Integrative Analysis of Terpenoid Profiles and Hormones from Fruits of Red-Flesh Citrus Mutants and Their Wild Types


Abstract
:1. Introduction
2. Results
2.1. Carotenoids
2.2. Limonoid Aglycones
2.3. Volatiles
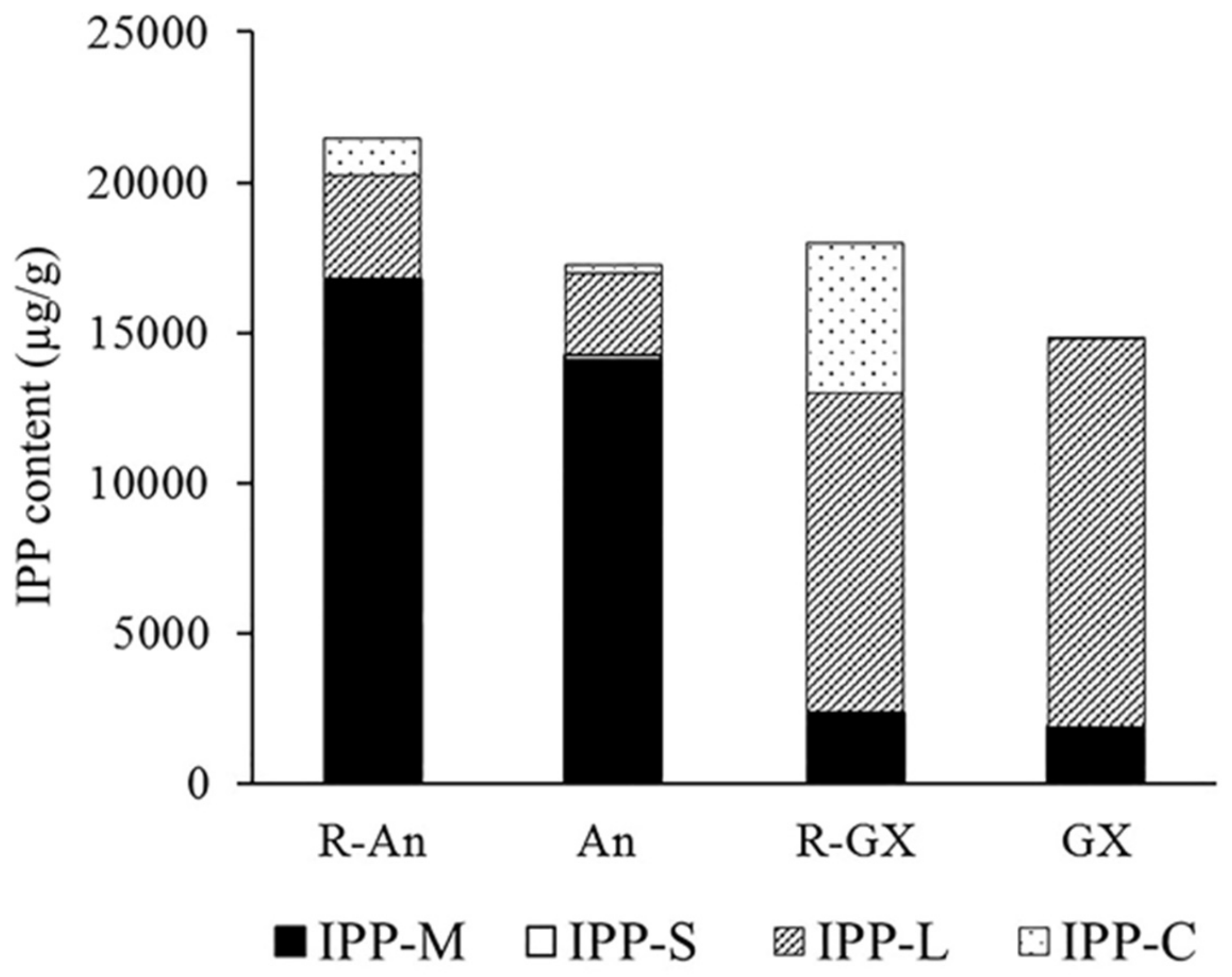
2.4. Isopentenyl Diphosphate (IPP) Content Analysis
2.5. Phytohormones in Citrus Fruits
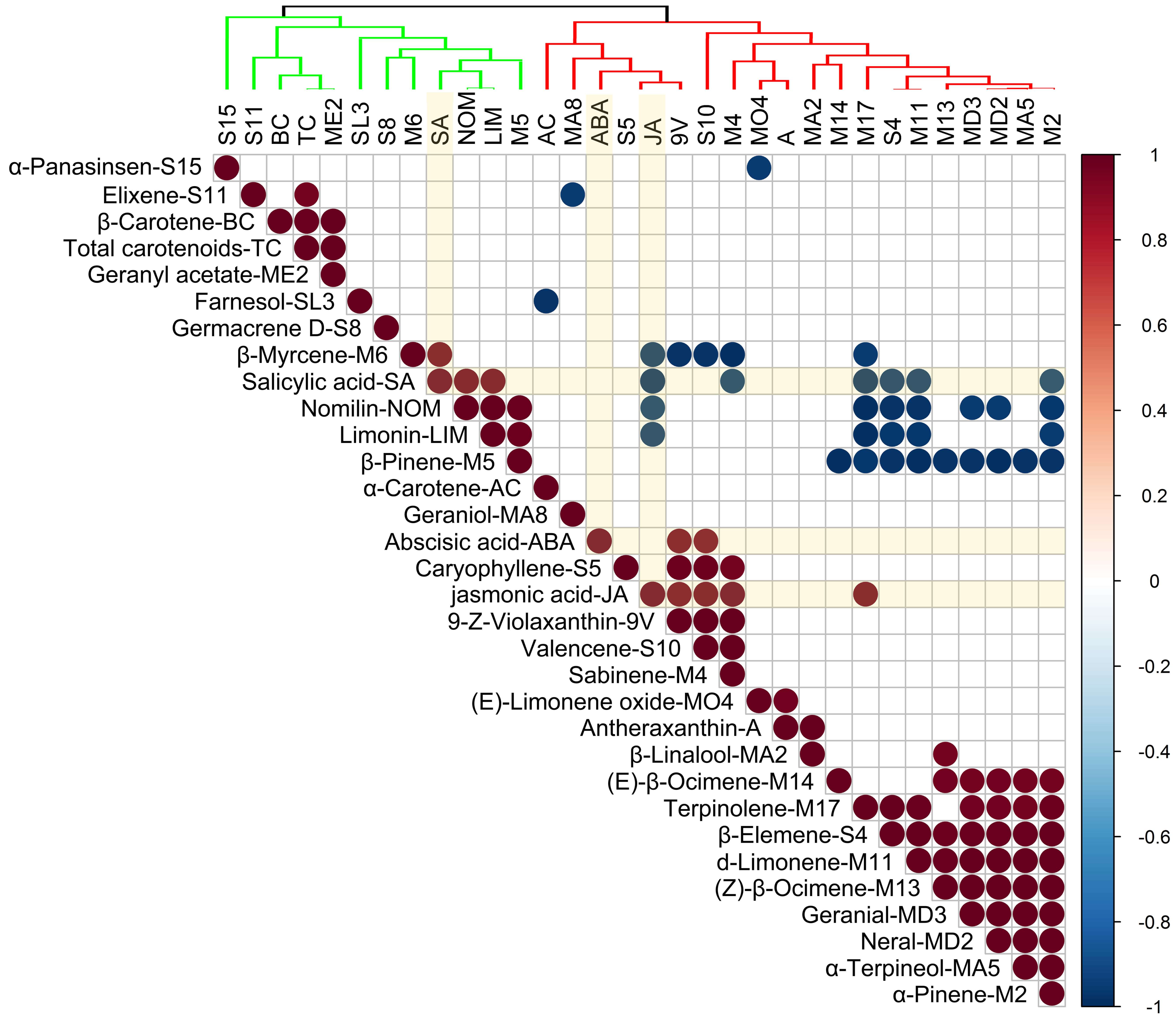
2.6. The Correlation Between Plant Hormones and Terpenoids
2.7. Principal Component Analysis Based on Carotenoids and Volatiles
3. Discussion
3.1. Equivalent Quantities of IPP in the Terpenoid Biosynthetic Pathway Are Prone to Stability
3.2. Effects of Hormones on Terpenoid Metabolism
3.3. Effects of Red-Flesh Bud Mutations on Metabolite Profiles
4. Materials and Methods
4.1. Plant Materials
4.2. Standards and Reagents
4.3. Carotenoid Extraction with MTBE and HPLC Analysis
4.4. Limonoid Aglycone Extraction and HPLC Analysis
4.5. Plant Hormone Extraction and Analysis with LC-MS
4.6. Volatile Compound Extraction and GC-MS Analysis
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Butelli, E.; Garcia-Lor, A.; Licciardello, C.; Las Casas, G.; Hill, L.; Recupero, G.R.; Keremane, M.L.; Ramadugu, C.; Krueger, R.; Xu, Q.; et al. Changes in anthocyanin production during domestication of Citrus. Plant Physiol. 2017, 173, 2225–2242. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, R. History. world distribution, botany and varieties. In The Citrus Industry; Reuther, W., Webber, H., Batchelor, L., Eds.; University of California Press: Berkeley, CA, USA, 1967; pp. 431–591. [Google Scholar]
- Goodwin, T.W. Metabolism, nutrition, and function of carotenoids. Annu. Rev. Nutr. 1986, 6, 273–297. [Google Scholar] [CrossRef] [PubMed]
- Monselise, S.; Halevy, A. Detection of lycopene in pink orange fruit. Science 1961, 133, 1478. [Google Scholar] [CrossRef] [PubMed]
- Saunt, J. Citrus Varieties of the World; Sinclair International Ltd.: Norwich, England, 2000; pp. 16–17. [Google Scholar]
- Liu, Q.; Xu, J.; Liu, Y.; Zhao, X.; Deng, X.; Guo, L.; Guo, J. A novel bud mutation that confers abnormal patterns of lycopene accumulation in sweet orange fruit (Citrus sinensis L. Osbeck). J. Exp. Bot. 2007, 58, 4161–4171. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yan, F.; Gao, H.; He, M.; Wang, Z.; Cheng, Y.; Deng, X.; Xu, J. Features of citrus terpenoid production as revealed by carotenoid, limonoid and aroma profiles of two pummelos (Citrus maxima) with different flesh color. J. Sci. Food Agric. 