
Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems—Fate and Microbial Responses

 , ,
, ,
Abstract
:
1. Introduction
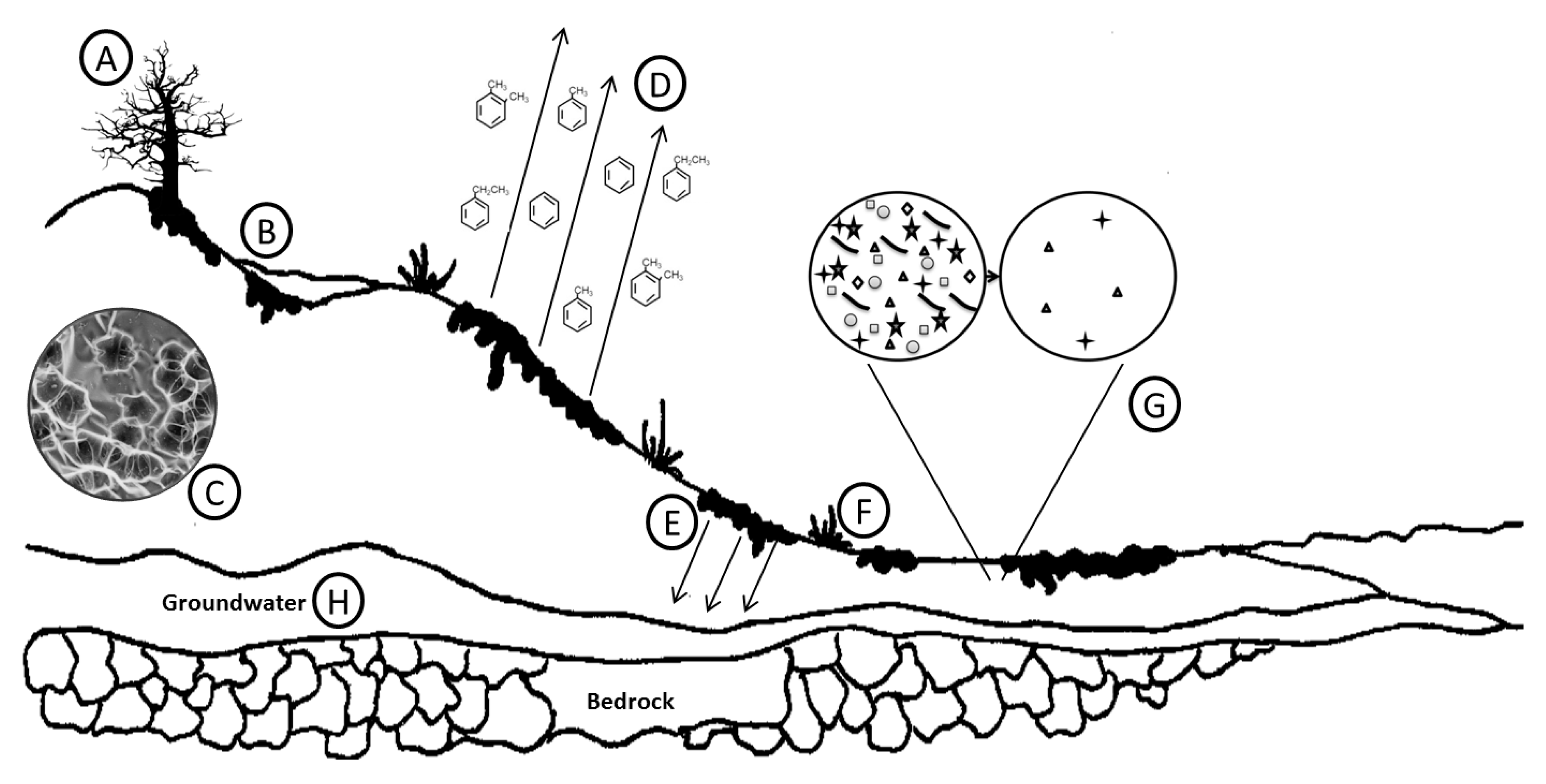
2. Oil Composition Changes upon Entering the Environment
2.1. Volatilization
2.2. Dissolution
2.3. Sorption and Desorption
3. Toxicity of Hydrocarbons to Microorganisms and Microbial Communities
- direct toxicity of the chemical compounds,
- the locking away of nutrients and water, preventing microorganisms from obtaining essential building blocks for proliferation,
- difficulty in adjusting to the highly nonpolar conditions, which can cause microbial cells to rupture by dissolving the cytoplasm membrane lipids [20].
- The hydrocarbon fraction itself: In saturated hydrocarbons, the chain length is highly correlated with direct toxicity. Fractions of lower molecular weight structures (C6 to C20) have been shown to be more toxic due to their high bioavailability; however, larger saturated chained hydrocarbons have increased mutagenic potential [5]. The toxicity of unsaturated hydrocarbon structures is not as predictable and may be influenced by reactive functional groups, water and membrane solubility, viscosity, and interactions of these compounds with the membrane and with membrane constituents.
- The concentration of nutrients: the diversity of the microbial communities in soil with a higher concentration of N and P has been shown to increase the abundance and activity of the hydrocarbon-degrading microbial community [22].
- Co-contamination with other chemicals (i.e., heavy metals): higher toxicity of petroleum hydrocarbon to soil microorganisms was reported with the presence of heavy metals as a co-contaminant when compared with the toxicity of petroleum hydrocarbon as the only contaminant [27]. Khudur et al. (2019) showed that hydrocarbon-contaminated soils were more recalcitrant to microbial bioremediation when co-contaminated with lead (Pb). Lead inhibits many metabolic pathways, such as the enzymatic and respiratory processes of many bacteria, and creates additional stress to hydrocarbon-degrading species [28].
