
New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.)

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Characterization of the Polyphenolic Extract
2.2. Microencapsulation Efficiency and Characterization of the Powders
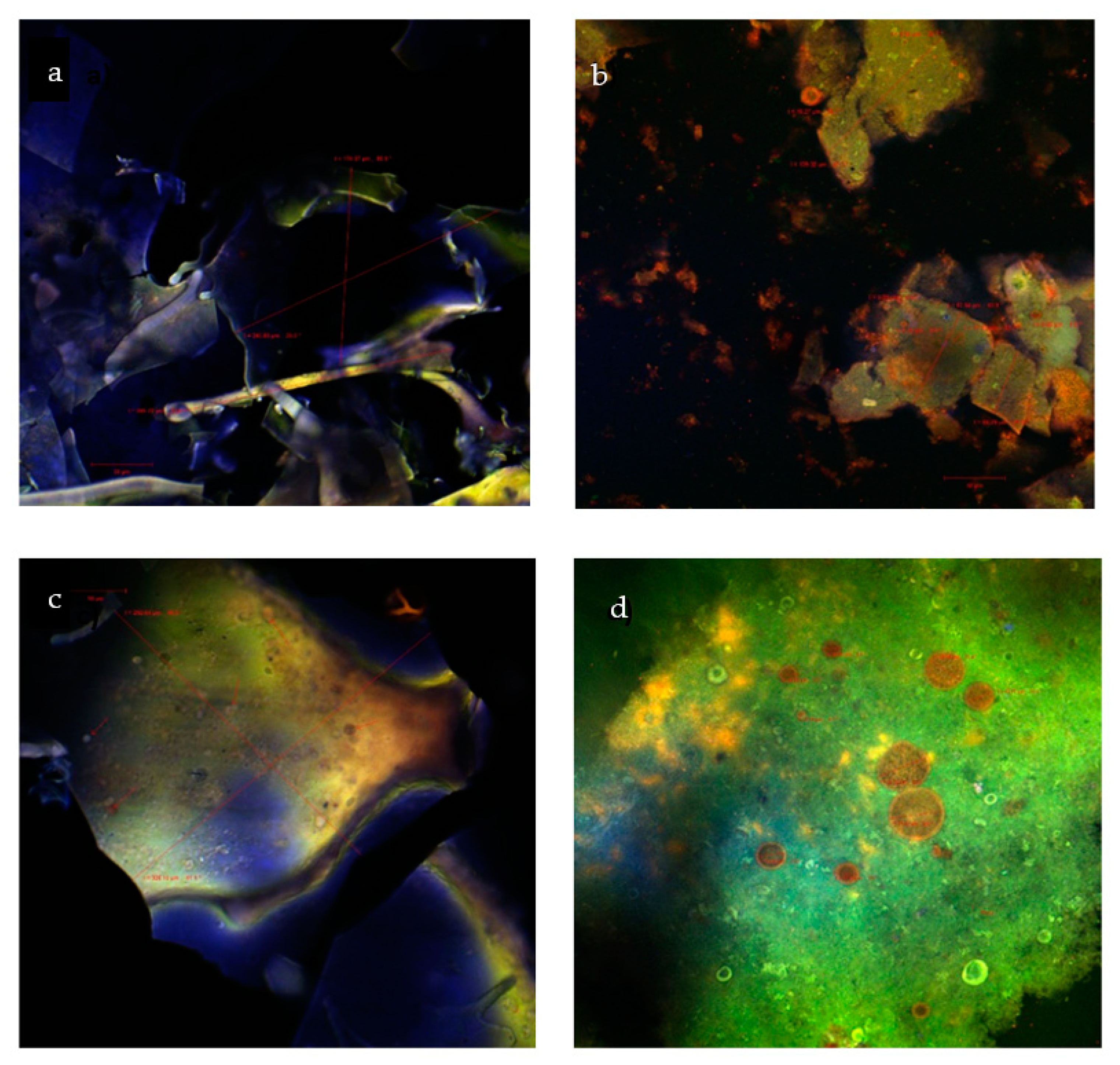
2.3. Morphological Structure of the Microencapsulated Powders
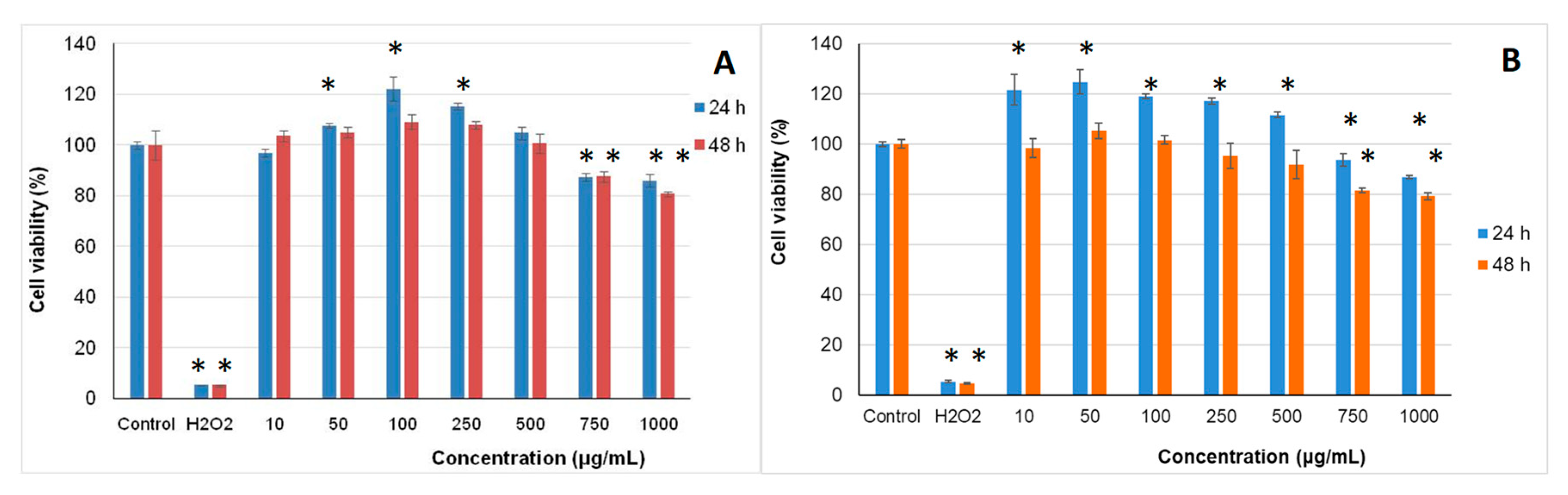
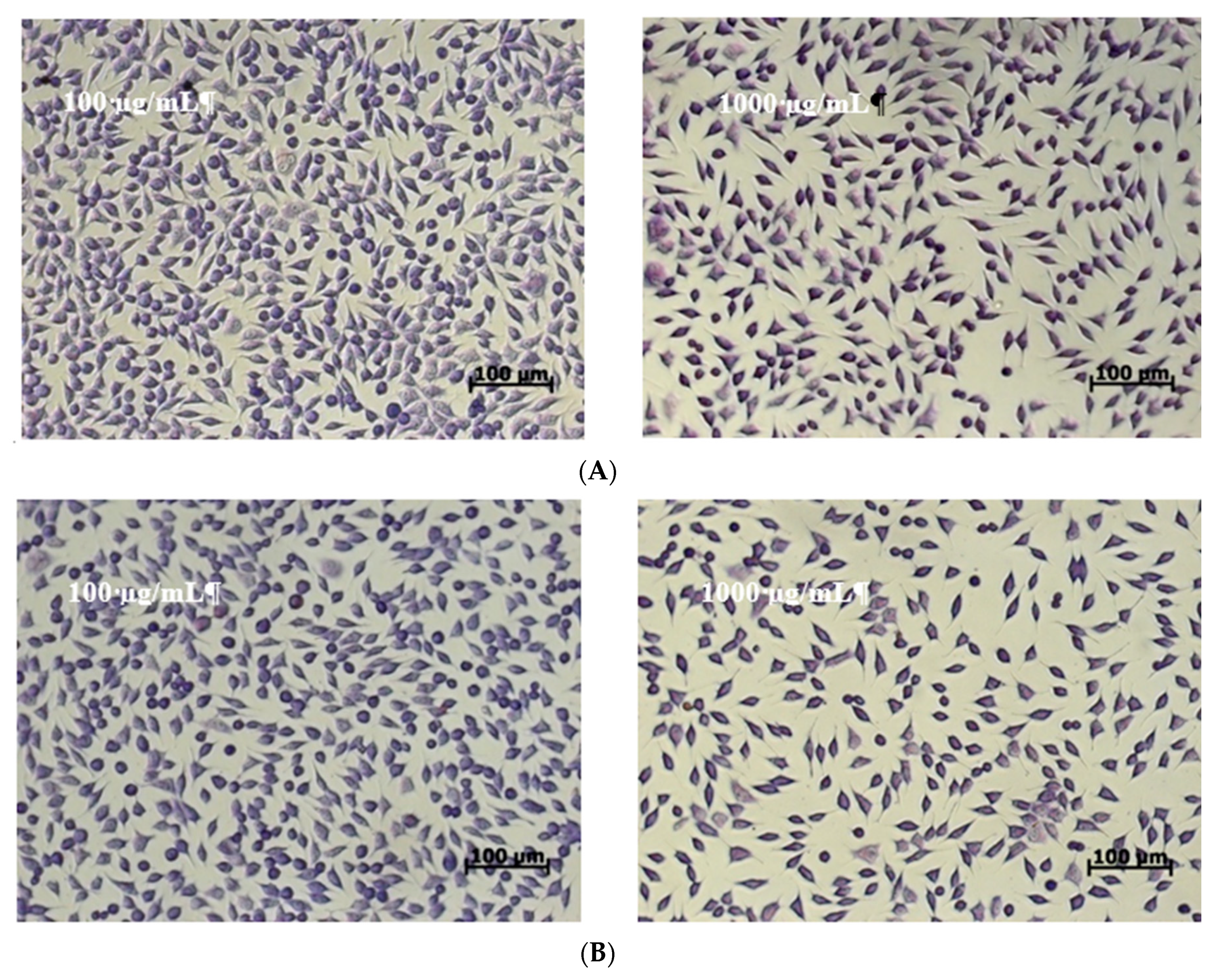
2.4. In Vitro Cytotoxicity of the Microencapsulated Powders
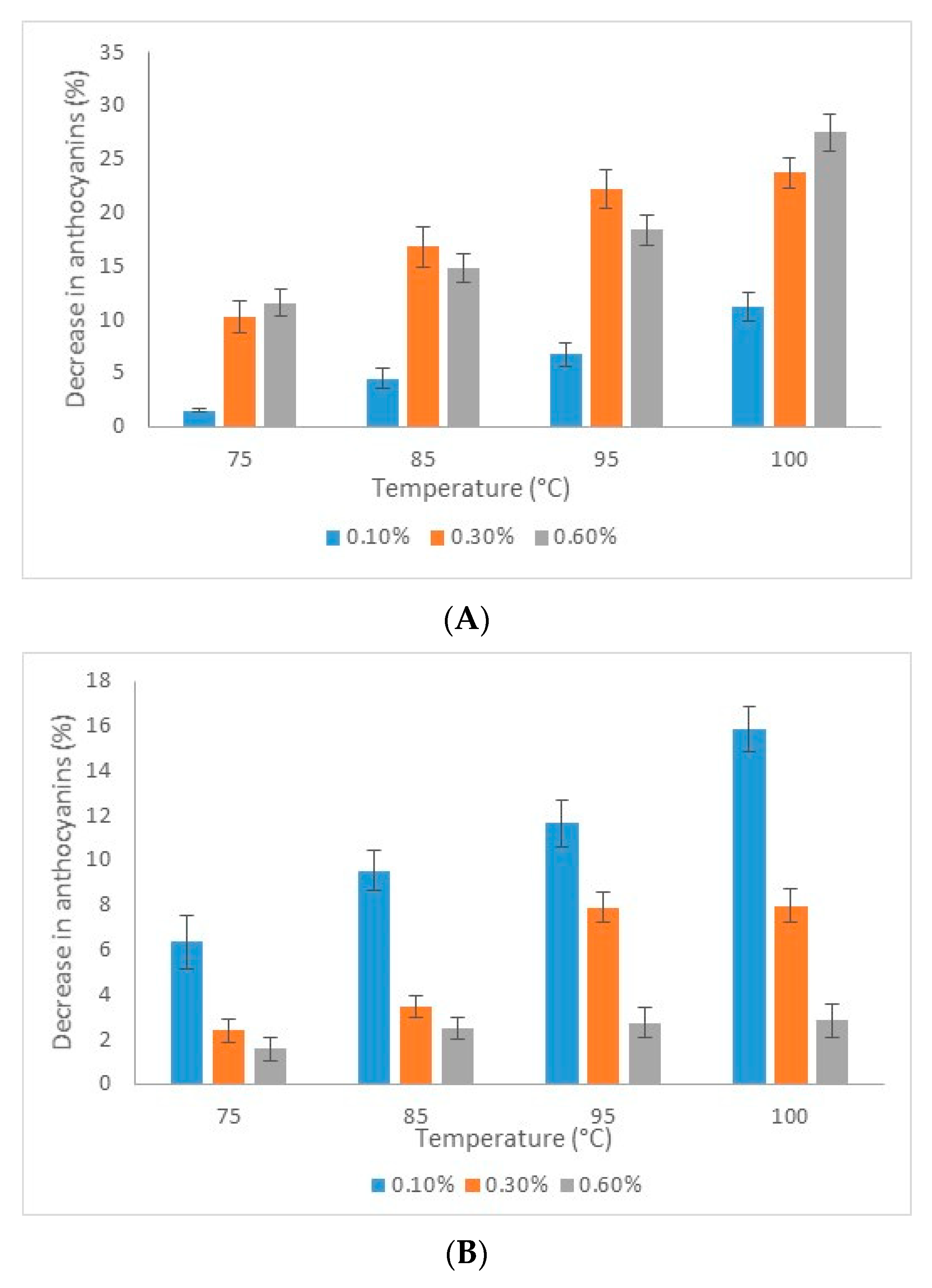
2.5. Thermal Stability of Anthocyanins in Microencapsulated Powder
2.6. Formulation of Anthocyanins-Rich Pastry Cream
3. Materials and Methods
3.1. Materials
3.2. Phytochemicals Extraction and Characterization
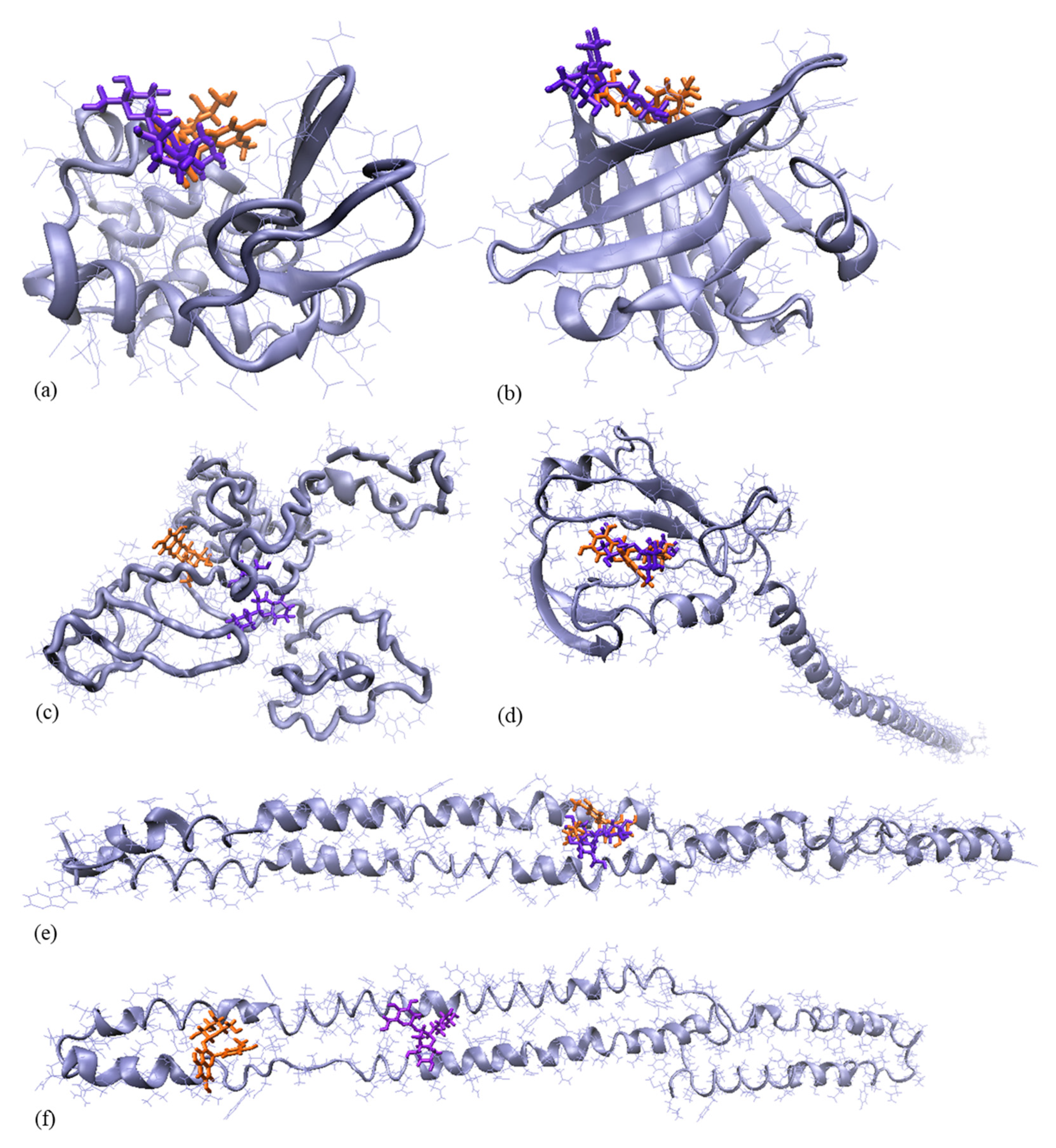
3.3. Molecular Modelling Investigations
3.4. Microencapsulation of Anthocyanins
3.5. Powder Characterization
3.6. Confocal Laser Scanning Microscopy
3.7. Cytocompatibility Testing of the Microencapsulated Powders
3.7.1. Cell Culture and Treatment
3.7.2. Cell Viability
3.7.3. Cell Morphology
3.8. Formulation of a Pastry Model Food (Cream for Cakes)
3.9. Statistical Analysis of Data
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shao, Y.; Hu, Z.; Yu, Y.; Mou, R.; Zhu, Z.; Beta, T. Phenolic acids, anthocyanins, proanthocyanidins, antioxidant activity, minerals and their correlations in non-pigmented, red, and black rice. Food Chem. 2018, 239, 733–741. [Google Scholar] [CrossRef]
