Inhibition of T Cell Receptor Activation by Semi-Synthetic Sesquiterpene Lactone Derivatives and Molecular Modeling of Their Interaction with Glutathione and Tyrosine Kinase ZAP-70

 ,
,  , , ,
, , , 
Abstract
:
1. Introduction
2. Results and Discussion
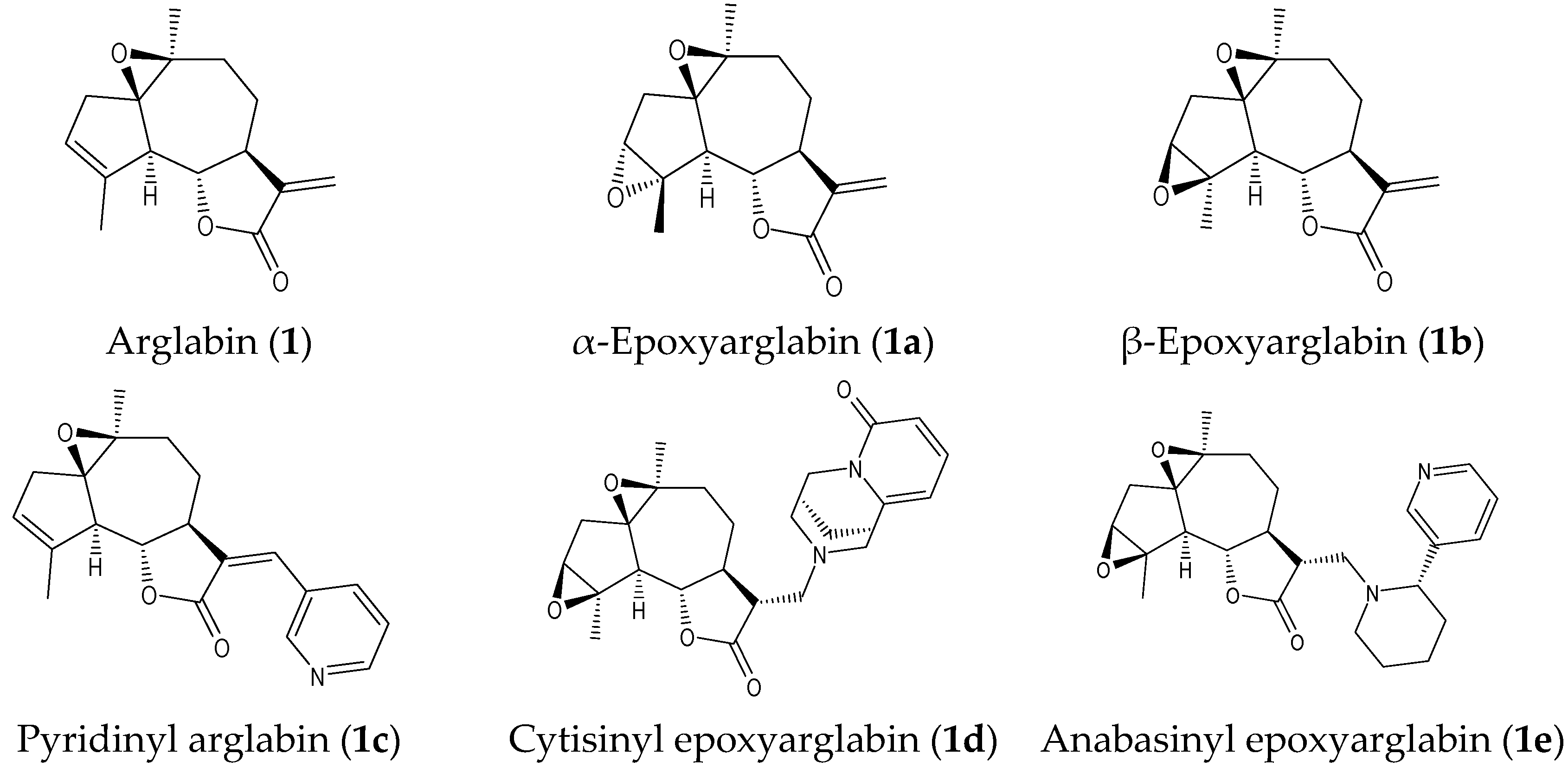