2015, 95, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ye, Q.; Jin, X.; Han, F.; Huang, X.; Cai, S.; Yang, L. A spontaneous bud mutant that causes lycopene and β-carotene accumulation in the juice sacs of the parental Guanxi pummelo fruits (Citrus grandis (L.) Osbeck). Sci. Hortic. 2016, 198, 379–384. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Pang, Y.; Cheng, Y.; Deng, X.; Xu, J. Comparative study of flavonoid production in lycopene-accumulated and blonde-flesh sweet oranges (Citrus sinensis) during fruit development. Food Chem. 2015, 184, 238–246. [Google Scholar] [CrossRef]
- Langenheim, J.H. Higher plant terpenoids: A phytocentric overview of their ecological roles. J. Chem. Ecol. 1994, 20, 1223–1280. [Google Scholar] [CrossRef]
- Cutler, A.J.; Krochko, J.E. Formation and breakdown of ABA. Trends Plant Sci. 1999, 4, 472–478. [Google Scholar] [CrossRef]
- Matusova, R.; Rani, K.; Verstappen, F.W.A.; Franssen, M.C.R.; Beale, M.H.; Bouwmeester, H.J. The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol. 2005, 139, 920–934. [Google Scholar] [CrossRef]
- Burg, S.P.; Burg, E.A. Role of ethylene in fruit ripening. Plant Physiol. 1962, 37, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Setha, S. Roles of abscisic acid in fruit ripening. Walailak J. Sci. Tech. 2012, 9, 297–308. [Google Scholar]
- Trainotti, L.; Tadiello, A.; Casadoro, G. The involvement of auxin in the ripening of climacteric fruits comes of age: The hormone plays a role of its own and has an intense interplay with ethylene in ripening peaches. J. Exp. Bot. 2007, 58, 3299–3308. [Google Scholar] [CrossRef] [PubMed]
- McAtee, P.; Karim, S.; Schaffer, R.; David, K. A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 2013, 4, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, K.; Zhang, S.; Ferguson, I. The role of salicylic acid in postharvest ripening of kiwifruit. Postharvest Biol. Tec. 2003, 28, 67–74. [Google Scholar] [CrossRef]
- Valero, D.; Díaz-Mula, H.M.; Zapata, P.J.; Castillo, S.; Guillén, F.; Martínez-Romero, D.; Serrano, M. Postharvest treatments with salicylic acid, acetylsalicylic acid or oxalic acid delayed ripening and enhanced bioactive compounds and antioxidant capacity in sweet cherry. J. Agric. Food Chem. 2011, 59, 5483–5489. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Zhang, C.; Pervaiz, T.; Zhao, P.; Liu, Z.; Wang, B.; Wang, C.; Zhang, L.; Fang, J.; Qian, J. Jasmonic acid involves in grape fruit ripening and resistant against Botrytis cinerea. Funct. Integr. Genom. 