Peptides for Skin Protection and Healing in Amphibians


 ,
,  and
and 
Abstract
:1. Introduction
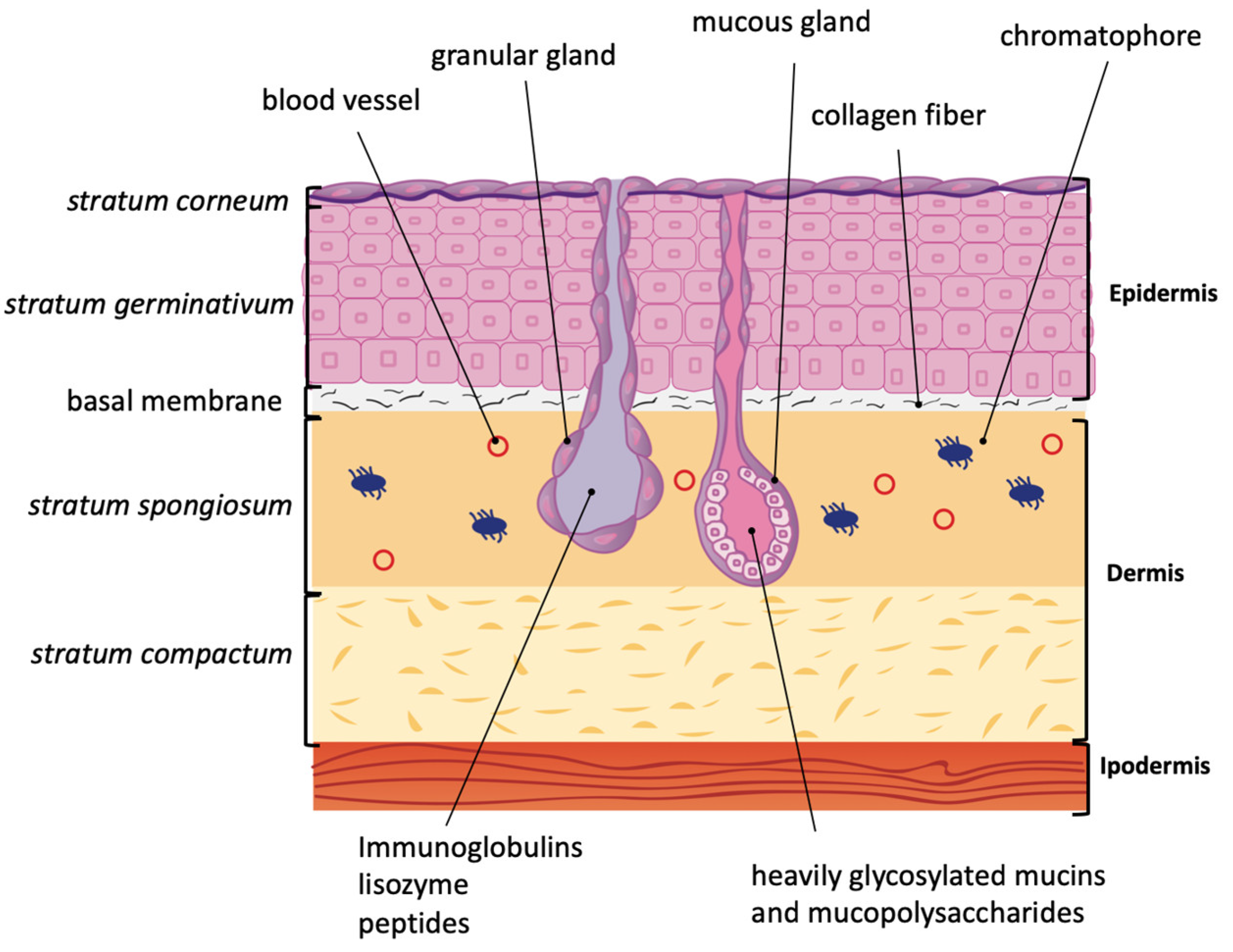
2. Amphibian Skin Glands
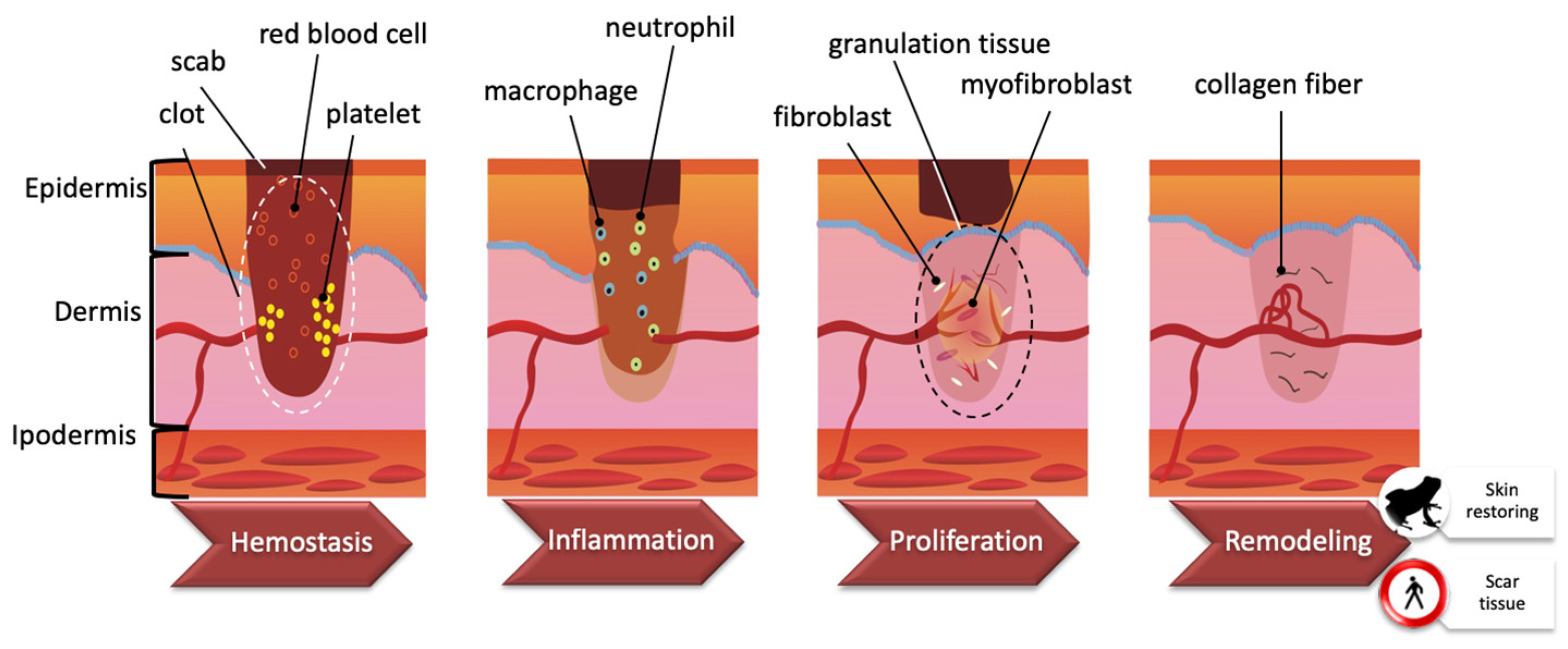
3. Wound Healing Process
4. Amphibian Peptides Involved in WH
5. Amphibian Peptides Known for Antioxidant/Free Radical Scavenging Activities
6. Amphibian Peptide Production for Ultraviolet Irradiation Adaptation
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duellman, W.E.; Trueb, L. Biology of Amphibians, 2nd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 1994; pp. 367–414. [Google Scholar]
- Felsemburgh, F.A.; de Almeida, P.G.; de Carvalho, E.S.S.P.; de Brito-Gitirana, L. Microscopical methods promote the understanding of the integument biology of Rhinella ornata. Micron 2009, 40, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Wells, K.D. The Ecology and Behaviour of Amphibians, 1st ed.; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Schumacher, U.; Adam, E.; Hauser, F.; Probst, J.C.; Hoffmann, W. Molecular anatomy of a skin gland: Histochemical and biochemical investigations on the mucous glands of Xenopus laevis. J. Histochem. Cytochem. 1994, 42, 57–65. [Google Scholar] [CrossRef]
- Ramsey, J.P.; Reinert, L.K.; Harper, L.K.; Woodhams, D.C.; Rollins-Smith, L.A. Immune defenses against Batrachochytrium dendrobatidis, a fungus linked to global amphibian declines, in the South African clawed frog, Xenopus laevis. Infect. Immun. 2010, 78, 3981–3992. [Google Scholar] [CrossRef] [PubMed]
- Rollins-Smith, L.A.; Conlon, J.M. Antimicrobial peptide defenses against chytridiomycosis, an emerging infectious disease of amphibian populations. Dev. Comp. Immunol. 2005, 29, 589–598. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A.; Ramsey, J.P.; Pask, J.D.; Reinert, L.K.; Woodhams, D.C. Amphibian immune defenses against chytridiomycosis: Impacts of changing environments. Integr. Comp. Biol. 2011, 51, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Gammill, W.M.; Fites, J.S.; Rollins-Smith, L.A. Norepinephrine depletion of antimicrobial peptides from the skin glands of Xenopus laevis. Dev. Comp. Immunol. 2012, 37, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.K.; Pasmans, F.; Dhaenens, M.; Deforce, D.; Bonte, D.; Verheyen, K.; Lens, L.; Martel, A. Skin mucosome activity as an indicator of Batrachochytrium salamandrivorans susceptibility in salamanders. PLoS ONE 2018, 13, e0199295. [Google Scholar] [CrossRef] [PubMed]
- Woodhams, D.C.; Brandt, H.; Baumgartner, S.; Kielgast, J.; Kupfer, E.; Tobler, U.; Davis, L.R.; Schmidt, B.R.; Bel, C.; Hodel, S.; et al. Interacting symbionts and immunity in the amphibian skin mucosome predict disease risk and probiotic effectiveness. PLoS ONE 2014, 9, e96375. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef]