- Shao, Y.F.; Xu, F.F.; Sun, X.; Bao, J.S.; Beta, T. Identification and quantification of phenolic acids and anthocyanins as antioxidants in bran, embryo and endosperm of white, red and black rice kernels (Oryza sativa L.). J. Cereal Sci. 2014, 59, 211–218. [Google Scholar] [CrossRef]
- Deng, G.F.; Xu, X.R.; Guo, Y.J.; Xia, E.Q.; Li, S. Determination of antioxidant property and their lipophilic and hydrophilic phenolic contents in cereal grains. J. Funct. Foods 2012, 4, 906–914. [Google Scholar] [CrossRef]
- Deng, G.F.; Xu, X.R.; Zhang, Y.; Li, D.; Gan, R.Y.; Li, H.B. Phenolic compounds and bioactivities of pigmented rice. Crit. Rev. Food Sci. Nutr. 2013, 53, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Qin, P.; Zhang, Y.; Cui, S.; Ren, G. Identification of anthocyanins isolated from black rice (Oryza sativa L.) and their degradation kinetics. Food Res. Int. 2013, 50, 691–697. [Google Scholar] [CrossRef]
- Dipti, S.S.; Bergman, C.; Indrasari, S.D.; Herath, T.; Hall, R.; Lee, H.; Habibi, F.; Bassinello, P.Z.; Graterol, E.; Ferraz, J.P.; et al. The potential of rice to offer solutions for malnutrition and chronic diseases. Rice 2012, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Samyor, D.; Das, A.B.; Deka, S.C. Pigmented rice a potential source of bioactive compounds: A review. Int. J. Food Sci. Technol. 2017, 52, 1073–1081. [Google Scholar] [CrossRef]