2016, 16, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, Y.; Zhang, L.; Ji, Y.; Tan, D.; Yuan, H.; Wang, A. The Jasmonate-activated transcription factor MdMYC2 regulates ETHYLENE RESPONSE FACTOR and ethylene biosynthetic genes to promote ethylene biosynthesis during apple fruit ripening. Plant Cell 2017, 29, 1316–1334. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cheng, Y.; Zhang, H.; Deng, X.; Chen, F.; Xu, J. Volatile constituents of wild citrus Mangshanyegan (Citrus nobilis Lauriro) peel oil. J. Agric. Food Chem. 2012, 60, 2617–2628. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, C.; He, M.; Li, J.; Cai, Y.; Ma, Y.; Xu, J. Largely different contents of terpenoids in beef red-flesh tangerine and its wild type. BMC Plant Biol. 2017, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Niedenfuhr, S.; ten Pierick, A.; van Dam, P.T.; Suarez-Mendez, C.A.; Noh, K.; Wahl, S.A. Natural isotope correction of MS/MS measurements for metabolomics and 13C fluxomics. Biotechnol. Bioeng. 2016, 113, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Toya, Y.; Banno, S.; Yoshikawa, K.; Matsuda, F.; Shimizu, H. 13C-metabolic flux analysis for mevalonate-producing strain of Escherichia coli. J. Biosci. Bioeng. 2017, 123, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, M.J. Characterization of Pinalate, a novel Citrus sinensis mutant with a fruit-specific alteration that results in yellow pigmentation and decreased ABA content. J. Exp. Bot. 2003, 54, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, S.; Devi, B.S.R.; Khorolragchaa, A.; Kim, Y.J.; Kim, J.H.; Jung, S.K.; Yang, D.C. Effect of salicylic acid and yeast extract on the accumulation of jasmonic acid and sesquiterpenoids in Panax ginseng adventitious roots. Russ. J. Plant Physl. 2014, 61, 811–817. [Google Scholar] [CrossRef]
- Huang, J.; Cardoza, Y.J.; Schmelz, E.A.; Raina, R.; Engelberth, J.; Tumlinson, J.H. Differential volatile emissions and salicylic acid levels from tobacco plants in response to different strains of Pseudomonas syringae. Planta 2003, 217, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Mosandl, A.; Wust, M. Induction of de novo volatile terpene biosynthesis via cytosolic and plastidial pathways by methyl jasmonate in foliage of Vitis vinifera L. J. Agric. Food Chem. 2005, 53, 2652–2657. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Chen, F.; Wang, X.; Rajapakse, N.C. Effect of methyl jasmonate on secondary metabolites of sweet basil (Ocimum basilicum L.). J. Agric. Food Chem. 2006, 54, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Van Schie, C.C.; Haring, M.A.; Schuurink, R.C. Tomato linalool synthase is induced in trichomes by jasmonic acid. Plant Mol. Biol. 2007, 64, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Talebi, M.; Moghaddam, M.; Pirbalouti, A.G. Methyl jasmonate effects on volatile oil compounds and antioxidant activity of leaf extract of two basil cultivars under salinity stress. Acta Physiol. Plant. 2018, 40, 34. [Google Scholar] [CrossRef]