- Bevins, C.L.; Zasloff, M. Peptides from frog skin. Annu. Rev. Biochem. 1990, 59, 395–414. [Google Scholar] [CrossRef] [PubMed]
- Laux-Biehlmann, A.; Mouheiche, J.; Veriepe, J.; Goumon, Y. Endogenous morphine and its metabolites in mammals: History, synthesis, localization and perspectives. Neuroscience 2013, 233, 95–117. [Google Scholar] [CrossRef] [PubMed]
- You, D.; Hong, J.; Rong, M.; Yu, H.; Liang, S.; Ma, Y.; Yang, H.; Wu, J.; Lin, D.; Lai, R. The first gene-encoded amphibian neurotoxin. J. Biol. Chem. 2009, 284, 22079–22086. [Google Scholar] [CrossRef] [PubMed]
- Martin, P. Wound healing--aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef] [PubMed]
- Kawasumi, A.; Sagawa, N.; Hayashi, S.; Yokoyama, H.; Tamura, K. Wound healing in mammals and amphibians: Toward limb regeneration in mammals. Curr. Top. Microbiol. Immunol. 2013, 367, 33–49. [Google Scholar] [PubMed]
- Murawala, P.; Tanaka, E.M.; Currie, J.D. Regeneration: The ultimate example of wound healing. Semin. Cell Dev. Biol. 2012, 23, 954–962. [Google Scholar] [CrossRef]
- Yoshizato, K. Molecular mechanism and evolutional significance of epithelial-mesenchymal interactions in the body- and tail-dependent metamorphic transformation of anuran larval skin. Int. Rev. Cytol. 2007, 260, 213–260. [Google Scholar]
- Campbell, L.J.; Crews, C.M. Wound epidermis formation and function in urodele amphibian limb regeneration. Cell. Mol. Life Sci. 2008, 65, 73–79. [Google Scholar] [CrossRef]
- Matsuda, H.; Yokoyama, H.; Endo, T.; Tamura, K.; Ide, H. An epidermal signal regulates Lmx-1 expression and dorsal-ventral pattern during Xenopus limb regeneration. Dev. Biol. 2001, 229, 351–362. [Google Scholar] [CrossRef]
- Satoh, A.; Graham, G.M.; Bryant, S.V.; Gardiner, D.M. Neurotrophic regulation of epidermal dedifferentiation during wound healing and limb regeneration in the axolotl (Ambystoma mexicanum). Dev. Biol. 2008, 319, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Levesque, M.; Villiard, E.; Roy, S. Skin wound healing in axolotls: A scarless process. J. Exp. Zool. B Mol. Dev. Evol. 2010, 314, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.W.; Monaghan, J.R.; Voss, S.R.; Maden, M. Skin regeneration in adult axolotls: A blueprint for scar-free healing in vertebrates. PLoS ONE 2012, 7, e32875. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Maruoka, T.; Aruga, A.; Amano, T.; Ohgo, S.; Shiroishi, T.; Tamura, K. Prx-1 expression in Xenopus laevis scarless skin-wound healing and its resemblance to epimorphic regeneration. J. Investig. Dermatol. 2011, 131, 2477–2485. [Google Scholar] [CrossRef] [PubMed]
- Spindel, E.R. Bombesin peptides. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 326–330. [Google Scholar]