4. Physiochemical Factors Affecting Natural Attenuation of Petrogenic Hydrocarbons
4.1. Nutrients and Additives
4.2. Salinity
4.3. Drought/Rainfall/Moisture
5. Microbe–Microbe Interactions
6. Microbial Hydrocarbon Utilization
6.1. Aerobic Hydrocarbon Catabolism
6.2. Anaerobic Hydrocarbon Catabolism
6.2.1. Addition of Fumarate to the Hydrocarbon Chain
6.2.2. Oxygen Independent Hydroxylation of the Hydrocarbon Chain
6.2.3. Carboxylation of Aromatics
6.2.4. Hydration of Alkenes/Alkynes
6.2.5. Reverse Methanogenesis
6.3. Syntrophy
6.4. Microbial Interactions That Can Indirectly Aid Hydrocarbon Degradation
7. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chikere, C.B.; Okpokwasili, G.C.; Chikere, B.O. Monitoring of microbial hydrocarbon remediation in the soil. 3 Biotech 2011, 1, 117–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, M.; Wang, Y.; Yu, J.; Xiao, M.; Jiang, L.; Yang, J.; Fang, C.; Chen, J.; Li, B. Understanding Plant-Microbe Interactions for Phytoremediation of Petroleum-Polluted Soil. PLoS ONE 2011, 6, e17961. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.C. Anthropogenic transformation of the terrestrial biosphere. Philosophical Transactions of the Royal Society A: Mathematical. Phys. Eng. Sci. 2011, 369, 1010–1035. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Lu, X.; Sun, Q.; Zhu, W. Aging effect of petroleum hydrocarbons in soil under different attenuation conditions. Agric. Ecosyst. Environ. 2012, 149, 109–117. [Google Scholar] [CrossRef]
- Bak, F.; Bonnichsen, L.; Jørgensen, N.O.G.; Nicolaisen, M.H.; Nybroe, O. The biosurfactant viscosin transiently stimulates n-hexadecane mineralization by a bacterial consortium. Appl. Microbiol. Biotechnol. 2015, 99, 1475–1483. [Google Scholar] [CrossRef]
- Colwell, R.R.; Walker, J.D.; Cooney, J.J. Ecological Aspects of Microbial Degradation of Petroleum in the Marine Environment. CRC Crit. Rev. Microbiol. 1977, 5, 423–445. [Google Scholar] [CrossRef]
- Boehm, P.D.; Fiest, D.L.; Mackay, D.; Paterson, S. Physical-chemical weathering of petroleum hydrocarbons from the IXTOC I blowout: chemical measurements and a weathering model. Environ. Sci. Technol. 1982, 16, 498–505. [Google Scholar] [CrossRef]
- Zhang, Z.; Gai, L.; Hou, Z.; Yang, C.; Ma, C.; Wang, Z.; Sun, B.; He, X.; Tang, H.; Xu, P. Characterization and biotechnological potential of petroleum-degrading bacteria isolated from oil-contaminated soils. Bioresour. Technol. 2010, 101, 8452–8456. [Google Scholar] [CrossRef]
- Van Metre, P.C.; Majewski, M.S.; Mahler, B.J.; Foreman, W.T.; Braun, C.L.; Wilson, J.T.; Burbank, T.L. Volatilization of polycyclic aromatic hydrocarbons from coal-tar-sealed pavement. Chemosphere 2012, 88, 1–7. [Google Scholar] [CrossRef]
- Essaid, H.I.; Bekins, B.A.; Herkelrath, W.N.; Delin, G.N. Crude Oil at the Bemidji Site: 25 Years of Monitoring, Modeling, and Understanding. Groundwater 2011, 49, 706–726. [Google Scholar] [CrossRef] [PubMed]
- De Hemptinne, J.C.; Delepine, H.; Jose, C.; Jose, J. Aqueous Solubility of Hydrocarbon Mixtures. Revue de l’Institut Français du Pétrole 1998, 53, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Lundegard, P.D.; Sweeney, R.E. Total Petroleum Hydrocarbons in Groundwater—Evaluation of Nondissolved and Nonhydrocarbon Fractions. Environ. Forensics 2004, 5, 85–95. [Google Scholar] [CrossRef]
- Landmeyer, J.E.; Effinger, T.N. Effect of phytoremediation on concentrations of benzene, toluene, naphthalene, and dissolved oxygen in groundwater at a former manufactured gas plant site, Charleston, South Carolina, USA, 1998–2014. Environ. Earth Sci. 2016, 75, 605. [Google Scholar] [CrossRef]
- Regitano, J.B.; Koskinen, W.C.; Sadowsky, M.J. Influence of Soil Aging on Sorption and Bioavailability of Simazine. J. Agric. Food Chem. 2006, 54, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Providenti, M.A.; Lee, H.; Trevors, J.T. Selected factors limiting the microbial degradation of recalcitrant compounds. J. Ind. Microbiol. Biotechnol. 1993, 12, 379–395. [Google Scholar] [CrossRef]
- Khan, S.; Afzal, M.; Iqbal, S.; Khan, Q.M. Plant–bacteria partnerships for the remediation of hydrocarbon contaminated soils. Chemosphere 2013, 90, 1317–1332. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Manna, S.; Pandiyan, K.; Singh, A.; Kumar, M.; Chakdar, H.; Kashyap, P.L.; Srivastava, A.K. Isolation and characterization of biosurfactant producing Bacillus sp. from diesel fuel-contaminated site. Microbiology 2016, 85, 56–62. [Google Scholar] [CrossRef]
- Ball, A.; Truskewycz, A. Polyaromatic hydrocarbon exposure: an ecological impact ambiguity. Environ. Sci. Pollut. Res. 2013, 20, 4311–4326. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Juottonen, H.; Siivonen, P.; Quesada, C.L.; Tuomi, P.; Pulkkinen, P.; Yrjälä, K. Spatial patterns of microbial diversity and activity in an aged creosote-contaminated site. ISME J. 2014, 8, 2131–2142. [Google Scholar] [CrossRef] [Green Version]
- Zengler, K.; Palsson, B.O. A road map for the development of community systems (CoSy) biology. Nat. Rev. Genet. 2012, 10, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Khudur, L.S.; Shahsavari, E.; Miranda, A.F.; Morrison, P.D.; Nugegoda, D.; Ball, A.S. Evaluating the efficacy of bioremediating a diesel-contaminated soil using ecotoxicological and bacterial community indices. Environ. Sci. Pollut. Res. 2015, 22, 14809–14819. [Google Scholar] [CrossRef] [PubMed]