- Zhang, X.N.; Liu, J.; Liu, Y.; Wang, Y.; Abozeid, A.; Yu, Z.G.; Tang, X. Metabolomics analysis reveals that ethylene and methyl jasmonate regulate different branch pathways to promote the accumulation of terpenoid indole alkaloids in Catharanthus roseus. J. Nat. Prod. 2018, 81, 335–342. [Google Scholar] [CrossRef]
- Patil, B.S.; Brodbelt, J.S.; Miller, E.G.; Turner, N.D. Potential health benefits of citrus: An overview. In Potential Health Benefits of Citrus; Patil, B.S., Turner, N.D., Miller, E.G., Brodbelt, J.S., Eds.; Oxford University Press: Washington, DC, USA, 2006; pp. 1–16. [Google Scholar]
- Wang, F.; Lin, J.; Xu, L.; Peng, Q.; Huang, H.; Tong, L.; Lu, Q.; Wang, C.; Yang, L. On higher nutritional and medical properties of a carotenoid-rich mutant pomelo (Citrus maxima (L.) Osbeck). Ind. Crop. Prod. 2019, 127, 142–147. [Google Scholar] [CrossRef]
- Li, S.J.; Wang, Z.; Ding, F.; Sun, D.; Ma, Z.C.; Cheng, Y.J.; Xu, J. Content changes of bitter compounds in ‘Guoqing No.1′ Satsuma mandarin (Citrus unshiu Marc.) during fruit development of consecutive 3 seasons. Food Chem. 2014, 145, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Ding, Y.; Chang, J.; Sun, X.; Zhang, L.; Wei, Q.; Cheng, Y.; Chen, L.; Xu, J.; Deng, X. Comprehensive insights on how 2,4-dichlorophenoxyacetic acid retards senescence in post-harvest citrus fruits using transcriptomic and proteomic approaches. J. Exp. Bot. 2014, 65, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.J.; Wu, H.; Lun, X.; Lu, D.W. Secretory cavity development and its relationship with the accumulation of essential oil in fruits of Citrus medica L. var. sarcodactylis (Noot.) Swingle. J. Integr. Plant Biol. 2006, 48, 573–583. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, D.; Cheng, Y.; Deng, X.; Chen, F.; Fang, L.; Ma, Z.; Xu, J. Chemotaxonomic Study of Citrus, Poncirus and Fortunella genotypes based on peel oil volatile compounds - deciphering the genetic origin of Mangshanyegan (Citrus nobilis Lauriro). PLoS ONE 2013, 8, e58411. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the materials are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Compounds | Code | R-An b | An | R-GX | GX |
|---|---|---|---|---|---|