- Papillo, V.A.; Locatelli, M.; Travaglia, F.; Bordiga, M.; Garino, C.; Arlorio, M.; Coïsson, J.D. Spray-dried polyphenolic extract from Italian black rice (Oryza sativa L., var. Artemide) as new ingredient for bakery products. Food Chem. 2018, 269, 603–609. [Google Scholar] [CrossRef]
- Cavalcanti, R.N.; Santos, D.T.; Meireles, M.A.A. Non-thermal stabilization mechanisms of anthocyanins in model and food systems – An overview. Food Res. Int. 2011, 44, 499–509. [Google Scholar] [CrossRef]
- Mahdavi, S.A.; Jafari, S.M.; Assadpoor, E.; Dehnad, D. Microencapsulation optimization of natural anthocyanins with maltodextrin, gum Arabic and gelatin. Int. J. Biol. Macromol. 2016, 85, 379–385. [Google Scholar] [CrossRef]
- Kuang, S.S.; Oliveira, J.C.; Crean, A.M. Microencapsulation as a tool for incorporating bioactive ingredients into food. Crit. Rev. Food Sci. Nutr. 2010, 50, 951–968. [Google Scholar] [CrossRef] [PubMed]
- Nontasan, S.; Moongngarm, A.; Deeseenthum, S. Application of functional colourant prepared from black rice bran in yogurt. APCBEE Procedia 2012, 2, 62–67. [Google Scholar] [CrossRef]
- Santos, D.T.; Albarelli, J.Q.; Beppu, M.M.; Meireles, M.A.A. Stabilization of anthocyanin extract from jabuticaba skins by encapsulation using supercritical CO2 as solvent. Food Res. Int. 2013, 50, 617–624. [Google Scholar] [CrossRef]