- van Dorst, J.; Siciliano, S.D.; Winsley, T.; Snape, I.; Ferrari, B.C. Bacterial Targets as Potential Indicators of Diesel Fuel Toxicity in Subantarctic Soils. Appl. Environ. Microbiol. 2014, 80, 4021. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Sinkko, H.; Penttinen, P.; Lindström, K. Characterization of successional changes in bacterial community composition during bioremediation of used motor oil-contaminated soil in a boreal climate. Sci. Total. Environ. 2016, 542, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semple, K.T.; Doick, K.J.; Wick, L.Y.; Harms, H. Microbial interactions with organic contaminants in soil: Definitions, processes and measurement. Environ. Pollut. 2007, 150, 166–176. [Google Scholar] [CrossRef]
- Johnsen, A.R.; Wick, L.Y.; Harms, H. Principles of microbial PAH-degradation in soil. Environ. Pollut. 2005, 133, 71–84. [Google Scholar] [CrossRef]
- Thavamani, P.; Malik, S.; Beer, M.; Megharaj, M.; Naidu, R. Microbial activity and diversity in long-term mixed contaminated soils with respect to polyaromatic hydrocarbons and heavy metals. J. Environ. Manag. 2012, 99, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Khudur, L.S.; Shahsavari, E.; Webster, G.T.; Nugegoda, D.; Ball, A.S. The impact of lead co-contamination on ecotoxicity and the bacterial community during the bioremediation of total petroleum hydrocarbon-contaminated soils. Environ. Pollut. 2019, 253, 939–948. [Google Scholar] [CrossRef]
- Sutton, N.B.; Maphosa, F.; Morillo, J.A.; Al-Soud, W.A.; Langenhoff, A.A.M.; Grotenhuis, T.; Rijnaarts, H.H.M.; Smidt, H. Impact of Long-Term Diesel Contamination on Soil Microbial Community Structure. Appl. Environ. Microbiol. 2013, 79, 619. [Google Scholar] [CrossRef]
- Polz, M.F.; Alm, E.J.; Hanage, W.P. Horizontal Gene Transfer and the Evolution of Bacterial and Archaeal Population Structure. Trends Genet. 2013, 29, 170–175. [Google Scholar] [CrossRef]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in multispecies biofilms: do they actually matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Coulon, F.; Pelletier, E.; Gourhant, L.; Delille, D. Effects of nutrient and temperature on degradation of petroleum hydrocarbons in contaminated sub-Antarctic soil. Chemosphere 2005, 58, 1439–1448. [Google Scholar] [CrossRef]
- Bento, F.M.; Camargo, F.A.; Okeke, B.C.; Frankenberger, W.T. Comparative bioremediation of soils contaminated with diesel oil by natural attenuation, biostimulation and bioaugmentation. Bioresour. Technol. 2005, 96, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Plaza, C.; Xing, B.; Fernández, J.M.; Senesi, N.; Polo, A. Binding of polycyclic aromatic hydrocarbons by humic acids formed during composting. Environ. Pollut. 2009, 157, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.F.; Peixoto, R.S. Biodegradation of petroleum hydrocarbons in hypersaline environments. Braz. J. Microbiol. 2012, 43, 865–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathepure, B.Z. Recent studies in microbial degradation of petroleum hydrocarbons in hypersaline environments. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Gao, Y.-C.; Wang, J.-N.; Guo, S.-H.; Hu, Y.-L.; Li, T.-T.; Mao, R.; Zeng, D.-H. Effects of salinization and crude oil contamination on soil bacterial community structure in the Yellow River Delta region. China. Appl. Soil Ecol. 2015, 86, 165–173. [Google Scholar] [CrossRef]
- Riis, V.; Kleinsteuber, S.; Babel, W. Influence of high salinities on the degradation of diesel fuel by bacterial consortia. Can. J. Microbiol. 2003, 49, 713–721. [Google Scholar] [CrossRef]
- Kleinsteuber, S.; Riis, V.; Fetzer, I.; Harms, H.; Müller, S. Population Dynamics within a Microbial Consortium during Growth on Diesel Fuel in Saline Environments†. Appl. Environ. Microbiol. 2006, 72, 3531–3542. [Google Scholar] [CrossRef]
- Oren, A. Microbial life at high salt concentrations: phylogenetic and metabolic diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef]
- Al-Mailem, D.; Eliyas, M.; Radwan, S. Bioremediation of oily hypersaline soil and water via potassium and magnesium amendment. Can. J. Microbiol. 2013, 59, 837–844. [Google Scholar] [CrossRef]
- Rhykerd, R.L.; Weaver, R.W.; McInnes, K.J. Influence of salinity on bioremediation of oil in soil. Environ. Pollut. 1995, 90, 127–130. [Google Scholar] [CrossRef]
- Lorenz, M.G.; Wackernagel, W. Bacterial gene transfer by natural genetic transformation in the environment. Microbiol. Rev. 1994, 58, 563–602. [Google Scholar]
- Poté, J.; Ceccherini, M.T.; Van, V.T.; Rosselli, W.; Wildi, W.; Simonet, P.; Vogel, T.M. Fate and transport of antibiotic resistance genes in saturated soil columns. Eur. J. Soil Boil. 2003, 39, 65–71. [Google Scholar] [CrossRef]
- Sheik, C.S.; Beasley, W.H.; Elshahed, M.S.; Zhou, X.; Luo, Y.; Krumholz, L.R. Effect of warming and drought on grassland microbial communities. ISME J. 2011, 5, 1692–1700. [Google Scholar] [CrossRef] [Green Version]
- Hueso, S.; García, C.; Hernández, T. Severe drought conditions modify the microbial community structure, size and activity in amended and unamended soils. Soil Biol. Biochem. 2012, 50, 167–173. [Google Scholar] [CrossRef]