- Anastasi, A.; Erspamer, V.; Bucci, M. Isolation and amino acid sequences of alytesin and bombesin, two analogous active tetradecapeptides from the skin of European discoglossid frogs. Arch. Biochem. Biophys. 1972, 148, 443–446. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Wan, S.G.; Wei, S.S.; Lee, W.H.; Zhang, Y. Bm-TFF2, a trefoil factor protein with platelet activation activity from frog Bombina maxima skin secretions. Biochem. Biophys. Res. Commun. 2005, 330, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, G.; Xiang, Y.; Wu, J.; Jiang, P.; Lee, W.; Zhang, Y. Bm-TFF2, a toad trefoil factor, promotes cell migration, survival and wound healing. Biochem. Biophys. Res. Commun. 2010, 398, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Ladram, A.; Besne, I.; Breton, L.; de Lacharriere, O.; Nicolas, P.; Amiche, M. Pharmacologic study of C-terminal fragments of frog skin calcitonin gene-related peptide. Peptides 2008, 29, 1150–1156. [Google Scholar] [CrossRef]
- Liu, H.; Mu, L.; Tang, J.; Shen, C.; Gao, C.; Rong, M.; Zhang, Z.; Liu, J.; Wu, X.; Yu, H.; et al. A potential wound healing-promoting peptide from frog skin. Int. J. Biochem. Cell Biol. 2014, 49, 32–41. [Google Scholar] [CrossRef]
- Di Grazia, A.; Luca, V.; Segev-Zarko, L.A.; Shai, Y.; Mangoni, M.L. Temporins A and B stimulate migration of HaCaT keratinocytes and kill intracellular Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 2520–2527. [Google Scholar] [CrossRef]
- Bian, W.; Meng, B.; Li, X.; Wang, S.; Cao, X.; Liu, N.; Yang, M.; Tang, J.; Wang, Y.; Yang, X. OA-GL21, a novel bioactive peptide from Odorrana andersonii, accelerated the healing of skin wounds. Biosci. Rep. 2018, 38, BSR20180215. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Y.; Zou, Z.; Yang, M.; Wu, C.; Su, Y.; Tang, J.; Yang, X. OM-LV20, a novel peptide from odorous frog skin, accelerates wound healing in vitro and in vivo. Chem. Biol. Drug Des. 2018, 91, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Duan, Z.; Tang, J.; Lv, Q.; Rong, M.; Lai, R. A short peptide from frog skin accelerates diabetic wound healing. FEBS J. 2014, 281, 4633–4643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Cheng, Y.; Yang, Y.; Liu, S.; Shi, H.; Lu, C.; Li, S.; Nie, L.; Su, D.; Deng, X.; et al. Polypeptides from the skin of Rana chensinensis exert the antioxidant and antiapoptotic activities on HaCaT cells. Anim. Biotechnol. 2017, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Longcore, J.E.; Pessier, A.P.; Nichols, D.K. Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to Amphibians. Mycologia 1999, 91, 219–227. [Google Scholar] [CrossRef]
- Berger, L.; Hyatt, A.D.; Speare, R.; Longcore, J.E. Life cycle stages of the amphibian chytrid Batrachochytrium dendrobatidis. Dis. Aquat. Organ. 2005, 68, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Briggs, C.J.; Daszak, P. The ecology and impact of chytridiomycosis: An emerging disease of amphibians. Trends Ecol. Evol. 2010, 25, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.; Marantelli, G.; Skerratt, L.F.; Speare, R. Virulence of the amphibian chytrid fungus Batrachochytium dendrobatidis varies with the strain. Dis. Aquat. Organ. 2005, 68, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Voyles, J.; Young, S.; Berger, L.; Campbell, C.; Voyles, W.F.; Dinudom, A.; Cook, D.; Webb, R.; Alford, R.A.; Skerratt, L.F.; et al. Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines. Science 2009, 326, 582–585. [Google Scholar] [CrossRef]