- Stănciuc, N.; Turturică, M.; Oancea, A.-M.; Barbu, V.; Ioniță, E.; Aprodu, I.; Râpeanu, G. Microencapsulation of anthocyanins from grapes skins by whey proteins isolates and different polymers. Food Bioproc. Technol. 2017, 10, 1715–1726. [Google Scholar]
- Rawel, H.M.; Kroll, J.; Hohl, U.C. Model studies on reaction of plant phenols with whey proteins. Nahrung 2001, 45, 72–81. [Google Scholar] [CrossRef]
- Viljanen, K.; Kivikari, R.; Heinonen, M. Protein-lipid interactions during liposome oxidation with added anthocyanin and other phenolic compounds. J. Agric. Food Chem. 2004, 52, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.F.; Brodkorb, A. The casein micelle: Historical aspects, current concepts and significance. Int. Dairy J. 2008, 18, 677–684. [Google Scholar] [CrossRef]
- Schokker, E.P.; Church, J.S.; Mata, J.P.; Gilbert, E.P.; Puvanenthiran, A.; Udabage, P. Reconstitution properties of micellar casein powder: effects of composition and storage. Int. Dairy J. 2011, 21, 877–886. [Google Scholar] [CrossRef]
- Banjerdpongchai, R.; Wudtiwai, B.; Sringarm, K. Cytotoxic and apoptotic-inducing effects of purple rice extracts and chemotherapeutic drugs on human cancer cell lines. Asian Pac. J. Cancer Prev. 2013, 14, 6541–6548. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.P.; O′Donnell, C.; Tiwari, B.K. Effect of thermal processing on anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends Food Sci. Technol. 2010, 21, 3–11. [Google Scholar] [CrossRef]