- Phillips, L.A.; Greer, C.W.; Farrell, R.; Germida, J.J. Field-scale assessment of weathered hydrocarbon degradation by mixed and single plant treatments. Appl. Soil Ecol. 2009, 42, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Barnard, R.L.; A Osborne, C.; Firestone, M.K. Responses of soil bacterial and fungal communities to extreme desiccation and rewetting. ISME J. 2013, 7, 2229–2241. [Google Scholar] [CrossRef]
- Taccari, M.; Milanovic, V.; Comitini, F.; Casucci, C.; Ciani, M. Effects of biostimulation and bioaugmentation on diesel removal and bacterial community. Int. Biodeterior. Biodegrad. 2012, 66, 39–46. [Google Scholar] [CrossRef]
- Davis, C.; Cort, T.; Dai, D.; Illangasekare, T.H.; Munakata-Marr, J. Effects of heterogeneity and experimental scale on the biodegradation of diesel. Biodegradation 2003, 14, 373–384. [Google Scholar] [CrossRef]
- Lahel, A.; Fanta, A.B.; Sergienko, N.; Shakya, M.; López, M.E.; Behera, S.K.; Rene, E.R.; Park, H.-S. Effect of process parameters on the bioremediation of diesel contaminated soil by mixed microbial consortia. Int. Biodeterior. Biodegrad. 2016, 113, 375–385. [Google Scholar] [CrossRef]
- Dheilly, N.M.; Poulin, R.; Thomas, F. Biological warfare: Microorganisms as drivers of host–parasite interactions. Infect. Genet. Evol. 2015, 34, 251–259. [Google Scholar] [CrossRef]
- Sabra, W.; Dietz, D.; Tjahjasari, D.; Zeng, A.-P. Biosystems analysis and engineering of microbial consortia for industrial biotechnology. Eng. Life Sci. 2010, 10, 407–421. [Google Scholar] [CrossRef]
- Thiengmag, S.; Chuencharoen, S.; Thasana, N.; Whangsuk, W.; Jangiam, W.; Mongkolsuk, S.; Loprasert, S. Bacterial consortium expressing surface displayed, intra- and extracellular lipases and pseudopyronine B for the degradation of oil. Int. J. Environ. Sci. Technol. 2016, 13, 2067–2078. [Google Scholar] [CrossRef]
- Varjani, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- Abed, R.M.; Köster, J. The direct role of aerobic heterotrophic bacteria associated with cyanobacteria in the degradation of oil compounds. Int. Biodeterior. Biodegrad. 2005, 55, 29–37. [Google Scholar] [CrossRef]
- Laurie, A.D.; Lloyd-Jones, G. The phn Genes of Burkholderia sp. Strain RP007 Constitute a Divergent Gene Cluster for Polycyclic Aromatic Hydrocarbon Catabolism. J. Bacteriol. 1999, 181, 531–540. [Google Scholar]
- Peng, R.-H.; Xiong, A.-S.; Xue, Y.; Fu, X.-Y.; Gao, F.; Zhao, W.; Tian, Y.-S.; Yao, Q.-H. Microbial biodegradation of polyaromatic hydrocarbons. FEMS Microbiol. Rev. 2008, 32, 927–955. [Google Scholar] [CrossRef] [Green Version]
- Kiyohara, H.; Torigoe, S.; Kaida, N.; Asaki, T.; Iida, T.; Hayashi, H.; Takizawa, N. Cloning and characterization of a chromosomal gene cluster, pah, that encodes the upper pathway for phenanthrene and naphthalene utilization by Pseudomonas putida OUS82. J. Bacteriol. 1994, 176, 2439–2443. [Google Scholar] [CrossRef] [Green Version]
- Sylvestre, M.; Sirois, M.; Hurtubise, Y.; Bergeron, J.; Ahmad, D.; Shareck, F.; Barriault, D.; Guillemette, I.; Juteau, J.M. Sequencing of Comamonas testosteroni strain B-356-biphenyl/chlorobiphenyl dioxygenase genes: evolutionary relationships among Gram-negative bacterial biphenyl dioxygenases. Gene 1996, 174, 195–202. [Google Scholar] [CrossRef]
- Dionisi, H.M.; Chewning, C.S.; Morgan, K.H.; Menn, F.M.; Easter, J.P.; Sayler, G.S. Abundance of dioxygenase genes similar to Ralstonia sp. strain U2 nagAc is correlated with naphthalene concentrations in coal tar-contaminated freshwater sediments. Appl. Environ. Microbiol. 2004, 70, 3988–3995. [Google Scholar] [CrossRef]
- Kim, S.-J.; Kweon, O.; Jones, R.C.; Freeman, J.P.; Edmondson, R.D.; Cerniglia, C.E. Complete and integrated pyrene degradation pathway in Mycobacterium vanbaalenii PYR-1 based on systems biology. J. Bacteriol. 2007, 189, 464–472. [Google Scholar] [CrossRef]
- Habe, H.; Chung, J.-S.; Kato, H.; Ayabe, Y.; Kasuga, K.; Yoshida, T.; Nojiri, H.; Yamane, H.; Omori, T. Characterization of the Upper Pathway Genes for Fluorene Metabolism in Terrabacter sp. Strain DBF63. J. Bacteriol. 2004, 186, 5938–5944. [Google Scholar] [CrossRef] [Green Version]
- Stingley, R.L.; Brezna, B.; Khan, A.A.; Cerniglia, C.E. Novel organization of genes in a phthalate degradation operon of Mycobacterium vanbaalenii PYR-1. Microbiology 2004, 150, 3749–3761. [Google Scholar] [CrossRef] [Green Version]
- Antonio, V. The Proceedings from Halophiles 2013, the International Congress on Halophilic Microorganisms; Frontiers Media SA: Lausanne, Switzerland, 2015. [Google Scholar]
- Vedler, E. Megaplasmids and the Degradation of Aromatic Compounds by Soil Bacteria. In Microbial Megaplasmids; Edward, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 11, pp. 33–53. [Google Scholar]
- Parke, D. Supraoperonic clustering of pca genes for catabolism of the phenolic compound protocatechuate in Agrobacterium tumefaciens. J. Bacteriol. 1995, 177, 3808–3817. [Google Scholar] [CrossRef] [Green Version]
- Daval, S.; Lebreton, L.; Gazengel, K.; Boutin, M.; Guillerm-Erckelboudt, A.; Sarniguet, A. The biocontrol bacterium Pseudomonas fluorescens Pf29Arp strain affects the pathogenesis-related gene expression of the take-all fungus Gaeumannomyces graminis var. tritici on wheat roots. Mol. Plant Pathol. 2011, 12, 839–854. [Google Scholar] [CrossRef]