| Violaxanthin | V | 4.92 ± 0.37 | 9.37 ± 1.16 * | / | / |
| 9-Z-Violaxanthin | 9V | 8.97 ± 0.85 | 18.98 ± 2.20 * | 0.95 ± 0.02 | 0.86 ± 0.01 * |
| Lutein | L | 0.82 ± 0.07 | 0.09 ± 0.07 * | / | / |
| α-Carotene | AC | 0.98 ± 0.02 | 0.92 ± 0.03 | 0.89 ± 0.02 | 0.48 ± 0.01 * |
| β-Carotene | BC | 2.21 ± 0.19 | 0.13 ± 0.01 * | 22.21 ± 0.19 | Trace * |
| Phytoene | P | / | / | 6.62 ± 0.04 | / * |
| Antheraxanthin | A | 5.22 ± 0.39 | 1.45 ± 0.30 * | 0.21 ± 0.01 | 0.42 ± 0.05 * |
| Zeaxanthin | Z | / | / | 1.06 ± 0.01 | 0.81 ± 0.01 * |
| β-Crytoxanthin | CR | 0.93 ± 0.02 | 4.39 ± 0.37 * | / | 0.83 ± 0.01 * |
| Lycopene | LY | 130.52 ± 6.80 | / * | 591.21 ± 9.65 | Trace * |
| γ-Carotene | CC | / | / | 30.56 ± 0.98 | / * |
| Total carotenoids | TC | 154.58 ± 7.82 | 35.34 ± 3.84 * | 622.55 ± 9.13 | 3.08 ± 0.34 * |
| Compounds | Code | R-An b | An | R-GX | GX |
|---|---|---|---|---|---|
| Limonin | LIM | 561.89 ± 46.41 | 449.16 ± 8.76 * | 1439.15 ± 153.45 | 1722.79 ± 200.16 |
| Nomilin | NOM | 13.20 ± 1.99 | 7.02 ± 0.29 * | 327.78 ± 36.16 | 426.01 ± 17.01 * |
| Total LAs | TLA | 575.09 ± 48.39 | 456.18 ± 9.03 * | 1766.93 ± 186.29 | 2148.80 ± 189.74 |
| Compounds b | Code | RI c | RIx d | R-An e | An | R-GX | GX |
|---|---|---|---|---|---|---|---|
| Monoterpenes | M | ||||||
| α-ThujeneT10 | M1 | 931 | 929 | 0.52 ± 0.06 | 0.84 ± 0.09 * | / | / |
| α-Pinene | M2 | 938 | 937 | 52.34 ± 0.18 | 40.52 ± 1.64 * | 4.14 ± 1.14 | 3.33 ± 0.74 |
| Camphene | M3 | 956 | 951 | 0.29 ± 0.02 | 0.26 ± 0.07 | / | / |
| Sabinene | M4 | 979 | 974 | 58.70 ± 7.11 | 101.00 ± 7.44 * | 3.09 ± 0.52 | 3.12 ± 0.21 |
| β-Pinene | M5 | 982 | 979 | 2.98 ± 0.33 | 4.56 ± 0.29 * | 9.56 ± 1.24 | 11.69 ± 0.81 |
| β-Myrcene | M6 | 994 | 991 | 158.44 ± 4.13 | 115.91 ± 5.68 * | 243.72 ± 82.07 | 226.04 ± 58.49 |
| PseudolimoneneT11 | M7 | 1008 | 1004 | 0.31 ± 0.05 | 0.13 ± 0.12 | / | / |
| α-Phellandrene | M8 | 1011 | 1005 | 13.10 ± 2.38 | 13.71 ± 1.40 | 0.14 ± 0.25 | / |
| α-Terpinene | M9 | 1023 | 1017 | 0.37 ± 0.07 | 0.44 ± 0.04 | / | / |
| SylvestreneT11 | M10 | 1027 | 1027 | 0.15 ± 0.13 | 0.24 ± 0.09 | / | / |
| d-Limonene | M11 | 1038 | 1030 | 7846.10 ± 436.16 | 6647.90 ± 347.94 * | 878.30 ± 282.27 | 656.70 ± 183.73 |
| β-PhellandreneT10 | M12 | 1040 | 1031 | 0.60 ± 0.25 | 1.48 ± 0.16 * | / | / |
| (Z)-β-Ocimene | M13 | 1044 | 1038 | 0.54 ± 0.04 | 0.43 ± 0.11 | 0.26 ± 0.01 | 0.24 ± 0.05 |
| (E)-β-Ocimene | M14 | 1055 | 1049 | 10.88 ± 1.07 | 9.20 ± 1.52 | 7.22 ± 0.18 | 5.51 ± 0.80 * |
| 4-CareneT11 | M15 | 1060 | 1009 | 0.33 ± 0.07 | / * | / | / |
