Complement Activation-Related Pathophysiological Changes in Anesthetized Rats: Activator-Dependent Variations of Symptoms and Mediators of Pseudoallergy


 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
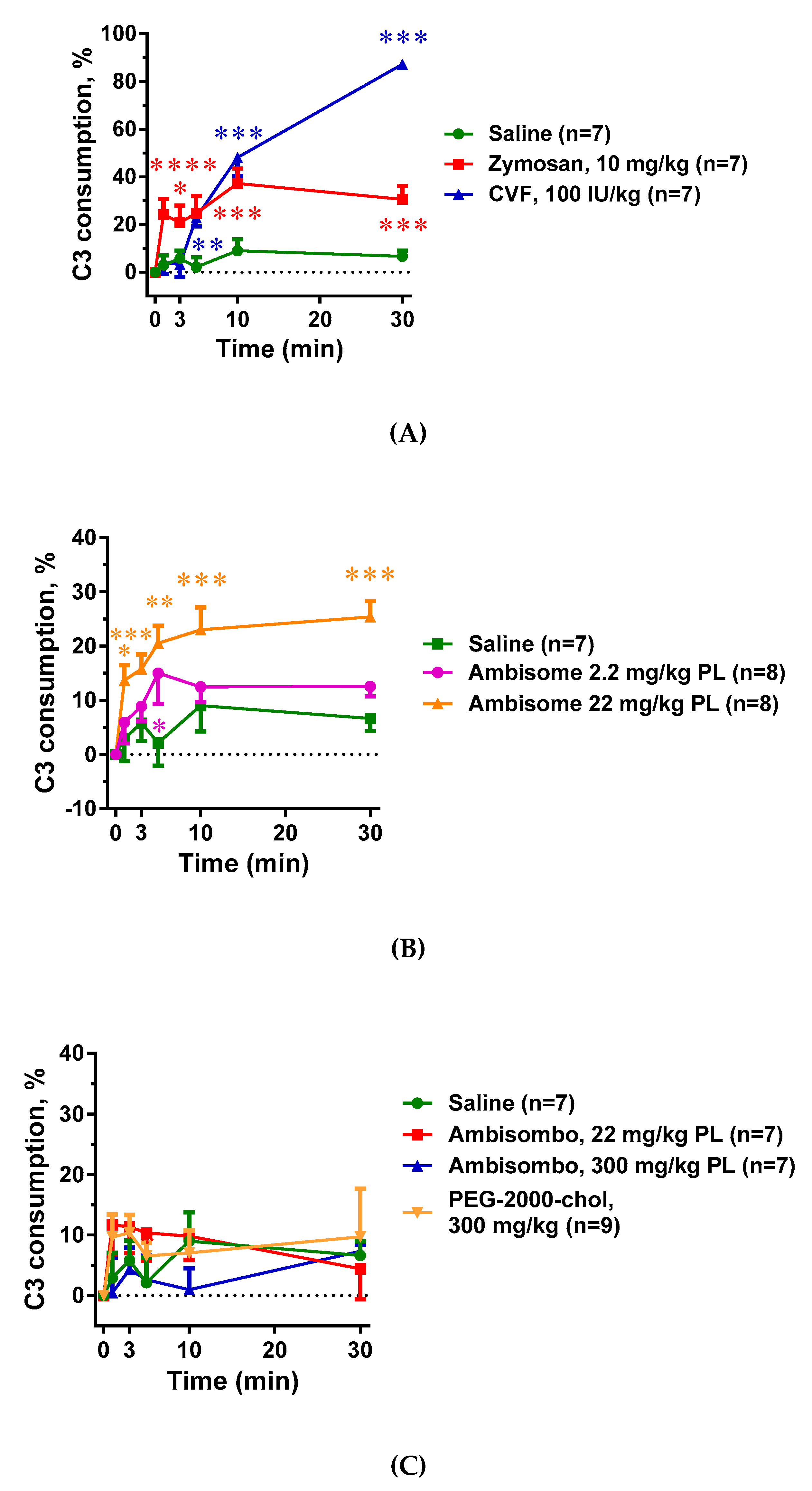
2.1. Complement Activation by the Different Reaction Inducers
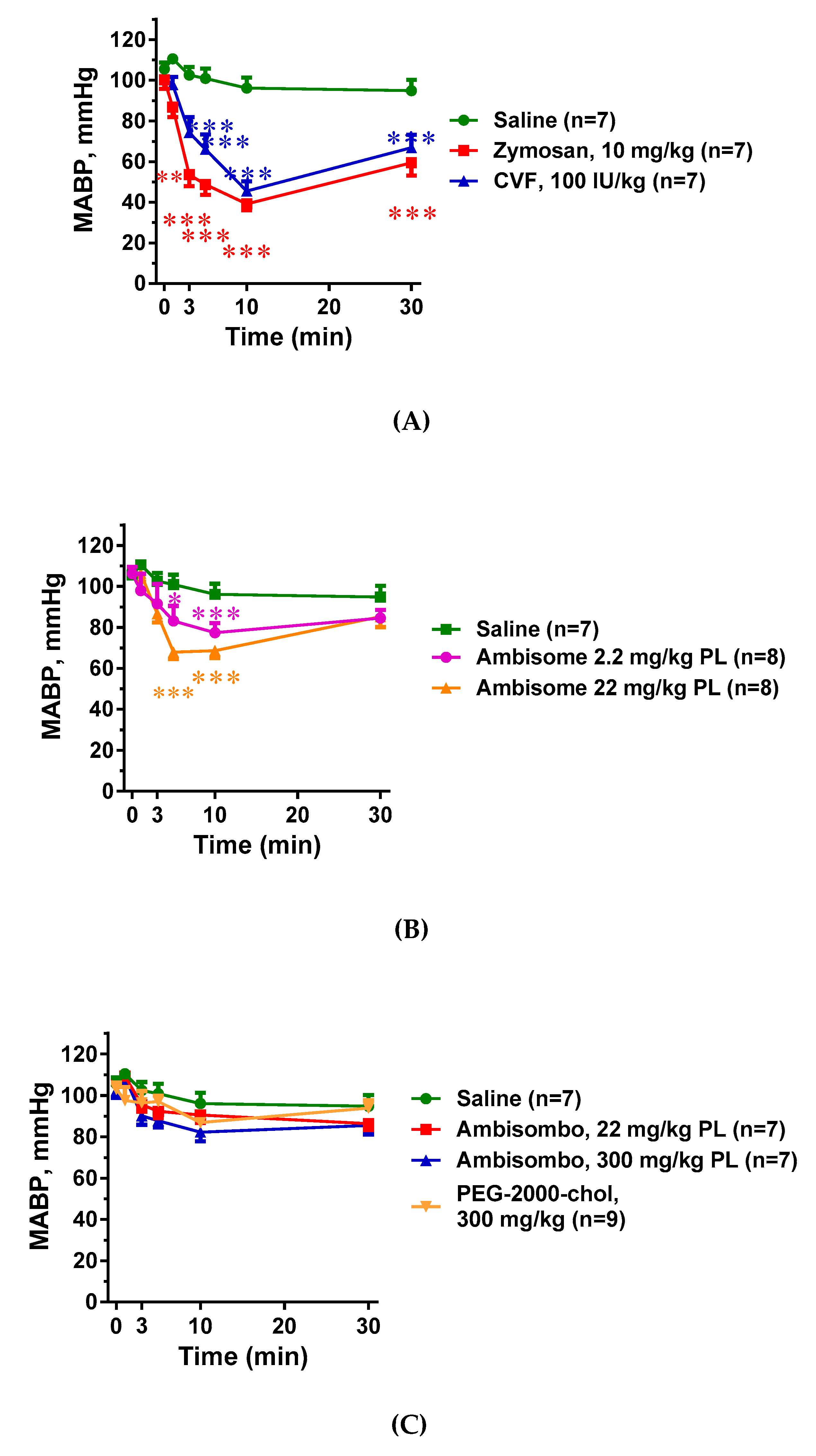
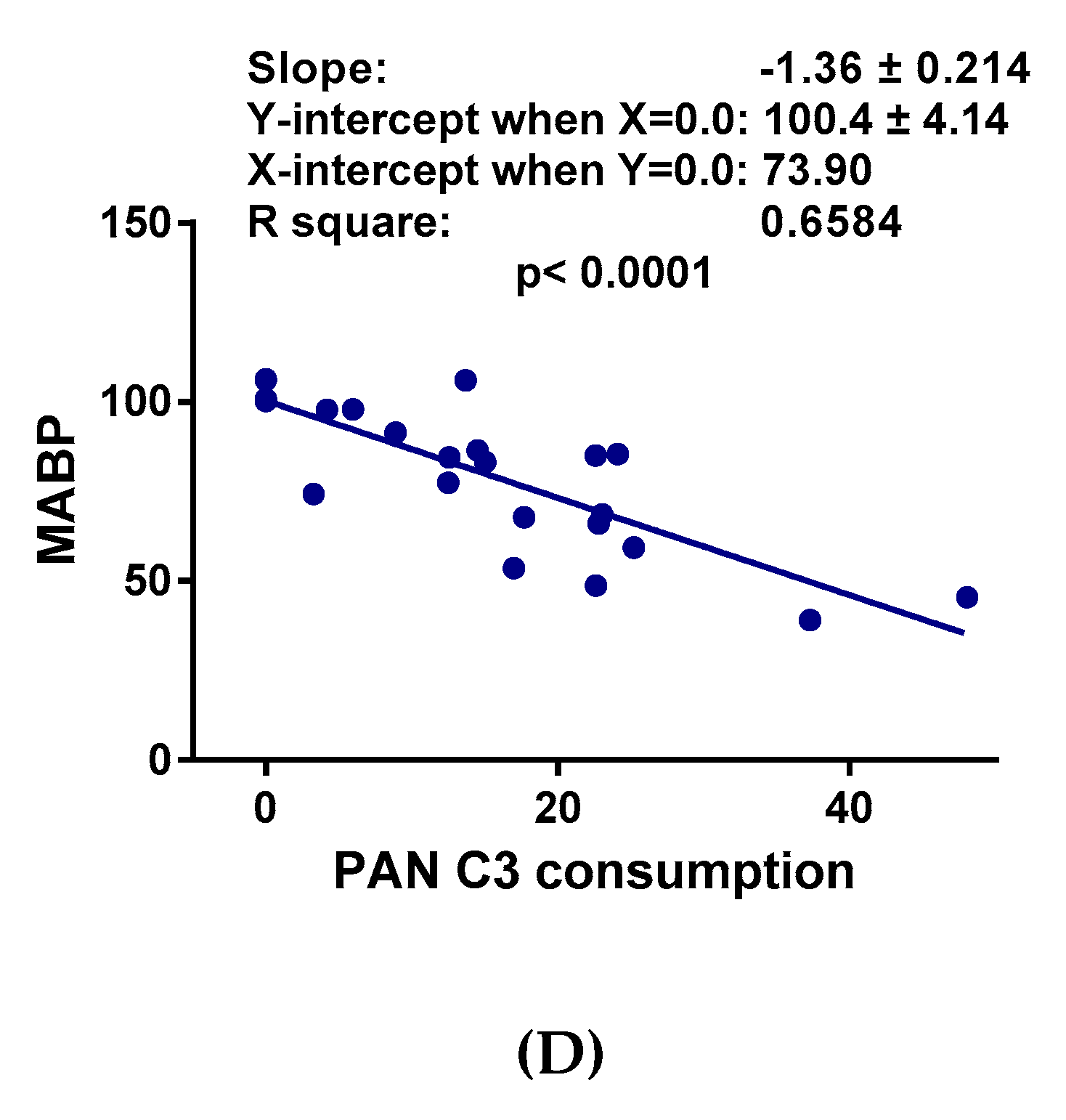
2.2. Hemodynamic Changes and Their Correlation with C Activation
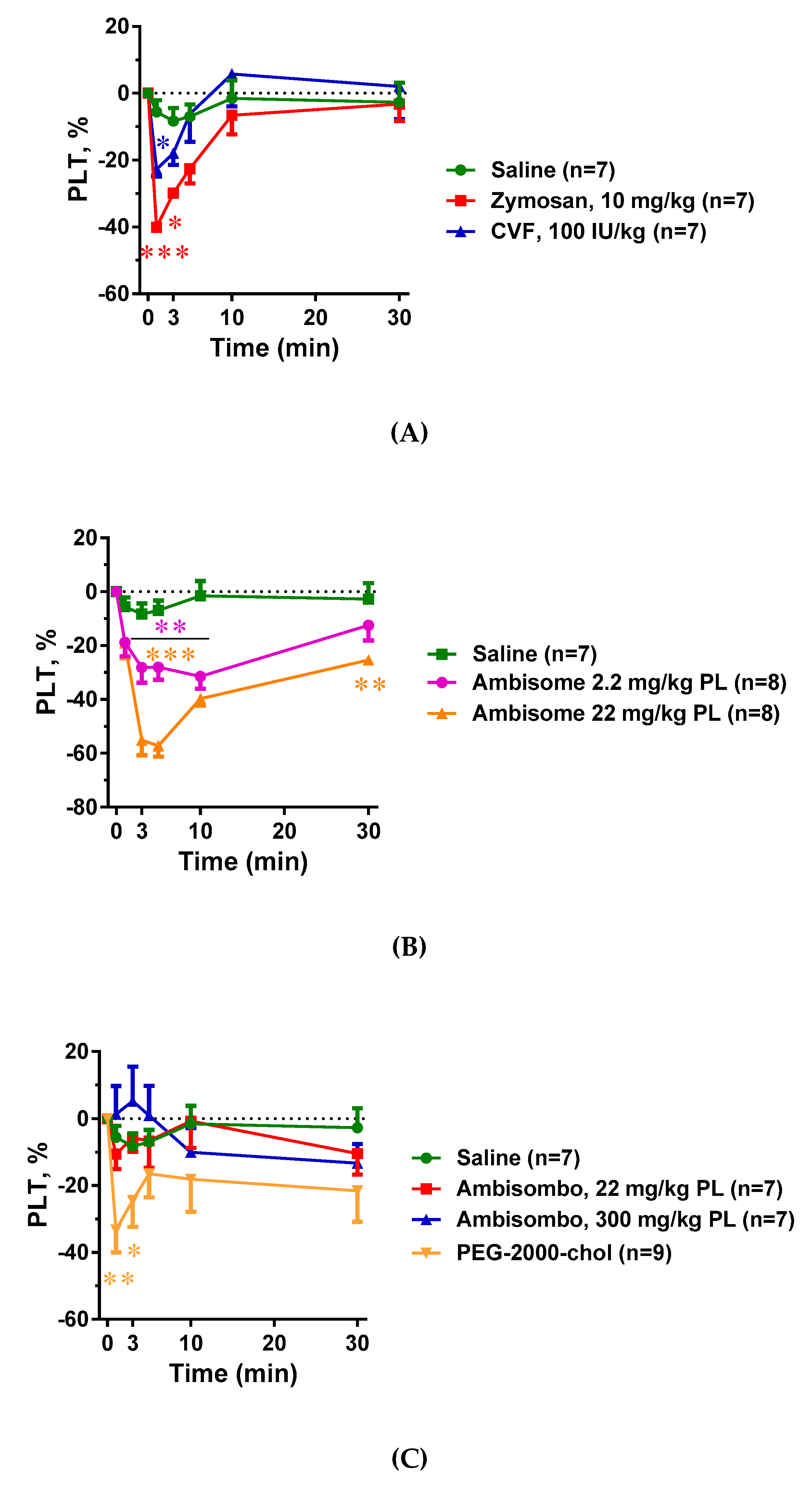
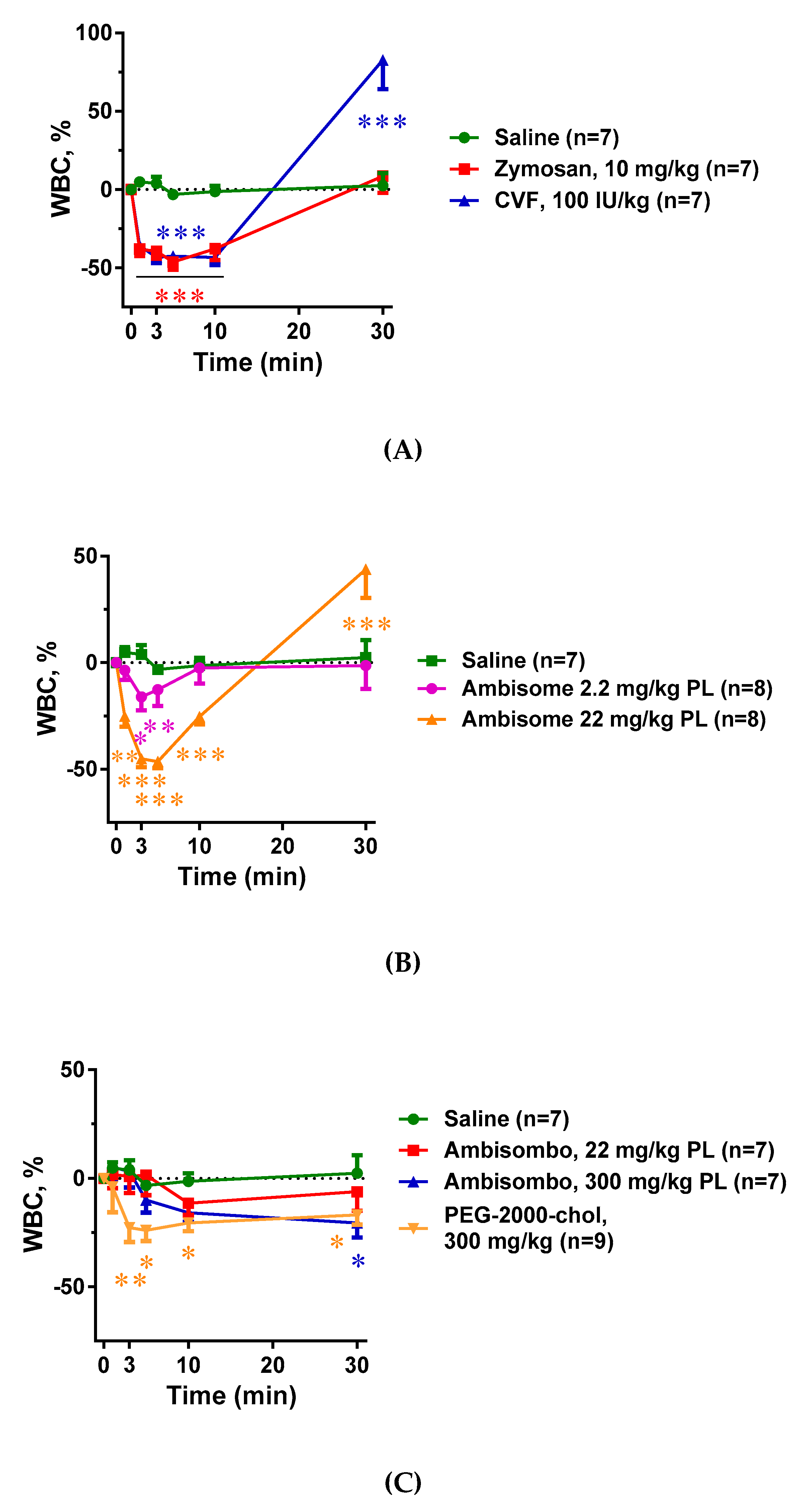
2.3. Blood Cell Changes and Their Correlation with C Activation
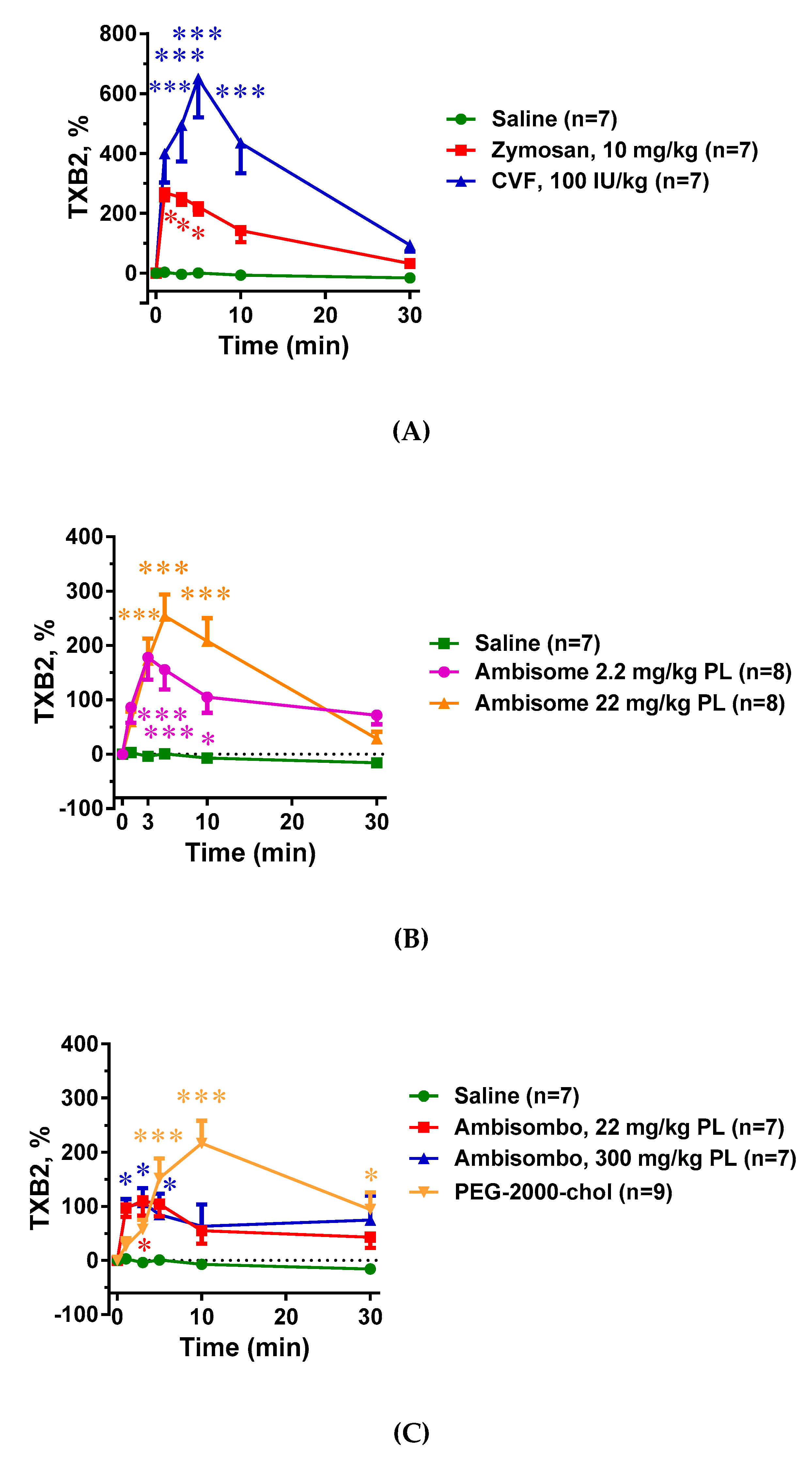
2.4. Plasma TXB2 Changes and Their Correlation with C Activation and Systemic Hypotension
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Ethical Approval
4.3. Liposomes and Chemicals
4.4. Experimental Protocol
4.5. Analytical Procedures