- Brown, L.M.; Gunasekera, T.S.; Bowen, L.L.; Ruiz, O.N. Draft Genome Sequence of Rhodovulum sp. Strain NI22, a Naphthalene-Degrading Marine Bacterium. Genome Announc. 2015, 3, e01475-14. [Google Scholar] [CrossRef]
- Yeo, C.C.; Wong, M.V.-M.; Feng, Y.; Song, K.P.; Poh, C.L. Molecular characterization of an inducible gentisate 1,2-dioxygenase gene, xlnE, from Pseudomonas alcaligenes NCIMB 9867. Gene 2003, 312, 239–248. [Google Scholar] [CrossRef]
- Papke, R.T. Preface to the proceedings of Halophiles 2013. Front. Microbiol. 2015, 6, 341. [Google Scholar] [CrossRef]
- Vomberg, A.; Klinner, U. Distribution of alkB genes within n-alkane-degrading bacteria. J. Appl. Microbiol. 2000, 89, 339–348. [Google Scholar] [CrossRef]
- Nakamura, R.; Kondo, R.; Shen, M.H.; Ochiai, H.; Hisamatsu, S.; Sonoki, S. Identification of cytochrome P450 monooxygenase genes from the white-rot fungus Phlebia brevispora. AMB Express 2012, 2, 8. [Google Scholar] [CrossRef]
- Zylstra, G.J.; Kim, E.; Goyal, A.K. Comparative Molecular Analysis of Genes for Polycyclic Aromatic Hydrocarbon Degradation. In Genetic Engineering: Principles and Methods; Setlow, J.K., Ed.; Springer: Boston, MA, USA, 1997; pp. 257–269. [Google Scholar]
- Williams, P.A.; Jones, R.M.; Shaw, L.E. A third transposable element, ISPpu12, from the toluene-xylene catabolic plasmid pWW0 of Pseudomonas putida mt-2. J. Bacteriol. 2002, 184, 6572–6580. [Google Scholar] [CrossRef]
- Throne-Holst, M.; Wentzel, A.; Ellingsen, T.E.; Kotlar, H.K.; Zotchev, S.B. Identification of novel genes involved in long-chain n-alkane degradation by Acinetobacter sp. strain DSM 17874. Appl. Environ. Microbiol. 2007, 73, 3327–3332. [Google Scholar] [CrossRef]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 107. [Google Scholar] [CrossRef]
- Wu, P.; Wang, Y.S.; Sun, F.L.; Wu, M.L.; Peng, Y.L. Bacterial polycyclic aromatic hydrocarbon ring-hydroxylating dioxygenases in the sediments from the Pearl River estuary, China. Appl. Microbiol. Biotechnol. 2014, 98, 875–884. [Google Scholar] [CrossRef]
- Kim, E.; Aversano, P.J.; Romine, M.F.; Schneider, R.P.; Zylstra, G.J. Homology between genes for aromatic hydrocarbon degradation in surface and deep-subsurface Sphingomonas strains. Appl. Environ. Microbiol. 1996, 62, 1467–1470. [Google Scholar] [Green Version]
- Velazquez, F.; De Lorenzo, V.; Valls, M. The m-xylene biodegradation capacity of Pseudomonas putida mt-2 is submitted to adaptation to abiotic stresses: evidence from expression profiling of xyl genes. Environ. Microbiol. 2006, 8, 591–602. [Google Scholar] [CrossRef]
- Zylstra, G.J.; Gibson, D.T. Aromatic Hydrocarbon Degradation: A Molecular Approach. In Genetic Engineering: Principles and Methods; Setlow, J.K., Ed.; Springer: Boston, MA, USA, 1991; pp. 183–203. [Google Scholar]
- Busch, A.; Lacal, J.; Silva-Jímenez, H.; Krell, T.; Ramos, J.L. Catabolite Repression of the TodS/TodT Two-Component System and Effector-Dependent Transphosphorylation of TodT as the Basis for Toluene Dioxygenase Catabolic Pathway Control▿. J. Bacteriol. 2010, 192, 4246–4250. [Google Scholar] [CrossRef]
- Tao, Y.; Bentley, W.E.; Wood, T.K. Regiospecific oxidation of naphthalene and fluorene by toluene monooxygenases and engineered toluene 4-monooxygenases ofPseudomonas mendocina KR1. Biotechnol. Bioeng. 2005, 90, 85–94. [Google Scholar] [CrossRef]
- Kukor, J.J.; Olsen, R.H. Genetic organization and regulation of a meta cleavage pathway for catechols produced from catabolism of toluene, benzene, phenol, and cresols by Pseudomonas pickettii PKO1. J. Bacteriol. 1991, 173, 4587–4594. [Google Scholar] [CrossRef] [Green Version]
- Parales, R.E.; Parales, J.V.; Pelletier, D.A.; Ditty, J.L. Chapter 1 Diversity of Microbial Toluene Degradation Pathways. In Advances in Applied Microbiology; Allen, L., Ed.; Academic Press: Cambridge, MA, USA, 2008; Volume 64, pp. 1–73. [Google Scholar]
- Muangchinda, C.; Chavanich, S.; Viyakarn, V.; Watanabe, K.; Imura, S.; Vangnai, A.S.; Pinyakong, O. Abundance and diversity of functional genes involved in the degradation of aromatic hydrocarbons in Antarctic soils and sediments around Syowa Station. Environ. Sci. Pollut. Res. 2015, 22, 4725–4735. [Google Scholar] [CrossRef] [PubMed]
- van der Meer, J.R. Evolution of novel metabolic pathways for the degradation of chloroaromatic compounds. Antonie van Leeuwenhoek 1997, 71, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, H.; Koyama, Y.; Miyakoshi, M.; Miyazaki, R.; Yano, H.; Sota, M.; Ohtsubo, Y.; Tsuda, M.; Miyazaki, K. Novel organization of aromatic degradation pathway genes in a microbial community as revealed by metagenomic analysis. ISME J. 2009, 3, 1335–1348. [Google Scholar] [CrossRef] [PubMed]
- Stuardo, M.; Vásquez, M.; Vicuña, R.; González, B. Molecular approach for analysis of model fungal genes encoding ligninolytic peroxidases in wood-decaying soil systems. Lett. Appl. Microbiol. 2004, 38, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashtban, M.; Schraft, H.; Syed, T.A.; Qin, W. Fungal biodegradation and enzymatic modification of lignin. Int. J. Biochem. Mol. Boil. 2010, 1, 36–50. [Google Scholar]
- Jansa, J.; Treseder, K.K. Chapter 19: Introduction: Mycorrhizas and the carbon cycle. In Mycorrhizal Mediation of Soil: Fertility, Structure, and Carbon Storage; Johnson, N.C., Gehring, C., Jansa, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 1, pp. 343–351. [Google Scholar]