| γ-Terpiene | M16 | 1066 | 1060 | 0.52 ± 0.09 | 0.62 ± 0.01 | / | / |
| Terpinolene | M17 | 1090 | 1088 | 3.50 ± 0.69 | 3.69 ± 0.33 | 0.23 ± 0.02 | 0.12 ± 0.11 |
| Monoterpene Alcohols | MA | ||||||
| (Z)-Sabinene hydrate | MA1 | 1079 | 1077 | 2.53 ± 0.25 | 3.28 ± 0.41 | / | / |
| β-Linalool | MA2 | 1107 | 1099 | 107.93 ± 4.81 | 38.65 ± 3.79 * | 2.67 ± 0.92 | 2.57 ± 0.15 |
| (E)-p-Mentha-2,8-dienolT12 | MA3 | 1133 | 1123 | 0.49 ± 0.05 | 0.24 ± 0.04 * | / | / |
| 4-Terpineol | MA4 | 1190 | 1177 | 0.65 ± 0.02 | 0.67 ± 0.10 | / | / |
| α-Terpineol | MA5 | 1206 | 1189 | 10.70 ± 0.65 | 7.65 ± 1.03 * | 1.15 ± 0.43 | 1.24 ± 0.25 |
| (E)-PiperitolT1 | MA6 | 1220 | 1208 | / | / | 0.14 ± 0.12 | 0.12 ± 0.11 |
| Citronellol | MA7 | 1236 | 1228 | 1.55 ± 0.13 | 1.66 ± 0.25 | / | / |
| Geraniol | MA8 | 1261 | 1255 | 2.03 ± 0.26 | 2.47 ± 0.54 | 1.47 ± 0.77 | 2.01 ± 0.25 |
| p-Mentha-1(7),8(10)-dien-9-olT1 | MA9 | 1303 | / | 1.68 ± 0.10 | 0.82 ± 0.15 * | / | / |
| Monoterpene alDehydes | MD | ||||||
| Citronellal | MD1 | 1164 | 1153 | 7.85 ± 0.38 | 5.27 ± 0.25 * | / | / |
| Neral | MD2 | 1251 | 1240 | 16.22 ± 0.58 | 12.01 ± 0.81 * | 2.28 ± 0.77 | 1.56 ± 0.54 |
| Geranial | MD3 | 1281 | 1270 | 22.28 ± 0.70 | 16.47 ± 1.60 * | 2.87 ± 1.02 | 2.22 ± 0.83 |
| PerillalT13 | MD4 | 1291 | 1272 | 6.44 ± 0.27 | 2.49 ± 0.20 * | / | / |
| Monoterpene Esters | ME | ||||||
| Neryl acetate | ME1 | 1366 | 1364 | 0.46 ± 0.06 | 0.30 ± 0.07 * | 0.26 ± 0.04 | / * |
| Geranyl acetate | ME2 | 1385 | 1382 | 0.32 ± 0.28 | 0.27 ± 0.14 | 0.57 ± 0.08 | 0.26 ± 0.06 * |
| Monoterpene Oxides | MO | ||||||
| (Z)-Linalool oxide | MO1 | 1078 | 1074 | / | / | 0.30 ± 0.07 | 0.09 ± 0.02 * |
| (E)-Linalool oxide | MO2 | 1093 | 1086 | / | / | 0.16 ± 0.14 | / |
| (Z)-Limonene oxide | MO3 | 1143 | 1134 | 0.46 ± 0.04 | 0.39 ± 0.07 | / | / |
| (E)-Limonene oxide | MO4 | 1148 | 1138 | 3.24 ± 0.09 | 0.21 ± 0.05 * | 0.29 ± 0.07 | 0.27 ± 0.09 |
| Sesquiterpenes | S | ||||||
| δ-Elemene T13 | S1 | 1338 | 1338 | / | / | 1.95 ± 0.58 | 1.64 ± 0.14 |
| Copaene T13 | S2 | 1380 | 1376 | 4.18 ± 0.37 | 2.26 ± 0.13 * | / | / |
| β-Cubebene T3 | S3 | 1391 | 1389 | 3.13 ± 0.18 | 1.59 ± 0.07 * | / | / |
| β-Elemene T13 | S4 | 1393 | 1391 | 1.17 ± 0.10 | 1.14 ± 0.05 | 0.93 ± 0.29 | 0.93 ± 0.17 |
| Caryophyllene | S5 | 1424 | 1419 | 2.88 ± 0.10 | 6.55 ± 0.09 * | 1.72 ± 0.22 | 0.75 ± 0.28 * |
| β-Farnesene | S6 | 1457 | 1457 | 0.84 ± 0.10 | 5.05 ± 0.29 * | / | / |
| α-Caryophyllene | S7 | 1461 | 1454 | 0.67 ± 0.03 | 0.50 ± 0.02 * | / | / |
| Germacrene D T2 | S8 | 1487 | 1481 | 2.85 ± 0.18 | 1.64 ± 0.08 * | 11.59 ± 1.98 | 7.11 ± 2.05 * |