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Otte, A.; Soh, B.K.; Yoon, G.; Park, K. Liquid crystalline drug delivery vehicles for oral and IV/subcutaneous administration of poorly soluble (and soluble) drugs. Int. J. Pharm. 2018, 539, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Watermann, A.; Brieger, J. Mesoporous Silica Nanoparticles as Drug Delivery Vehicles in Cancer. Nanomaterials 2017, 7, 189. [Google Scholar] [CrossRef]
- Szebeni, J.; Simberg, D.; Gonzalez-Fernandez, A.; Barenholz, Y.; Dobrovolskaia, M.A. Roadmap and strategy for overcoming infusion reactions to nanomedicines. Nat. Nanotechnol. 2018, 13, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J. Mechanism of nanoparticle-induced hypersensitivity in pigs: Complement or not complement? Drug Discov. Today 2018, 23, 487–492. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Tang, M. Safety of novel liposomal drugs for cancer treatment: Advances and prospects. Chem. Biol. Interact. 2018, 295, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Loo, A.S.; Muhsin, S.A.; Walsh, T.J. Toxicokinetic and mechanistic basis for the safety and tolerability of liposomal amphotericin B. Expert Opin. Drug Saf. 2013, 12, 881–895. [Google Scholar] [CrossRef]
- Szebeni, J. Complement activation-related pseudoallergy: A stress reaction in blood triggered by nanomedicines and biologicals. Mol. Immunol. 2014, 61, 163–173. [Google Scholar] [CrossRef]
- Wibroe, P.P.; Anselmo, A.C.; Nilsson, P.H.; Sarode, A.; Gupta, V.; Urbanics, R.; Szebeni, J.; Hunter, A.C.; Mitragotri, S.; Mollnes, T.E.; et al. Bypassing adverse injection reactions to nanoparticles through shape modification and attachment to erythrocytes. Nat. Nanotechnol. 2017, 12, 589–594. [Google Scholar] [CrossRef]
- Orfi, E.; Meszaros, T.; Hennies, M.; Fulop, T.; Dezsi, L.; Nardocci, A.; Rosivall, L.; Hamar, P.; Neun, B.W.; Dobrovolskaia, M.A.; et al. Acute physiological changes caused by complement activators and amphotericin B-containing liposomes in mice. Int. J. Nanomed. 2019, 14, 1563–1573. [Google Scholar] [CrossRef]
- Zamboni, W.C.; Szebeni, J.; Kozlov, S.V.; Lucas, A.T.; Piscitelli, J.A.; Dobrovolskaia, M.A. Animal models for analysis of immunological responses to nanomaterials: Challenges and considerations. Adv. Drug Deliv. Rev. 2018, 136, 82–96. [Google Scholar] [CrossRef]