- Ruiz-Dueñas, F.J.; Lundell, T.; Floudas, D.; Nagy, L.G.; Barrasa, J.M.; Hibbett, D.S.; Martinez, A.T.; Mgbeahuruike, A.C.; Kovalchuk, A.; Asiegbu, F.O. Lignin-degrading peroxidases in Polyporales: an evolutionary survey based on 10 sequenced genomes. Mycologia 2013, 105, 1428–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Zhang, P.; Qin, Y.; Tu, Q.C.; Yang, Y.F.; He, Z.; Schadt, C.W.; Zhou, J.Z. Network succession reveals the importance of competition in response to emulsified vegetable oil amendment for uranium bioremediation. Env. Microbiol. 2016, 18, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Nagarajan, K.; Loh, K.-C. Biodegradation of aromatic compounds: current status and opportunities for biomolecular approaches. Appl. Microbiol. Biotechnol. 2009, 85, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef] [PubMed]
- Rojo, F. Degradation of alkanes by bacteria. Environ. Microbiol. 2009, 11, 2477–2490. [Google Scholar] [CrossRef]
- Moreno, R.; Rojo, F. Enzymes for Aerobic Degradation of Alkanes in Bacteria. In Aerobic Utilization of Hydrocarbons, Oils, and Lipids; Rojo, F., Ed.; Springer Science and Business Media LLC: Berlin, Germany, 2017; pp. 1–25. [Google Scholar]
- Seo, J.-S.; Keum, Y.-S.; Li, Q.X. Bacterial Degradation of Aromatic Compounds. Int. J. Environ. Res. Public Health 2009, 6, 278–309. [Google Scholar] [CrossRef] [PubMed]
- Carmona, M.; Zamarro, M.T.; Blázquez, B.; Durante-Rodríguez, G.; Juárez, J.F.; Valderrama, J.A.; Barragán, M.J.L.; García, J.L.; Diaz, E. Anaerobic Catabolism of Aromatic Compounds: a Genetic and Genomic View. Microbiol. Mol. Biol. Rev. 2009, 73, 71–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, G.; Boll, M.; Heider, J. Microbial degradation of aromatic compounds — from one strategy to four. Nat. Rev. Genet. 2011, 9, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Ghattas, A.-K.; Fischer, F.; Wick, A.; Ternes, T.A. Anaerobic biodegradation of (emerging) organic contaminants in the aquatic environment. Water Res. 2017, 116, 268–295. [Google Scholar] [CrossRef] [PubMed]
- Abbasian, F.; Lockington, R.; Megharaj, M.A.; Naidu, R. Review on the Genetics of Aliphatic and Aromatic Hydrocarbon Degradation. Appl. Biochem. Biotechnol. 2016, 178, 224–250. [Google Scholar] [CrossRef] [PubMed]
- Musat, F. The anaerobic degradation of gaseous, nonmethane alkanes — From in situ processes to microorganisms. Comput. Struct. Biotechnol. J. 2015, 13, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.-Y.; Mbadinga, S.M.; Liu, Y.-F.; Yang, S.-Z.; Liu, J.-F.; Ye, R.-Q.; Gu, J.-D.; Mu, B.-Z. Insights into the Anaerobic Biodegradation Pathway of n-Alkanes in Oil Reservoirs by Detection of Signature Metabolites. Sci. Rep. 2015, 5, 9801. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.; Marsh, E.N.G. Mechanism of Benzylsuccinate Synthase: Stereochemistry of Toluene Addition to Fumarate and Maleate. J. Am. Chem. Soc. 2005, 127, 8608–8609. [Google Scholar] [CrossRef] [PubMed]
- Safinowski, M.; Meckenstock, R.U. Methylation is the initial reaction in anaerobic naphthalene degradation by a sulfate-reducing enrichment culture. Environ. Microbiol. 2006, 8, 347–352. [Google Scholar] [CrossRef]
- Rabus, R.; Boll, M.; Heider, J.; Meckenstock, R.U.; Buckel, W.; Einsle, O.; Ermler, U.; Golding, B.T.; Gunsalus, R.P.; Kroneck, P.M.; et al. Anaerobic Microbial Degradation of Hydrocarbons: From Enzymatic Reactions to the Environment. J. Mol. Microbiol. Biotechnol. 2016, 26, 5–28. [Google Scholar] [CrossRef] [Green Version]
- Widdel, F.; Musat, F. Diversity and Common Principles in Enzymatic Activation of Hydrocarbons. In Handbook of Hydrocarbon and Lipid Microbiology; Kenneth, N.T., Ed.; Springer Science and Business Media LLC: Berlin, Germany, 2010; pp. 981–1009. [Google Scholar]
- Grossi, V.; Cravo-Laureau, C.; Rontani, J.-F.; Cros, M.; Hirschler-Réa, A. Anaerobic oxidation of n-alkenes by sulphate-reducing bacteria from the genus Desulfatiferula: n-Ketones as potential metabolites. Res. Microbiol. 2011, 162, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Rontani, J.-F.; Mouzdahir, A.; Michotey, V.; Bonin, P. Aerobic and anaerobic metabolism of squalene by a denitrifying bacterium isolated from marine sediment. Arch. Microbiol. 2002, 178, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Ettwig, K.F.; Shima, S.; Kahnt, J.; Medema, M.H.; Camp, H.J.M.O.D.; Jetten, M.S.M.; Strous, M.; Van De Pas-Schoonen, K.T.; Van De Pas-Schoonen, K.T. Denitrifying bacteria anaerobically oxidize methane in the absence ofArchaea. Environ. Microbiol. 2008, 10, 3164–3173. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Ma, A.; Qi, H.; Zhuang, X.; Zhuang, G. Anaerobic oxidation of methane: An “active” microbial process. MicrobiologyOpen 2015, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Reynolds, D.; Liu, M.; Stark, M.; Kjelleberg, S.; Webster, N.S.; Thomas, T. Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts. Proc. Natl. Acad. Sci. USA 2012, 109, E1878–E1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wintermute, E.H.; Silver, P.A. Dynamics in the mixed microbial concourse. Genes Dev. 2010, 24, 2603–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seth, E.C.; Taga, M.E. Nutrient cross-feeding in the microbial world. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.A.; Mahmood, Q.; Shah, M.M.; Pervez, A.; Asad, S.A. Microbial Ecology of Anaerobic Digesters: The Key Players of Anaerobiosis. Sci. World J. 2014, 2014, 1–21. [Google Scholar]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar]