| γ-Selinene T3 | S9 | 1489 | 1481 | 0.41 ± 0.05 | 0.79 ± 0.16 * | / | / |
| Valencene | S10 | 1497 | 1492 | 8.89 ± 0.11 | 16.19 ± 0.62 * | 2.34 ± 0.34 | 2.37 ± 0.52 |
| Elixene T13 | S11 | 1500 | 1471 | 1.38 ± 0.05 | 1.10 ± 0.40 | 1.70 ± 0.20 | 1.22 ± 0.20 * |
| α-Muurolene T3 | S12 | 1505 | 1499 | 0.62 ± 0.12 | 0.50 ± 0.09 | / | / |
| α-Farnesene T4 | S13 | 1508 | 1508 | 1.11 ± 0.16 | 12.86 ± 1.42 * | / | / |
| δ-Cadinene T13 | S14 | 1524 | 1524 | 4.91 ± 0.40 | 2.43 ± 0.20 * | / | / |
| α-Panasinsen T13 | S15 | 1525 | 1527 | 0.13 ± 0.22 | 0.69 ± 0.15 * | 0.50 ± 0.13 | 0.60 ± 0.18 |
| β-Sesquiphellandrene T4 | S16 | 1530 | 1524 | 0.27 ± 0.06 | 0.46 ± 0.12 | / | / |
| Germacrene B T13 | S17 | 1566 | 1557 | / | / | 0.45 ± 0.39 | 0.49 ± 0.16 |
| Sesquiterpene aLcohols | SL | ||||||
| (E)-Nerolidol | SL1 | 1567 | 1564 | 0.99 ± 0.12 | 0.73 ± 0.16 | / | / |
| Germacrene D-4-ol T3 | SL2 | 1585 | 1574 | 0.43 ± 0.03 | 0.51 ± 0.11 | / | / |
| Farnesol | SL3 | 1727 | 1713 | 1.02 ± 0.09 | 0.89 ± 0.23 | 0.73 ± 0.66 | 4.82 ± 0.40 * |
| Sesquiterpene alDehydes | SD | ||||||
| β-Sinensal T3 | SD1 | 1705 | 1695 | 1.31 ± 0.16 | 1.04 ± 0.43 | / | / |
| α-Sinensal T3 | SD2 | 1767 | 1752 | 1.39 ± 0.53 | 2.46 ± 0.71 | / | / |
| Sesquiterpene Ketone | SK | ||||||
| Nootkatone | SK1 | 1824 | 1808 | 0.69 ± 0.12 | 1.47 ± 0.56 | / | / |
| Sesquiterpene oxide | SO | ||||||
| Caryophyllene oxide | SO1 | 1591 | 1581 | 0.58 ± 0.11 | 1.86 ± 0.31 * | / | / |
| ALcohols | AL | ||||||
| (Z)-3-Hexenol | AL1 | 868 | 856 | 2.35 ± 0.34 | 2.36 ± 0.20 | 0.70 ± 0.09 | 0.73 ± 0.12 |
| (E)-2-Hexenol | AL2 | 878 | 862 | / | / | / | 0.49 ± 0.12 * |
| Hexanol | AL3 | 881 | 868 | 0.94 ± 0.31 | 1.04 ± 0.06 | 0.06 ± 0.10 | 0.35 ± 0.08 * |
| Octanol | AL4 | 1081 | 1071 | 3.65 ± 1.01 | 2.91 ± 0.64 | / | / |
| AlDehydes | AD | ||||||
| 3-Hexenal T7 | AD1 | 811 | 810 | 0.56 ± 0.40 | 0.67 ± 0.12 | / | / |
| Hexanal | AD2 | 812 | 800 | 2.06 ± 0.59 | 1.76 ± 0.31 | 0.10 ± 0.17 | 0.21 ± 0.19 |
| (E)-2-Hexenal | AD3 | 870 | 854 | 1.09 ± 0.14 | 0.71 ± 0.15 * | / | / |
| Nonanal | AD4 | 1116 | 1104 | 2.62 ± 0.57 | 1.62 ± 0.22 * | / | / |
| Decanal | AD5 | 1216 | 1206 | 12.84 ± 3.15 | 8.99 ± 1.73 | 0.33 ± 0.12 | 0.42 ± 0.05 |
| Undecanal | AD6 | 1318 | 1307 | 0.88 ± 0.12 | 0.42 ± 0.11 * | / | / |
| Dodecanal | AD7 | 1418 | 1409 | 2.34 ± 0.52 | 1.61 ± 0.15 | / | / |
| Monoterpenes | TM | 8149.68 | 6940.94 | 1146.67 | 906.74 | ||
| Monoterpene Alcohols | TMA | 127.57 | 55.45 | 5.43 | 5.93 | ||
| Monoterpene alDehydes | TMD | 52.78 | 36.23 | 5.14 | 3.78 | ||