- Burkart, D.; Flechas, S.V.; Vredenburg, V.T.; Catenazzi, A. Cutaneous bacteria, but not peptides, are associated with chytridiomycosis resistance in Peruvian marsupial frogs. Anim. Cons. 2017, 20, 483–491. [Google Scholar] [CrossRef]
- Conlon, J.M.; Woodhams, D.C.; Raza, H.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; Rollins-Smith, L.A. Peptides with differential cytolytic activity from skin secretions of the lemur leaf frog Hylomantis lemur (Hylidae: Phyllomedusinae). Toxicon 2007, 50, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Holden, W.M.; Reinert, L.K.; Hanlon, S.M.; Parris, M.J.; Rollins-Smith, L.A. Development of antimicrobial peptide defenses of southern leopard frogs, Rana sphenocephala, against the pathogenic chytrid fungus, Batrachochytrium dendrobatidis. Dev. Comp. Immunol. 2015, 48, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Rollins-Smith, L.A.; Carey, C.; Longcore, J.; Doersam, J.K.; Boutte, A.; Bruzgal, J.E.; Conlon, J.M. Activity of antimicrobial skin peptides from ranid frogs against Batrachochytrium dendrobatidis, the chytrid fungus associated with global amphibian declines. Dev. Comp. Immunol. 2002, 26, 471–479. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A.; Reinert, L.K.; Miera, V.; Conlon, J.M. Antimicrobial peptide defenses of the Tarahumara frog, Rana tarahumarae. Biochem. Biophys. Res Commun. 2002, 297, 361–367. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A.; Woodhams, D.C.; Reinert, L.K.; Vredenburg, V.T.; Briggs, C.J.; Nielsen, P.F.; Conlon, J.M. Antimicrobial peptide defenses of the mountain yellow-legged frog (Rana muscosa). Dev. Comp. Immunol. 2006, 30, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wang, Y.; Wu, C.; Li, X.; Fu, Z.; Yang, M.; Bian, W.; Wang, S.; Song, Y.; Tang, J.; et al. Cathelicidin-OA1, a novel antioxidant peptide identified from an amphibian, accelerates skin wound healing. Sci. Rep. 2018, 8, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Hu, J.Y.; Wang, S.Q. The role of antioxidants in photoprotection: A critical review. J. Am. Acad. Dermatol. 2012, 67, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.; Lee, M.G. Oxidative stress and antioxidant strategies in dermatology. Redox Rep. 2016, 21, 164–169. [Google Scholar] [CrossRef]
- Xu, N.; Chen, G.; Liu, H. Antioxidative Categorization of Twenty Amino Acids Based on Experimental Evaluation. Molecules 2017, 22, 2066. [Google Scholar] [CrossRef]