- Plugge, C.M.; Zhang, W.; Scholten, J.C.M.; Stams, A.J.M. Metabolic Flexibility of Sulfate-Reducing Bacteria. Front. Microbiol. 2011, 2, 81. [Google Scholar] [CrossRef] [Green Version]
- Mbadinga, S.M.; Wang, L.-Y.; Zhou, L.; Liu, J.-F.; Gu, J.-D.; Mu, B.-Z. Microbial communities involved in anaerobic degradation of alkanes. Int. Biodeterior. Biodegrad. 2011, 65, 1–13. [Google Scholar] [CrossRef]
- Aitken, C.; Jones, D.; Maguire, M.; Gray, N.; Sherry, A.; Bowler, B.; Ditchfield, A.; Larter, S.; Head, I.; Larter, S. Evidence that crude oil alkane activation proceeds by different mechanisms under sulfate-reducing and methanogenic conditions. Geochim. Et Cosmochim. Acta 2013, 109, 162–174. [Google Scholar] [CrossRef]
- Musat, F.; Wilkes, H.; Behrends, A.; Woebken, D.; Widdel, F. Microbial nitrate-dependent cyclohexane degradation coupled with anaerobic ammonium oxidation. ISME J. 2010, 4, 1290–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinsteuber, S.; Schleinitz, K.M.; Vogt, C. Key players and team play: anaerobic microbial communities in hydrocarbon-contaminated aquifers. Appl. Microbiol. Biotechnol. 2012, 94, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, A.C.; Beller, H.R.; Edwards, E.A. Metabolites detected during biodegradation of 13C6-benzene in nitrate-reducing and methanogenic enrichment cultures. Environ. Sci. Technol. 2005, 39, 6681–6691. [Google Scholar] [CrossRef] [PubMed]
- Coates, J.D.; Chakraborty, R.; McInerney, M.J. Anaerobic benzene biodegradation—a new era. Res. Microbiol. 2002, 153, 621–628. [Google Scholar] [CrossRef]
- Gieg, L.M.; Fowler, S.J.; Berdugo-Clavijo, C. Syntrophic biodegradation of hydrocarbon contaminants. Curr. Opin. Biotechnol. 2014, 27, 21–29. [Google Scholar] [CrossRef]
- Gray, N.D.; Sherry, A.; Grant, R.J.; Rowan, A.K.; Hubert, C.R.J.; Callbeck, C.M.; Aitken, C.M.; Jones, D.M.; Adams, J.J.; Larter, S.R.; et al. The quantitative significance of Syntrophaceae and syntrophic partnerships in methanogenic degradation of crude oil alkanes. Environ. Microbiol. 2011, 13, 2957–2975. [Google Scholar] [CrossRef]
- Gray, N.D.; Sherry, A.; Hubert, C.; Dolfing, J.; Head, I.M. Chapter 5 - Methanogenic Degradation of Petroleum Hydrocarbons in Subsurface Environments: Remediation, Heavy Oil Formation, and Energy Recovery. In Advances in Applied Microbiology; Laskin, A.I., Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2010; Volume 72, pp. 137–161. [Google Scholar]
- Aburto, A.; Fahy, A.; Coulon, F.; Lethbridge, G.; Timmis, K.N.; Ball, A.S.; McGenity, T.J. Mixed aerobic and anaerobic microbial communities in benzene-contaminated groundwater. J. Appl. Microbiol. 2009, 106, 317–328. [Google Scholar] [CrossRef]
- Aburto-Medina, A.; Ball, A.S. Microorganisms involved in anaerobic benzene degradation. Ann. Microbiol. 2015, 65, 1201–1213. [Google Scholar] [CrossRef]
- Lade, H.; Paul, D.; Kweon, J.H. N-Acyl Homoserine Lactone-Mediated Quorum Sensing with Special Reference to Use of Quorum Quenching Bacteria in Membrane Biofouling Control. BioMed Res. Int. 2014, 2014, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum Sensing in Bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voloshin, S.A.; Kaprelyants, A.S. Cell-cell interactions in bacterial populations. Biochemistry (Moscow) 2004, 69, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, E.; Fodelianakis, S.; Korkakaki, E.; Kalogerakis, N. Biosurfactant production from marine hydrocarbon-degrading consortia and pure bacterial strains using crude oil as carbon source. Front. Microbiol. 2015, 6, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Amrani, A.; Dumas, A.-S.; Wick, L.Y.; Yergeau, E.; Berthomé, R. “Omics” Insights into PAH Degradation toward Improved Green Remediation Biotechnologies. Environ. Sci. Technol. 2015, 49, 11281–11291. [Google Scholar] [CrossRef] [PubMed]
- Galán, J.E.; Lara-Tejero, M.; Marlovits, T.C.; Wagner, S. Bacterial Type III Secretion Systems: Specialized Nanomachines for Protein Delivery into Target Cells. Annu. Rev. Microbiol. 2014, 68, 415–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfante, P.; Anca, I.-A. Plants, Mycorrhizal Fungi, and Bacteria: A Network of Interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Genes | Function | Example Organism of Origin | Gene Location (Chromosome or Plasmid) | Reference |
|---|---|---|---|---|
| Nah genes (G, T, H, I, N, L, O, M, K, J, Y, W) | Naphthalene catabolic genes (nah Y is chemotaxis gene and nah W may aid in adapting to extreme conditions) | Pseudomonas putida G7 | Chromosome and/or plasmid | [57] |
| Ndo genes | Naphthalene dioxygenases | Pseudomonas putida | Chromosome and/or plasmid | [58,59] |
| Dox genes (bphA bphE bphF bphG) | Dibenzothiophene oxidation (meta cleavage pathway) | Pseudomonas. sp.C18 | Plasmid | [58,59,60] |
| Pah | Naphthalene-Phenanthrene dioxygenase genes | Pseudomonas putida OUS82, Pseudomonas aeruginosa Pak1 | Plasmid | [58] |
| Nag gene | Naphthalene dioxygenase genes | Ralstonia sp. strain U2 | Plasmid | [61] |
