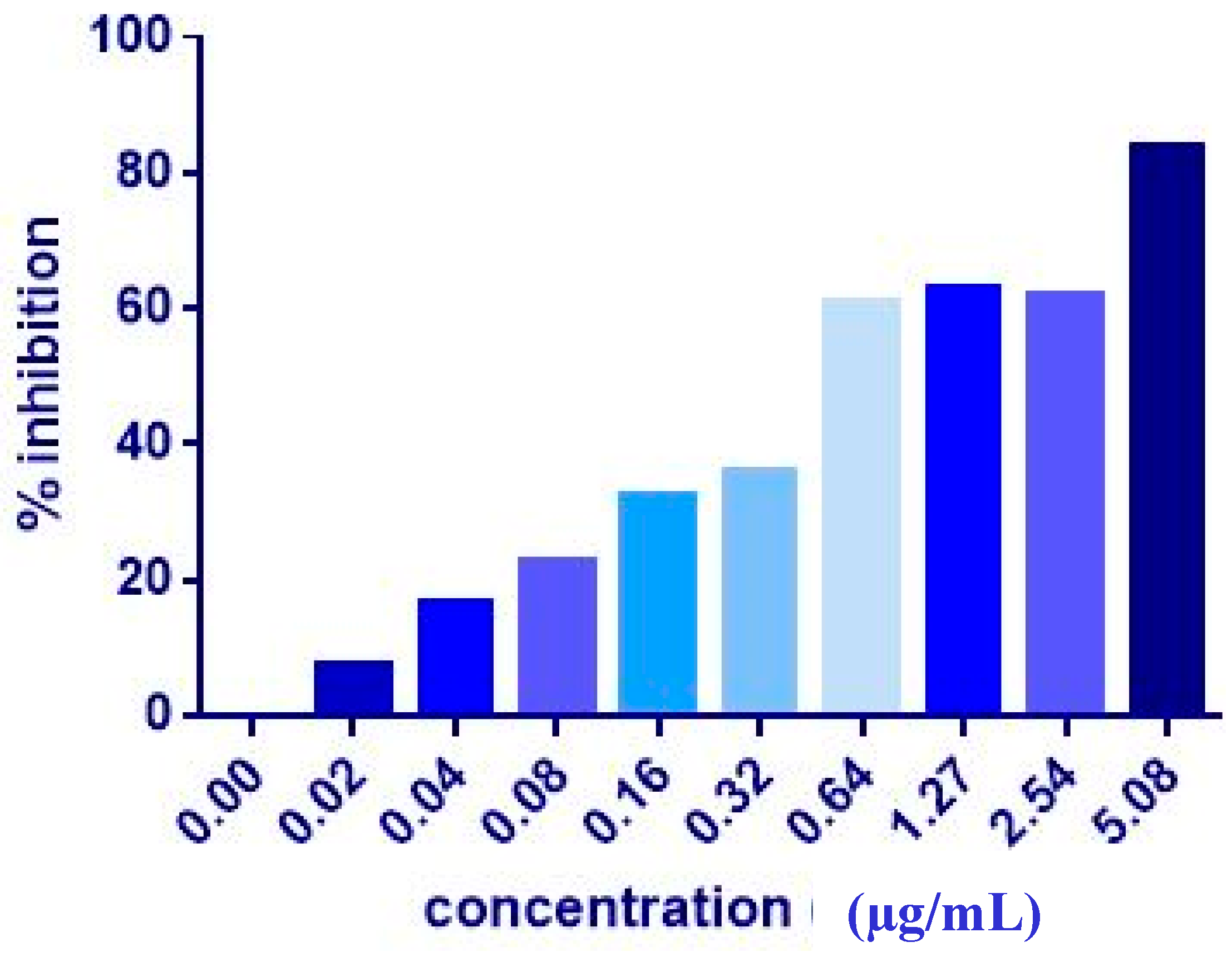
| MCF7 | 17.36 ± 2.3 | 9.06 ± 1.0 | 7.51 ± 0.2 | 20.81 ± 9.3 | 4.43 ± 0.2 | 5.08 ± 0.2 | >1000 | >1000 |
| MDA-MB-231 | 50.93 ± 3.0 | 23.44 ± 10.0 | 14.96 ± 1.5 | 40.73 ± 1.9 | 21.1 ± 6.7 | 12.18 ± 0.7 | >1000 | >1000 |
| Hs27 | >100 | >100 | >100 | 42.99 ± 2.3 | 46.14 ± 2.4 | 21.45 ± 1.3 | >1000 | >1000 |
| Co-administration Ratio | MCF-7 (% of Inhibition) | MDA-MB-231 (% of Inhibition) | ||
|---|---|---|---|---|
| Compound (IC50): TP5-iRGD | ZER | ZER-HPβCD | ZER | ZER-HPβCD |
| 1:10 X | 91.4 ± 3.0 | 90.4 ± 4.2 | 58.1 ± 8.8 | 41.5 ± 20.4 |
| 1:20 X | 67.6 ± 3.9 | 85.8 ± 7.0 | 44.0 ± 8.6 | 72.8 ± 2.3 |
| 1:30 X | 72.1 ± 8.6 | 68.5 ± 13.2 | 48.3 ± 5.4 | 79.1 ± 1.8 |
| Co-Administration | MCF-7–IC50 (µg/mL) |
|---|---|
| ZER with TP5-iRGD | 3.13 ± 0.7 |
| ZER-HPβCDwith TP5-iRGD | 0.49 ± 0.004 |
| Parameters | ZER | ZER in the Presence of TP5-iRGD |
|---|---|---|
| Binding Energy, ΔGb (kcal/mol) | −6.77 | −8.13 |
| Predicted Ki (µM) | 10.97 | 1.10 |
| RMSD | 4.39 | 4.16 |
| Residues involved in hydrophobic interactions | Met-180(B), Arg-214(B) | Tyr-122(B), Tyr-166(B), Tyr-178(A), Arg-214(B), Arg-216(B), Ala-218(B) |
| Residues involved in hydrophilic interactions | Nil | Nil |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
E. M. Eid, E.; S. Alanazi, A.; Koosha, S.; A. Alrasheedy, A.; Azam, F.; M. Taban, I.; Khalilullah, H.; Sadiq Al-Qubaisi, M.; A. Alshawsh, M. Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide. Molecules 2019, 24, 2554. https://doi.org/10.3390/molecules24142554
E. M. Eid E, S. Alanazi A, Koosha S, A. Alrasheedy A, Azam F, M. Taban I, Khalilullah H, Sadiq Al-Qubaisi M, A. Alshawsh M. Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide. Molecules. 2019; 24(14):2554. https://doi.org/10.3390/molecules24142554
Chicago/Turabian StyleE. M. Eid, Eltayeb, Abdulrahman S. Alanazi, Sanaz Koosha, Alian A. Alrasheedy, Faizul Azam, Ismail M. Taban, Habibullah Khalilullah, Mothanna Sadiq Al-Qubaisi, and Mohammed A. Alshawsh. 2019. "Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide" Molecules 24, no. 14: 2554. https://doi.org/10.3390/molecules24142554
APA StyleE. M. Eid, E., S. Alanazi, A., Koosha, S., A. Alrasheedy, A., Azam, F., M. Taban, I., Khalilullah, H., Sadiq Al-Qubaisi, M., & A. Alshawsh, M. (2019). Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide. Molecules, 24(14), 2554. https://doi.org/10.3390/molecules24142554