- Yang, H.; Wang, X.; Liu, X.; Wu, J.; Liu, C.; Gong, W.; Zhao, Z.; Hong, J.; Lin, D.; Wang, Y.; et al. Antioxidant peptidomics reveals novel skin antioxidant system. Mol. Cell. Proteom. 2009, 8, 571–583. [Google Scholar] [CrossRef]
- Wang, X.; Ren, S.; Guo, C.; Zhang, W.; Zhang, X.; Zhang, B.; Li, S.; Ren, J.; Hu, Y.; Wang, H. Identification and functional analyses of novel antioxidant peptides and antimicrobial peptides from skin secretions of four East Asian frog species. Acta Biochim. Biophys. Sin. 2017, 49, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Tevini, M. UV-B Radiation and Ozone Depletion-Effects on Humans, Animals, Plants, Microorganisms, and Materials, 1st ed.; Lewis Publishers: Boca Raton, FL, USA, 1993. [Google Scholar]
- Darr, D.; Fridovich, I. Free radicals in cutaneous biology. J. Investig. Dermatol. 1994, 102, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, A.R.; Belden, L.K. Amphibian defenses against ultraviolet-B radiation. Evol. Dev. 2003, 5, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, Y.; Zhang, Y.; Lee, W.H.; Zhang, Y. Rich diversity and potency of skin antioxidant peptides revealed a novel molecular basis for high-altitude adaptation of amphibians. Sci. Rep. 2016, 6, 19866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Hong, J.; Yang, H.; Wu, J.; Ma, D.; Li, D.; Lin, D.; Lai, R. Frog skins keep redox homeostasis by antioxidant peptides with rapid radical scavenging ability. Free Radic. Biol. Med. 2010, 48, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Lee, W.H.; Gao, Z.; Zhang, W.; Peng, M.; Sun, T.; Gao, Y. Protective effects of antioxidin-RL from Odorrana livida against ultraviolet B-irradiated skin photoaging. Peptides 2018, 101, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Lanoo, M. Amphibian Declines, 1st ed.; University of California Press: Berkeley, CA, USA, 2005. [Google Scholar]
- Kolbert, E. The Sixth Exctinction: An Unnatural History, 1st ed.; Henry Holt and Company: New York, NY, USA, 2014. [Google Scholar]
- Crump, M.L. Why are some species in decline but others not. In Amphibian Declines, 1st ed.; Lanoo, M., Ed.; University of California Press: Berkeley, CA, USA, 2005; pp. 7–9. [Google Scholar]
- Bovo, R.; Navas, C.; Tejedo, M.; Valença, S.; Gouveia, S. Ecophysiology of Amphibians: Information for Best Mechanistic Models. Diversity 2018, 10, 118. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A. The role of amphibian antimicrobial peptides in protection of amphibians from pathogens linked to global amphibian declines. Biochim. Biophys. Acta 2009, 1788, 1593–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Catenazzi, A. State of the World’s Amphibians. Ann. Rev. Environ. Resour. 2015, 40, 91–119. [Google Scholar] [CrossRef]
- Bletz, M.C.; Kelly, M.; Sabino-Pinto, J.; Bales, E.; Van Praet, S.; Bert, W.; Boyen, F.; Vences, M.; Steinfartz, S.; Pasmans, F.; et al. Disruption of skin microbiota contributes to salamander disease. Proc. Biol. Sci. 2018, 285, 20180758. [Google Scholar] [CrossRef] [PubMed]