| Phn genes (C, I, H) | Phenanthrene degradation genes | Alcaligenes faecalis AFK2 strain | Plasmid | [62] |
| Fln genes (RB, ED1) | Fluorine degradation | Terrabacter sp. strain DBF63 | Plasmid | [63] |
| Pht genes (Aa, Ab, B, Ac, Ad) | Degrading polycyclic aromatic hydrocarbons (PAHs) to ring cleavage metabolites (phthalate) | Mycobacterium vanbaalenii PYR-1 | Plasmid | [64] |
| CAT genes (A, B, C, R) | Catechol catabolic genes, cat central ortho pathway | Burkholderia strain LB400 | Chromosome | [65,66] |
| Pca genes (ligA, fldV, pmdA, proOa, (ligB, fldU, pmdB, pmdC, proD, pcmB, | Catabolism of the phenolic compounds (protocatechuate) | Agrobacterium tumefaciens | Chromosome | [67] |
| Gdo genes | Lignin and salicylate degradation (bacterial and fungal), gentisate 1,2-dioxygenase, cleavage of the gentisate aromatic ring | Rhodovulum sp. Strain NI22, Pseudomonas alcaligenes NCIMB 9867 (strain P25X) | Chromosome and/or plasmid | [68,69,70] |
| Alk genes (B, F, G, H, J, K, L) | Hydroxylation of aliphatic hydrocarbons | Pseudomonas oleovorans | Chromosome and/or plasmid | [71,72] |
| Ben genes (A, B, C, D) | Benzoate catabolic | Halomonas organivorans | Chromosome | [71] |
| CYP genes (pb-1, pb-2 and pb-3, CYP153) | Fungal and Bacterial Cytochrome P450 monooxgenase | Basidiomycetes, Acinetobacter calcoaceticus EB104 | Chromosome | [73] |
| LadA | Long-chain alkane mooxygenase-Degradation of long-chained alkanes | Geobacillus thermodenitrificans NG80-2 | Plasmid | [71] |
| Alm (A) | A flavin-binding mooxygenase-Degradation of long-chained alkanes | Acinetobacter sp. DSM 17874 | Chromosome | [71] |
| Phd genes (E, F, G, H, I, J, K) | Aromatic hydrocarbon degradation | Comamonas testosteroni GZ39 | Chromosome | [62,74] |
| Nid genes (A, A3, B2, B3, D,) | Pyrene degradation | Mycobacterium sp. PYR-1 | Chromosome | [58] |
| TOL genes (xyl) | Toluene degradation | Pseudomonas putida mt-2 | Plasmid | [75] |
| Alm | N-Alkanes (C32 and beyond) | Acinetobacter sp. DSM 17874 | Chromosome | [76] |
| Phn genes (B, C, R, S) | PAH degradation | Burkholderia sp. RP007 | Plasmid | [57] |
| Nar genes (Aa, Ab, B) | Naphthalene degradation | Rhodococcus sp. NCIMB12038 | Plasmid | [77] |
| Nid (A) | High-molecular-weight pahs degradation (e.g., pyrene and Fluoranthene) | Mycobacterium sp. PYR-1, | Chromosome | [78] |
| Dbf genes (A1, A2) | Terminal oxygenase genes of angular dioxygenase (Fluorine degradation) | Terrabacter sp. strain DBF63 | Plasmid | [63] |
| Xyl genes (X, Y, Z, L) | M-xylene degradation | Pseudomonas putida mt2 | Plasmid | [79,80] |
| Tod genes (A, B, C1, C2, D, E, C1C2BA, C1C2BAD, CIC2BADE) | Toluene dioxygenase (tod) metabolism of toluene, benzene, and ethylbenzene. | Pseudomonas putida | Chromosome and/or plasmid | [81,82] |
| tom | Toluene ortho-monooxygenase, oxidation of the polycyclic Aromatic hydrocarbons naphthalene and fluorene | Pseudomonas mendocina KR1 | Plasmid | [83] |
| Tbu genes (E, F, G, K, I, H, J) | BTEX, meta cleavage | Pseudomonas aeruginosa PAO1, R. | Chromosome | [84] |
| tmo | Multicomponent monooxygenase Enzyme complexes involved in aerobic benzene, toluene, ethylbenzene and xylene (BTEX) degradation | Ralstonia pickettii PKO1 | Chromosome | [85] |
| Xyl (E) | Catechol 2,3-dioxygenase (ring cleavage reaction in PAH degradation) | Sphingomonas yanoikuyae | Chromosome | [86] |
| Dmp genes (K, L, M, N, O, P, Q, B, C, D, E, H, F, G, I) | Involved in the meta cleavage pathway | Pseudomonas sp. CF600 | Plasmid | [87,88] |
| LiP genes (A, D, LipJ | Fungal Lignin peroxidase | Phanerochaete chrysosporium | Chromosome | [89,90] |
| MnP | Fungal Manganese Peroxidase | Phanerochaete chrysosporium | Chromosome | [89] |
| POD genes | Fungal Lignin degradation (heme – peroxidase encoding) | Bjerkandera adusta, Ganoderma sp. | Chromosome | [91,92] |
| HTP | Fungal Lignin degradation heme-thiolate peroxidase (heme – peroxidase encoding) | Rhodonia placenta | Chromosome | [92] |
| DyP | Fungal Lignin degradation Dye-decolorizing peroxidase (heme – peroxidase encoding) | Thanatephorus cucumeris | Chromosome | [92] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truskewycz, A.; Gundry, T.D.; Khudur, L.S.; Kolobaric, A.; Taha, M.; Aburto-Medina, A.; Ball, A.S.; Shahsavari, E. Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems—Fate and Microbial Responses. Molecules 2019, 24, 3400. https://doi.org/10.3390/molecules24183400
Truskewycz A, Gundry TD, Khudur LS, Kolobaric A, Taha M, Aburto-Medina A, Ball AS, Shahsavari E. Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems—Fate and Microbial Responses. Molecules. 2019; 24(18):3400. https://doi.org/10.3390/molecules24183400
Chicago/Turabian StyleTruskewycz, Adam, Taylor D. Gundry, Leadin S. Khudur, Adam Kolobaric, Mohamed Taha, Arturo Aburto-Medina, Andrew S. Ball, and Esmaeil Shahsavari. 2019. "Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems—Fate and Microbial Responses" Molecules 24, no. 18: 3400. https://doi.org/10.3390/molecules24183400
APA StyleTruskewycz, A., Gundry, T. D., Khudur, L. S., Kolobaric, A., Taha, M., Aburto-Medina, A., Ball, A. S., & Shahsavari, E. (2019). Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems—Fate and Microbial Responses. Molecules, 24(18), 3400. https://doi.org/10.3390/molecules24183400