- Turturică, M.; Stănciuc, N.; Bahrim, G.; Râpeanu, R. Effect of thermal treatment on phenolic compounds from plum (Prunus domestica) extracts – A kinetic study. J. Food Eng. 2016, 171, 200–207. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: a hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Chrysina, E.D.; Brew, K.; Acharya, K.R. Crystal structures of apo-and holo-bovine α-lactalbumin at 2.2-Å resolution reveal an effect of calcium on inter-lobe interactions. J. Biol. Chem. 2000, 275, 37021–37029. [Google Scholar] [CrossRef] [PubMed]
- Loch, J.; Polit, A.; Górecki, A.; Bonarek, P.; Kurpiewska, K.; Dziedzicka-Wasylewska, M.; Lewiński, K. Two modes of fatty acid binding to bovine β-lactoglobulin-crystallographic and spectroscopic studies. J. Mol. Recog. 2011, 24, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inform. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Krissinel, E. Crystal contacts as nature’s docking solutions. J. Comput. Chem. 2010, 31, 133–143. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD - Visual Molecular Dynamics. J. Mol. Grap. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Oancea, A.-M.; Hasan, M.; Vasile, A.M.; Barbu, V.; Enachi, E.; Bahrim, G.; Râpeanu, G.; Stănciuc, N. Functional evaluation of microencapsulated anthocyanins from sour cherries skins extract in whey proteins isolate. LWT – Food Sci. Technol. 2018, 95, 129–134. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. (Eds.) Anthocyanins. Characterization and measurement with UV-visible spectroscopy. In Current Protocols in Food Analytic Chemistry; John Wiley & Sons: New York, NY, USA, 2001; Volume 1, pp. 137–139. [Google Scholar]
- Saénz, C.; Tapia, S.; Chávez, J.; Robert, P. Microencapsulation by spray drying of bioactive compounds from cactus pear (Opuntia ficus-indica). Food Chem. 2009, 114, 616–622. [Google Scholar] [CrossRef]
- Crăciunescu, O.; Gaspar, A.; Toma, L.; Utoiu, E.; Moldovan, L. Evaluation of antioxidant and cytoprotective activities of Arnica montana L. and Artemisia absinthium L. ethanolic extracts. Chem. Central J. 2012, 6, 97–107. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein-Anthocyanins Complex | Interaction Energy, kJ/mol | Contact Area, Å2 | ΔGint, kcal/mol | ΔGdiss, kcal/mol | Residues Involved in Hydrophobic Contacts with the Anthocyanins | Residues Involved in Hb with the Anthocyanins |
|---|---|---|---|---|---|---|
| αLA-C3G | −172.59 | 560.70 | −0.1 | 2.4 | Thr33, Glu49, Phe53, Ile59, Asn102, Tyr103, Trp104, Ala106, Lys108 | Thr33, Asn102, Ala106 |