- Withers, P.C.; Guppy, M. Do Australian desert frogs co-accumulate counteracting solutes with urea during aestivation? J. Exp. Biol. 1996, 199, 1809–1816. [Google Scholar] [PubMed]
- Ufnal, M.; Zadlo, A.; Ostaszewski, R. TMAO: A small molecule of great expectations. Nutrition 2015, 31, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Pouokam, E.; Althaus, M. Epithelial Electrolyte Transport Physiology and the Gasotransmitter Hydrogen Sulfide. Oxid. Med. Cell. Longev. 2016, 2016, 4723416. [Google Scholar] [CrossRef] [PubMed]
- Tomasova, L.; Konopelski, P.; Ufnal, M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules 2016, 21, 1558. [Google Scholar] [CrossRef] [PubMed]
- Vergani, L.; Vecchione, G.; Baldini, F.; Grasselli, E.; Voci, A.; Portincasa, P.; Ferrari, P.F.; Aliakbarian, B.; Casazza, A.A.; Perego, P. Polyphenolic extract attenuates fatty acid-induced steatosis and oxidative stress in hepatic and endothelial cells. Eur. J. Nutr. 2018, 57, 1793–1805. [Google Scholar] [CrossRef] [PubMed]
- Garnier, J.; Gibrat, J.F.; Robson, B. GOR method for predicting protein secondary structure from amino acid sequence. Methods Enzymol. 1996, 266, 540–553. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Peptide Name | AA Sequences | Species | Length (AA) | Secondary Structure Prediction | Ref. |
|---|---|---|---|---|---|
| AH-90 | ATAWDFGPHGLLPIRPIRIRPLCG | Odorrana grahami | 24 | Ee: 29.17% Cc 70.83% | [32] |
| Alytesin | Pyr-GRLGTQWAVGHLM | Alytes obstetricans | 14 | Ee: 46.15% Cc: 53.85% | [28] |
| ARPs/ERPs | - | Rana chensinensis | 2–20 | - | [37] |
| Bombesin | Pyr-QRLGNQWAVGHLM | Bombina bombina Bombina variegata Bombina orientalis | 14 | Ee: 38.46% Cc: 61.54% | [27] |
| Bm-TFF2 | GFPIYEIDNRPGCYVDPAERVAC AGAGVTKAECKAKGCCFISARR NTIWCFKLKESADAWKCAVPM NTRVACAGAGVTPAECKGKGC CFNSSYYGTVWCFKPQE | Bombina maxima | 104 | Ee: 39.42% Cc: 60.58% | [29,30] |
| Cathelicidin-OA1 | IGRDPTWSHLAASCLKCIFDDLPKTHN | Odorrana andersonii | 27 | Hh: 29.63% Ee: 7.41% Cc: 62.96% | [49] |
| CW49 | APFRMGICTTN | Odorrana grahami | 11 | Ee: 45.45% Cc: 54.55% | [36] |
| OA-GL21 | GLLSGHYGRVVSTQSGHYGRG | Odorrana andersonii | 21 | Ee: 52.38% Cc: 47.62% | [34] |
| OM-LV20 | LVGKLLKGAVGDVCGLLPIC | Odorrana margaretae | 20 | Ee: 45.00% Cc: 55.00% | [35] |
| pbCGRP | SCDTSTCATQRLADFLSRSGGIGSPDFVPTDVSANSF | Pyllomedusa bicolor | 37 | Hh: 18.92% Ee: 13.51% Cc: 67.57% | [9,31] |
| Temporins-A | FLPLIGRVLSGIL | Rana temporaria | 13 | Ee: 69.23% Cc: 30.77% | [33] |
| Temporins-B | LLPIVGNLLKSLL | Rana temporaria | 13 | Ee: 30.77% Cc: 69.23% | [33] |
| Peptide | AA Sequences | Species | Length (AA) | Secondary Structure Prediction | Ref. |
|---|---|---|---|---|---|
| Andersonin-C1 | TSRCIFYRRKKCS | Odorrana margaratae | 13 | Ee: 53.85% Cc: 46.15% | [9] |