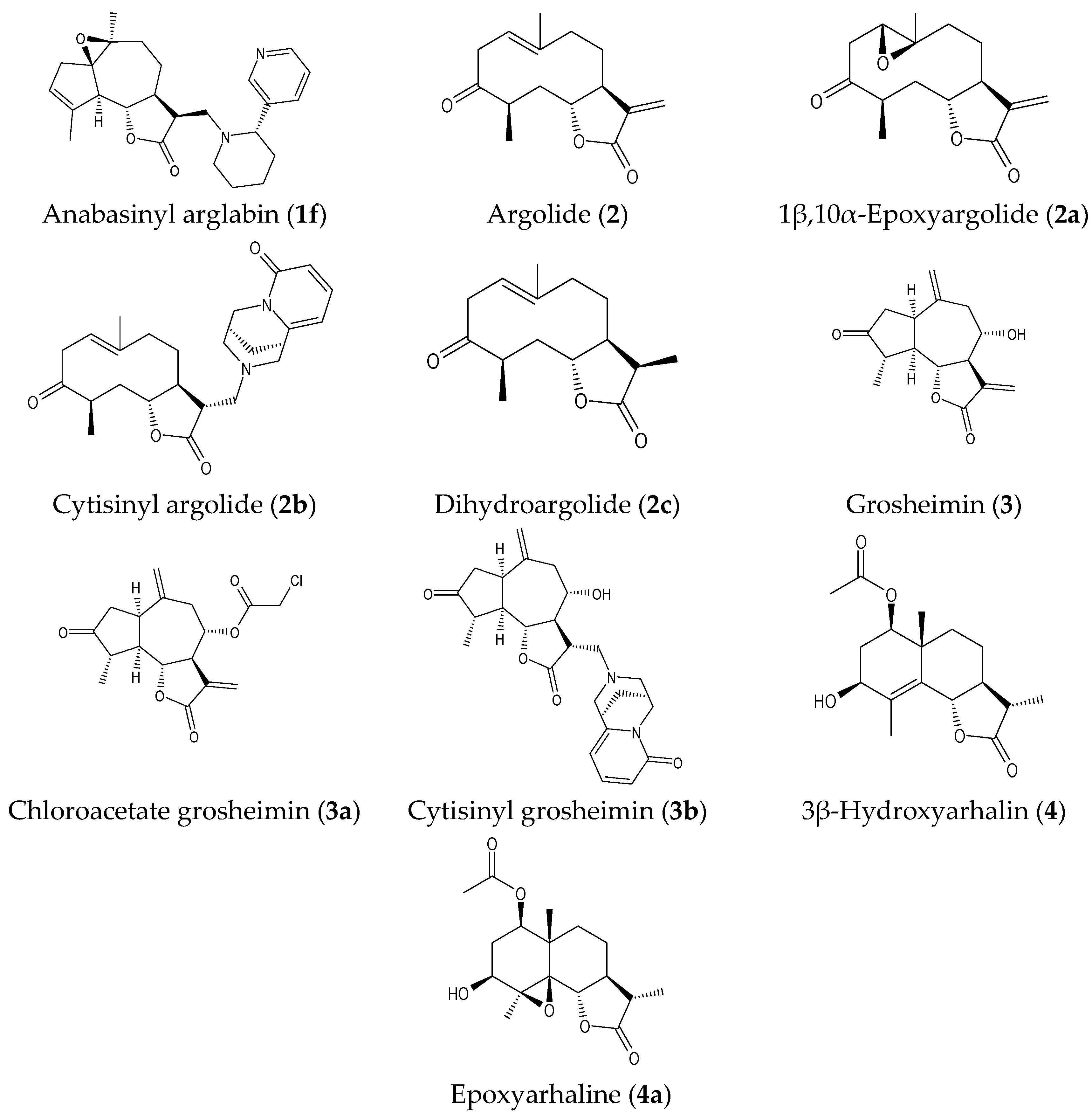
2.1. Isolation of Parent Natural SLs and Synthesis of Their Derivatives