- Rabinovici, R.; Rudolph, A.S.; Feuerstein, G. Characterization of hemodynamic, hematologic, and biochemical responses to administration of liposome-encapsulated hemoglobin in the conscious, freely moving rat. Circ. Shock 1989, 29, 115–132. [Google Scholar] [PubMed]
- Rabinovici, R.; Rudolph, A.S.; Yue, T.L.; Feuerstein, G. Biological responses to liposome-encapsulated hemoglobin (LEH) are improved by a PAF antagonist. Circ. Shock 1990, 31, 431–445. [Google Scholar]
- Goins, B.; Phillips, W.T.; Klipper, R.; Rudolph, A.S. Role of complement in rats injected with liposome-encapsulated hemoglobin. J. Surg. Res. 1997, 68, 99–105. [Google Scholar] [CrossRef]
- Szebeni, J.; Wassef, N.M.; Spielberg, H.; Rudolph, A.S.; Alving, C.R. Complement activation in rats by liposomes and liposome-encapsulated hemoglobin: Evidence for anti-lipid antibodies and alternative pathway activation. Biochem. Biophys. Res. Commun. 1994, 205, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Spielberg, H.; Cliff, R.O.; Wassef, N.M.; Rudolph, A.S.; Alving, C.R. Complement activation and thromboxane secretion by liposome-encapsulated hemoglobin in rats in vivo: Inhibition by soluble complement receptor type 1. Artif. Cells Blood Substit. Immobil. Biotechnol. 1997, 25, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Baranyi, L.; Szebeni, J.; Sávay, S.; Bodo, M.; Basta, M.; Bentley, T.B.; Bunger, R.; Alving, C.R. Complement-Dependent Shock and Tissue Damage Induced by Intravenous Injection of Cholesterol-Enriched Liposomes in Rats. J. Appl. Res. 2003, 3, 221–231. [Google Scholar]
- Dezsi, L.; Fulop, T.; Meszaros, T.; Szenasi, G.; Urbanics, R.; Vazsonyi, C.; Orfi, E.; Rosivall, L.; Nemes, R.; Kok, R.J.; et al. Features of complement activation-related pseudoallergy to liposomes with different surface charge and PEGylation: Comparison of the porcine and rat responses. J. Control. Release 2014, 195, 2–10. [Google Scholar] [CrossRef] [PubMed]