| Andersonin-G1 | KEKLKLKAKAPKCYNDKLACT | Odorrana andersonii | 21 | Ee: 23.81% Cc: 76.19% | [9] |
| Andersonin-H3 | VAIYGRDDRSDVCRQVQHNWLVCDTY | Odorrana margaratae | 26 | Ee: 42.31% Cc: 57.69% | [9] |
| Andersonin-N1 | ENMFNIKSSVESDSFWG | Odorrana margaratae | 17 | Ee: 52.94% Cc: 47.06% | [9] |
| Andersonin-R1 | ENAEEDEVLMENLFCSYIVGSADSFWT | Odorrana margaratae | 27 | Hh: 18.52% Ee: 33.33% Cc: 48.15% | [9] |
| Antioxidin-RP1 | AMRLTYNKPCLYGT | Rana pleuraden | 14 | Ee: 28.57% Cc: 71.43% | [9,53] |
| Antioxidin-RP2 | SMRLTYNKPCLYGT | Rana pleuraden | 14 | Ee: 28.57% Cc: 71.43% | [9,53] |
| APBMH | LEQQVDDLEGSLEQEKK | Rana catesbeiana | 17 | Hh: 35.29% Ee: 11.76% Cc: 52.94% | [9] |
| APBSP | LEELEEELEGCE | Rana catesbeiana | 12 | Ee: 16.67% Cc: 83.33% | [9] |
| Hejiangin-A1 | RFIYMKGFGKPRFGKR | Odorrana hejiangensis | 16 | Ee: 31.25% Cc: 68.75% | [9] |
| Hejiangin-E1 | SADQTGMNKAALSPIRFISKSV | Odorrana hejiangensis | 22 | Ee: 28.57% Cc: 71.43% | [9] |
| Hejiangin-F1 | IPWKLPATFRPVERPFSKPFCRKD | Odorrana hejiangensis | 24 | Ee: 16.67% Cc: 83.33% | [9] |
| Japonicin-1Npa | FLLFPLMCKIQGKC | Nanorana parkeri | 14 | Ee: 35.71% Cc: 64.29% | [9] |
| Japonicin-1Npb | FVLPLVMCKILRKC | Nanorana parkeri | 14 | Ee: 50.00% Cc: 50.00% | [9] |
| Lividin-D1 | KNNFCQVLYVWLLRLGKQCFVKFSKDVET | Odorrana livida | 29 | Ee: 51.72% Cc: 48.28% | [9] |
| Macrotympanain A1 | FLPGLECVW | Odorrana macrotympana | 9 | Ee: 33.33% Cc: 66.67% | [9] |
| Margaratain-A1 | VTPPWARIYYGCAKA | Odorrana margaratae | 15 | Ee: 33.33% Cc: 66.67% | [9] |
| Margaratain-B1 | FFSTSCRSGC | Odorrana margaratae | 10 | Ee: 60.00% Cc: 40.00% | [9] |
| Margaratain-C1 | GMLKWKNDFFHFLQWLLISCQNYFVK | Odorrana margaratae | 26 | Ee: 50.00% Cc: 50.00% | [9] |
| Nigroain-B-MS1 | CVVSSGWKWNYKIRCKLTGNC | Hylarana maosuoensis | 21 | Ee: 47.62% Cc: 52.38% | [54] |
| Odorranian-A-OA11 | VVKCSYRQGSPDSR | Odorrana margaratae | 14 | Ee: 42.86% Cc: 57.14% | [9] |
| Odorranian-A-OA12 | VVKFSYRKGSPAPQKN | Odorrana margaratae | 16 | Ee: 37.50% Cc:62.50% | [9] |
| Parkerin | GWANTLKNVAGGLCKITGAA | Nanorana parkeri | 20 | Ee: 45.00% Cc: 55.00% | [9] |
| Pleurain-A1 | SIITMTKEAKLPQLWKQIACRLYNTC | Rana pleuraden | 26 | Hh: 19.23% Ee: 30.77% Cc: 50.00% | [53] |
| Pleurain-D1 | FLSGILKLAFKIPSVLCAVLKNC | Rana pleuraden | 23 | Ee: 47.83% Cc: 52.17% | [53] |
| Pleurain-E1 | AKAWGIPPHVIPQIVPVRIRPLCGNV | Rana pleuraden | 26 | Ee: 30.77% Cc:69.23% | [53] |
| Pleurain-G1 | GFWDSVKEGLKNAAVTILNKIKCKISECPPA | Rana pleuraden | 31 | Hh: 45.16% Cc: 54.84% | [53] |
| Pleurain-J1 | FIPGLRRLFATVVPTVVCAINKLPPG | Rana pleuraden | 26 | Ee: 34.62% Cc: 65.38% | [53] |
| Pleurain-K1 | DDPDKGMLKWKNDFFQEF | Rana pleuraden | 18 | E: 22.22% Cc: 77.78% | [53] |
| Pleurain-M1 | GLLDSVKEGLKKVAGQLLDTLKCKISGCTPA | Rana pleuraden | 31 | Hh: 38.71% Ee: 19.35% Cc: 41.94% | [53] |
| Pleurain-N1 | GFFDRIKALTKNVTLELLNTITCKLPVTPP | Rana pleuraden | 30 | Hh: 40.00% Ee: 6.67% Cc: 53.33% | [53] |