| βLG-C3G | −197.49 | 539.10 | −0.3 | 2.6 | Lys69, Ile71, Asp85, Ala86, Asn88, Asn90, Met107 | Lys69, Ala86 |
| βCN-C3G | −154.90 | 643.30 | −0.5 | 2.8 | Ile41, Lys43, Lys44, Ile45, Gln104, Pro105, Glu106, Val107, Pro165, Pro167, Leu180 | Lys43, Lys44 |
| αS1CN-C3G | −170.33 | 507.00 | −0.8 | 3.1 | Ser56, Ile59, Gly60, Glu62, Ser63, Leu157, Tyr161, Leu164, Phe165 | Gly60 |
| αS2CN-C3G | −187.69 | 561.20 | −0.6 | 2.9 | Leu114, Leu176, Asn177, Leu179, Ile182, Ser183, Gln187 | Gln187 |
| kCN-C3G | −181.25 | 677.50 | 0.0 | 2.3 | Ala106, Lys107, Ser108, His119, Pro120, His123, Thr154, Pro171, Glu172, Val173, Glu175, Ser176, Pro177 | Lys107, His123, Thr154, Glu172, Glu175 |
| αLA-P3G | −179.98 | 487.60 | −0.5 | 2.0 | His32, Thr33, Val42, Asn44, Glu49, Tyr103, Trp104, Leu105, Ala106 | Tyr103 |
| βLG-P3G | −142.36 | 497.10 | −0.1 | 1.6 | Leu39, Val41, Leu58, Lys69, Ile71, Ile84, Asp85, Ala86, Leu87, Asn90, Glu108, Asn109 | Asp85, Ala86, Glu108, Asn109 |
| βCN-P3G | −232.18 | 676.20 | −0.3 | 1.8 | Lys63, His65, Tyr75, Pro76, Pro78, Glu106, Val107, Met108, Gly109, Val110, Ser111, Lys112, Leu155, Gln156, Met159, His160, Gln161 | His65, Ser111, Gln156 |
| S1CN-P3G | −219.83 | 485.80 | −1.1 | 2.5 | Ser56, Ile59, Gly60, Glu62, Ser63, Glu65, Leu157, Tyr161, Leu164, Phe165 | Glu65 |
| S2CN-P3G | −136.75 | 484.70 | −0.7 | 2.2 | Leu121, Pro123, Asp125, Val127, Arg129, Asn130, Val154, Glu157, Ser158 | - |