- van den Hoven, J.M.; Nemes, R.; Metselaar, J.M.; Nuijen, B.; Beijnen, J.H.; Storm, G.; Szebeni, J. Complement activation by PEGylated liposomes containing prednisolone. Eur. J. Pharm. Sci. 2013, 49, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Proctor, L.M.; Moore, T.A.; Monk, P.N.; Sanderson, S.D.; Taylor, S.M.; Woodruff, T.M. Complement factors C3a and C5a have distinct hemodynamic effects in the rat. Int. Immunopharmacol. 2009, 9, 800–806. [Google Scholar] [CrossRef]
- Damas, J. Involvement of platelet-activating factor in the hypotensive response to zymosan in rats. J. Lipid Mediat. 1991, 3, 333–344. [Google Scholar]
- Smedegard, G.; Cui, L.X.; Hugli, T.E. Endotoxin-induced shock in the rat. A role for C5a. Am. J. Pathol. 1989, 135, 489–497. [Google Scholar] [PubMed]
- Chanan-Khan, A.; Szebeni, J.; Savay, S.; Liebes, L.; Rafique, N.M.; Alving, C.R.; Muggia, F.M. Complement activation following first exposure to pegylated liposomal doxorubicin (Doxil): Possible role in hypersensitivity reactions. Ann. Oncol. 