| Pleurain-P1 | SFGAKNAVKNGLQKLRNQCQANNYQGPFCDIFKKNP | Rana pleuraden | 36 | Hh: 33.33% Ee: 19.44% Cc: 47.22% | [53] |
| Pleurain-R1 | CVHWMTNTARTACIAP | Rana pleuraden | 16 | Ee: 37.50% Cc: 62.50% | [53] |
| Schmackerin-C1 | AAPRGGKGFFCKLFKDC | Odorrana schmackeri | 17 | Ee: 35.29% Cc: 64.71% | [9] |
| Tiannanin-A1 | LLPPWLRPRNG | Odorrana tiannanensis | 11 | Ee: 36.36% Cc: 63.64% | [9] |
| Wuchuanin-A1 | APDRPRKFCGILG | Odorrana wuchuanensis | 13 | Ee: 38.46% Cc: 61.54% | [9] |
| Wuchuanin-C1 | VFLGNIVSMGKKI | Odorrana wuchuanensis | 13 | Ee: 53.85% Cc: 46.15% | [9] |
| Wuchuanin-D1 | DAAVEPELYHWGKVWLPN | Odorrana wuchuanensis | 18 | Ee: 27.78% Cc: 72.22% | [9] |
| Wuchuanin-E1 | CVDIGFSPTGKRPPFCPYPG | Odorrana wuchuanensis | 20 | Ee: 10.00% Cc: 90.00% | [9] |
| Peptide | AA Sequences | Species | Length (AA) | Secondary Structure Prediction | Ref. |
|---|---|---|---|---|---|
| Andersonin-AOP 1 | FLPGLECVM | Odorrana andersonii | 9 | Ee: 44.44% Cc: 55.56% | [58] |
| Antioxidin-RL | AMRLTYNRPCIYAT | Odorrana livida | 14 | Ee: 28.57% Cc: 71.43% | [59,60] |
| Daiyunin-1 | CGYKYGCMVKVDR | Amolops daiyunensis | 13 | Ee: 30.77% Cc: 69.23% | [59] |
| Maosonensis-1MS1 | QYRPGSFGPLNQK | Hylarana maosuoensis | 13 | Ee: 23.08% Cc: 76.92% | [54] |
| Odorranaopin-MS2 | DYSIRTRLHQESSRNVF | Hylarana maosuoensis | 17 | Ee: 52.94% Cc: 47.06% | [54] |
| Pleskein-2 | FFLLPIPNDVKCKVLGICKS | Nanorana pleskei | 20 | Ee: 35.00% Cc: 65.00% | [54] |
| Ranacyclin-HB1 | GAPKGCWTKSYPPQPCFGKK | Pelophylax hubeiensis | 20 | Ee: 15.00% Cc: 85.00% | [9] |
| Wuchuanin-AOP 5 | TVWGFRPSKPPSGYR | Odorrana wuchuanensi | 15 | Ee: 20.00% Cc: 80.00% | [58] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demori, I.; El Rashed, Z.; Corradino, V.; Catalano, A.; Rovegno, L.; Queirolo, L.; Salvidio, S.; Biggi, E.; Zanotti-Russo, M.; Canesi, L.; et al. Peptides for Skin Protection and Healing in Amphibians. Molecules 2019, 24, 347. https://doi.org/10.3390/molecules24020347
Demori I, El Rashed Z, Corradino V, Catalano A, Rovegno L, Queirolo L, Salvidio S, Biggi E, Zanotti-Russo M, Canesi L, et al. Peptides for Skin Protection and Healing in Amphibians. Molecules. 2019; 24(2):347. https://doi.org/10.3390/molecules24020347
Chicago/Turabian StyleDemori, Ilaria, Zeinab El Rashed, Viola Corradino, Annamaria Catalano, Leila Rovegno, Linda Queirolo, Sebastiano Salvidio, Emanuele Biggi, Matteo Zanotti-Russo, Laura Canesi, and et al. 2019. "Peptides for Skin Protection and Healing in Amphibians" Molecules 24, no. 2: 347. https://doi.org/10.3390/molecules24020347
APA StyleDemori, I., El Rashed, Z., Corradino, V., Catalano, A., Rovegno, L., Queirolo, L., Salvidio, S., Biggi, E., Zanotti-Russo, M., Canesi, L., Catenazzi, A., & Grasselli, E. (2019). Peptides for Skin Protection and Healing in Amphibians. Molecules, 24(2), 347. https://doi.org/10.3390/molecules24020347