| kCN-P3G | −200.86 | 656.10 | −0.4 | 1.8 | Pro105, Ala106, Lys107, Ser108, His119, Pro120, His123, Thr154, Asp169, Pro171, Glu172, Val173, Ile174, Glu175, Ser176 | Ser108, His123, Asp169, Glu172, Ser176 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aprodu, I.; Milea, Ș.A.; Anghel, R.-M.; Enachi, E.; Barbu, V.; Crăciunescu, O.; Râpeanu, G.; Bahrim, G.E.; Oancea, A.; Stănciuc, N. New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.). Molecules 2019, 24, 3389. https://doi.org/10.3390/molecules24183389
Aprodu I, Milea ȘA, Anghel R-M, Enachi E, Barbu V, Crăciunescu O, Râpeanu G, Bahrim GE, Oancea A, Stănciuc N. New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.). Molecules. 2019; 24(18):3389. https://doi.org/10.3390/molecules24183389
Chicago/Turabian StyleAprodu, Iuliana, Ștefania Adelina Milea, Roxana-Mădălina Anghel, Elena Enachi, Vasilica Barbu, Oana Crăciunescu, Gabriela Râpeanu, Gabriela Elena Bahrim, Anca Oancea, and Nicoleta Stănciuc. 2019. "New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.)" Molecules 24, no. 18: 3389. https://doi.org/10.3390/molecules24183389
APA StyleAprodu, I., Milea, Ș. A., Anghel, R. -M., Enachi, E., Barbu, V., Crăciunescu, O., Râpeanu, G., Bahrim, G. E., Oancea, A., & Stănciuc, N. (2019). New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.). Molecules, 24(18), 3389. https://doi.org/10.3390/molecules24183389