2003, 14, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Kozma, G.T.; Meszaros, T.; Vashegyi, I.; Fulop, T.; Orfi, E.; Dezsi, L.; Rosivall, L.; Bavli, Y.; Urbanics, R.; Mollnes, T.E.; et al. Pseudo-anaphylaxis to Polyethylene Glycol (PEG)-Coated Liposomes: Roles of Anti-PEG IgM and Complement Activation in a Porcine Model of Human Infusion Reactions. ACS Nano 2019, 13, 9315–9324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, R.C.; MacDonald, R.I.; Menco, B.P.; Takeshita, K.; Subbarao, N.K.; Hu, L.R. Small-volume extrusion apparatus for preparation of large, unilamellar vesicles. Biochim. Biophys. Acta 1991, 1061, 297–303. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dézsi, L.; Mészáros, T.; Őrfi, E.; Fülöp, T.G.; Hennies, M.; Rosivall, L.; Hamar, P.; Szebeni, J.; Szénási, G. Complement Activation-Related Pathophysiological Changes in Anesthetized Rats: Activator-Dependent Variations of Symptoms and Mediators of Pseudoallergy. Molecules 2019, 24, 3283. https://doi.org/10.3390/molecules24183283
Dézsi L, Mészáros T, Őrfi E, Fülöp TG, Hennies M, Rosivall L, Hamar P, Szebeni J, Szénási G. Complement Activation-Related Pathophysiological Changes in Anesthetized Rats: Activator-Dependent Variations of Symptoms and Mediators of Pseudoallergy. Molecules. 2019; 24(18):3283. https://doi.org/10.3390/molecules24183283
Chicago/Turabian StyleDézsi, László, Tamás Mészáros, Erik Őrfi, Tamás G. Fülöp, Mark Hennies, László Rosivall, Péter Hamar, János Szebeni, and Gábor Szénási. 2019. "Complement Activation-Related Pathophysiological Changes in Anesthetized Rats: Activator-Dependent Variations of Symptoms and Mediators of Pseudoallergy" Molecules 24, no. 18: 3283. https://doi.org/10.3390/molecules24183283
APA StyleDézsi, L., Mészáros, T., Őrfi, E., Fülöp, T. G., Hennies, M., Rosivall, L., Hamar, P., Szebeni, J., & Szénási, G. (2019). Complement Activation-Related Pathophysiological Changes in Anesthetized Rats: Activator-Dependent Variations of Symptoms and Mediators of Pseudoallergy. Molecules, 24(18), 3283. https://doi.org/10.3390/molecules24183283