| Monoterpene Oxides | TMO | 3.70 | 0.60 | 0.75 | 0.36 | ||
| Monoterpene Esters | TME | 0.78 | 0.57 | 0.83 | 0.26 | ||
| Sesquiterpenes | TS | 33.40 | 53.74 | 21.17 | 15.12 | ||
| Sesquiterpene aLcohols | TSL | 2.43 | 2.13 | 0.73 | 4.82 | ||
| Sesquiterpene alDehydes | TSD | 2.70 | 3.49 | / | / | ||
| Sesquiterpene Ketone | TSK | 0.69 | 1.47 | / | / | ||
| Sesquiterpene Oxide | TSO | 0.58 | 1.86 | / | / | ||
| Alcohols | TA | 6.94 | 6.31 | 0.76 | 1.57 | ||
| AlDehydes | TAD | 22.38 | 15.79 | 0.43 | 0.64 | ||
| MonoTerpenoids | TMT | 8334.51 | 7033.79 | 1158.82 | 917.08 | ||
| SesquiTerpenoids | TST | 39.80 | 62.70 | 21.90 | 19.93 | ||
| Non-Terpenoids | TNT | 29.33 | 22.10 | 1.19 | 2.21 | ||
| Total Volatiles | TV | 8403.64 | 7118.58 | 1181.91 | 939.22 |
| R-An b | An | R-GX | GX | |
|---|---|---|---|---|
| ABA | 1428.49 ± 103.87 | 2242.52 ± 126.82 * | 1089.35 ± 50.03 | 1276.14 ± 53.30 * |
| IAA | / | 17.35 ± 0.51 * | / | / |
| JA | 19.01 ± 4.32 | 23.02 ± 7.43 | 9.90 ± 0.38 | 9.04 ± 0.24 * |
| SA | 13.28 ± 2.09 | 12.24 ± 2.85 | 19.12 ± 3.01 | 19.38 ± 6.82 |
| Cultivar | Flesh Color | Code | Production Area | Remarks |
|---|---|---|---|---|
| Red Anliu orange | Red flesh | R-An | Citrus Research Institute of Guangxi, Guilin, Guangxi province | Red-flesh mutant of Anliu orange |
| Anliu orange | Yellow flesh | An | Common sweet orange | |
| Red-flesh Guanxi pummelo | Red flesh | R-GX | Fujian Academy of Agricultural Sciences, Xiamen, Fujian province | Red-flesh mutant of Guanxi pummelo |
| Guanxi pummelo | Pale yellow flesh | GX | Common white-flesh pummelo |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; He, M.; Wang, Z.; Xu, J. Integrative Analysis of Terpenoid Profiles and Hormones from Fruits of Red-Flesh Citrus Mutants and Their Wild Types. Molecules 2019, 24, 3456. https://doi.org/10.3390/molecules24193456
Liu C, He M, Wang Z, Xu J. Integrative Analysis of Terpenoid Profiles and Hormones from Fruits of Red-Flesh Citrus Mutants and Their Wild Types. Molecules. 2019; 24(19):3456. https://doi.org/10.3390/molecules24193456
Chicago/Turabian StyleLiu, Cuihua, Min He, Zhuang Wang, and Juan Xu. 2019. "Integrative Analysis of Terpenoid Profiles and Hormones from Fruits of Red-Flesh Citrus Mutants and Their Wild Types" Molecules 24, no. 19: 3456. https://doi.org/10.3390/molecules24193456
APA StyleLiu, C., He, M., Wang, Z., & Xu, J. (2019). Integrative Analysis of Terpenoid Profiles and Hormones from Fruits of Red-Flesh Citrus Mutants and Their Wild Types. Molecules, 24(19), 3456. https://doi.org/10.3390/molecules24193456