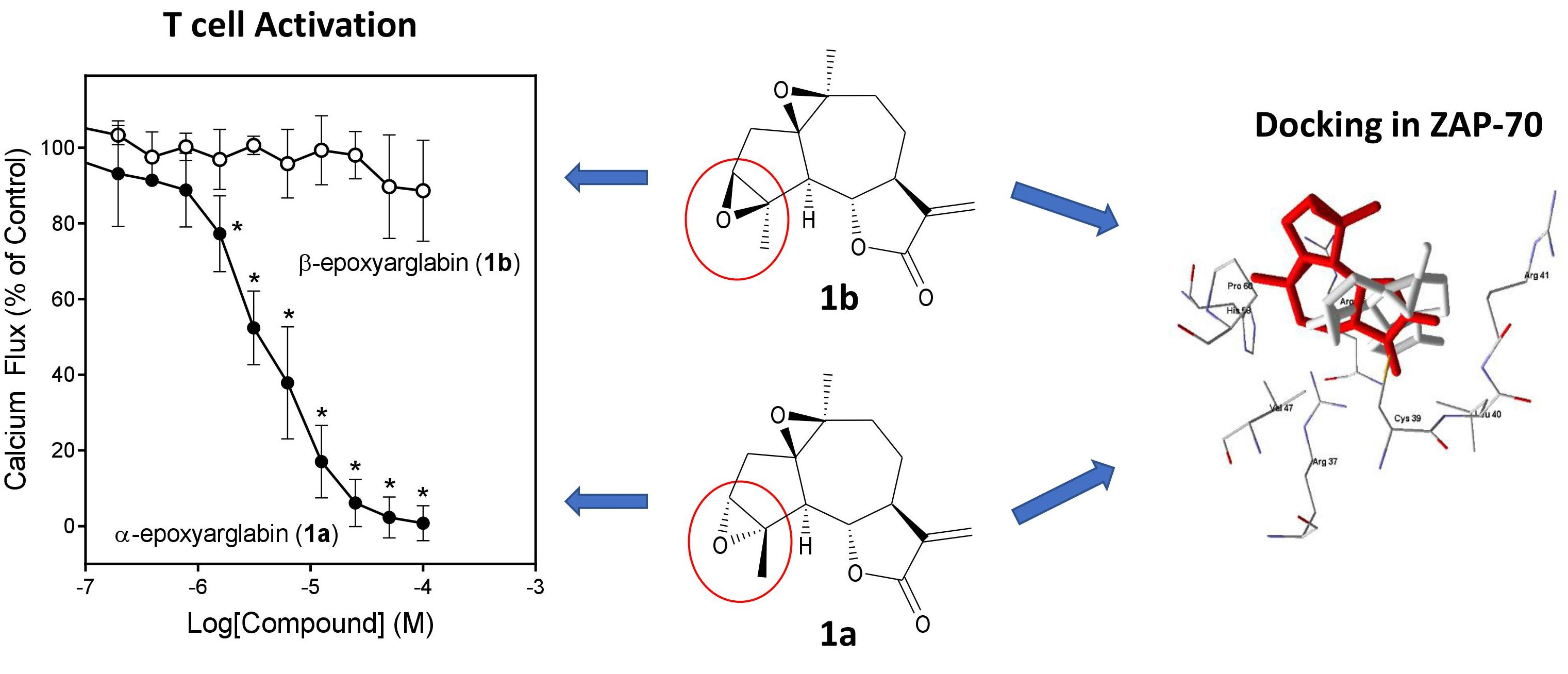
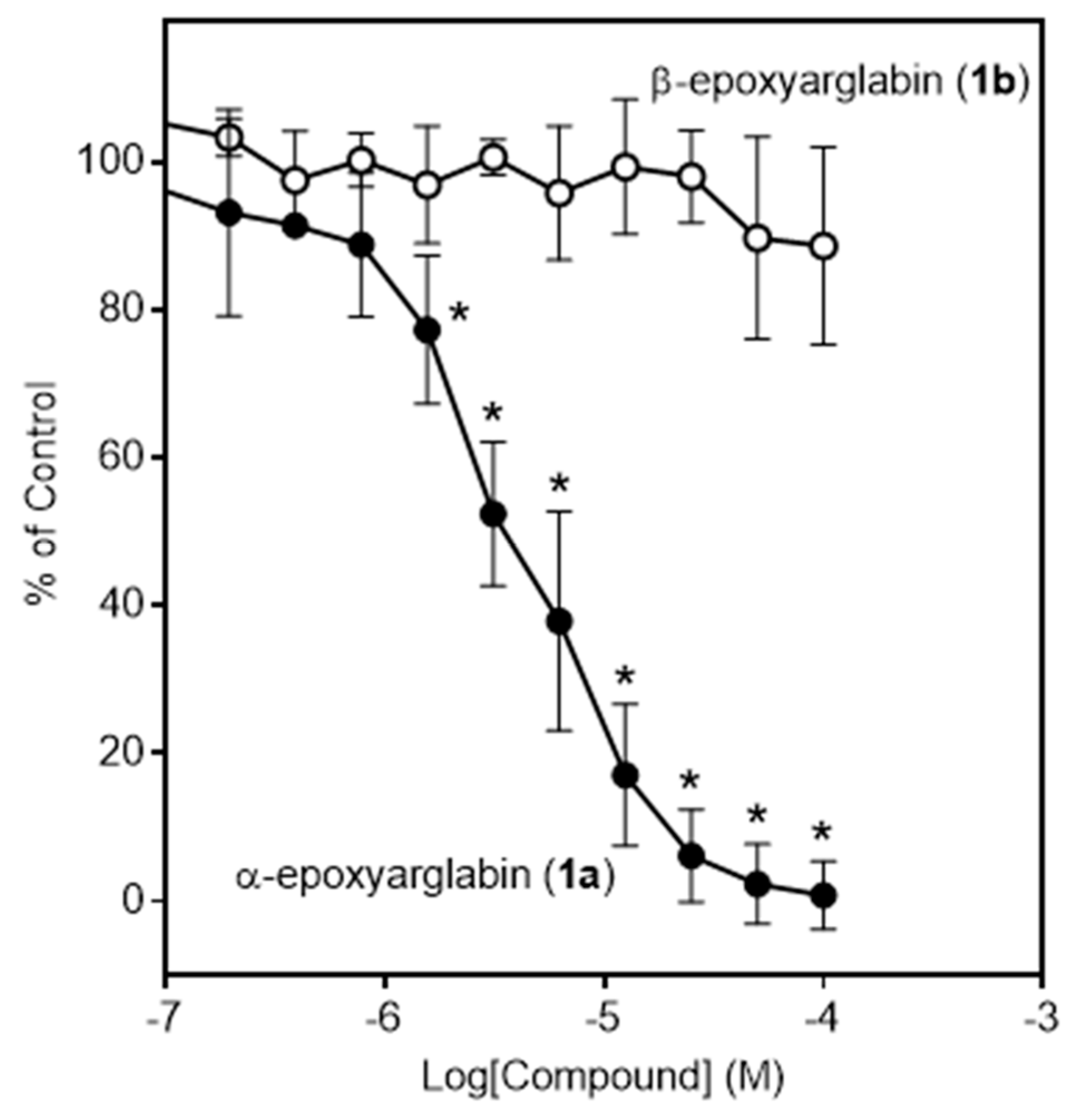
2.2. Effects of Semi-Synthetic SL Derivatives on TCR Activation
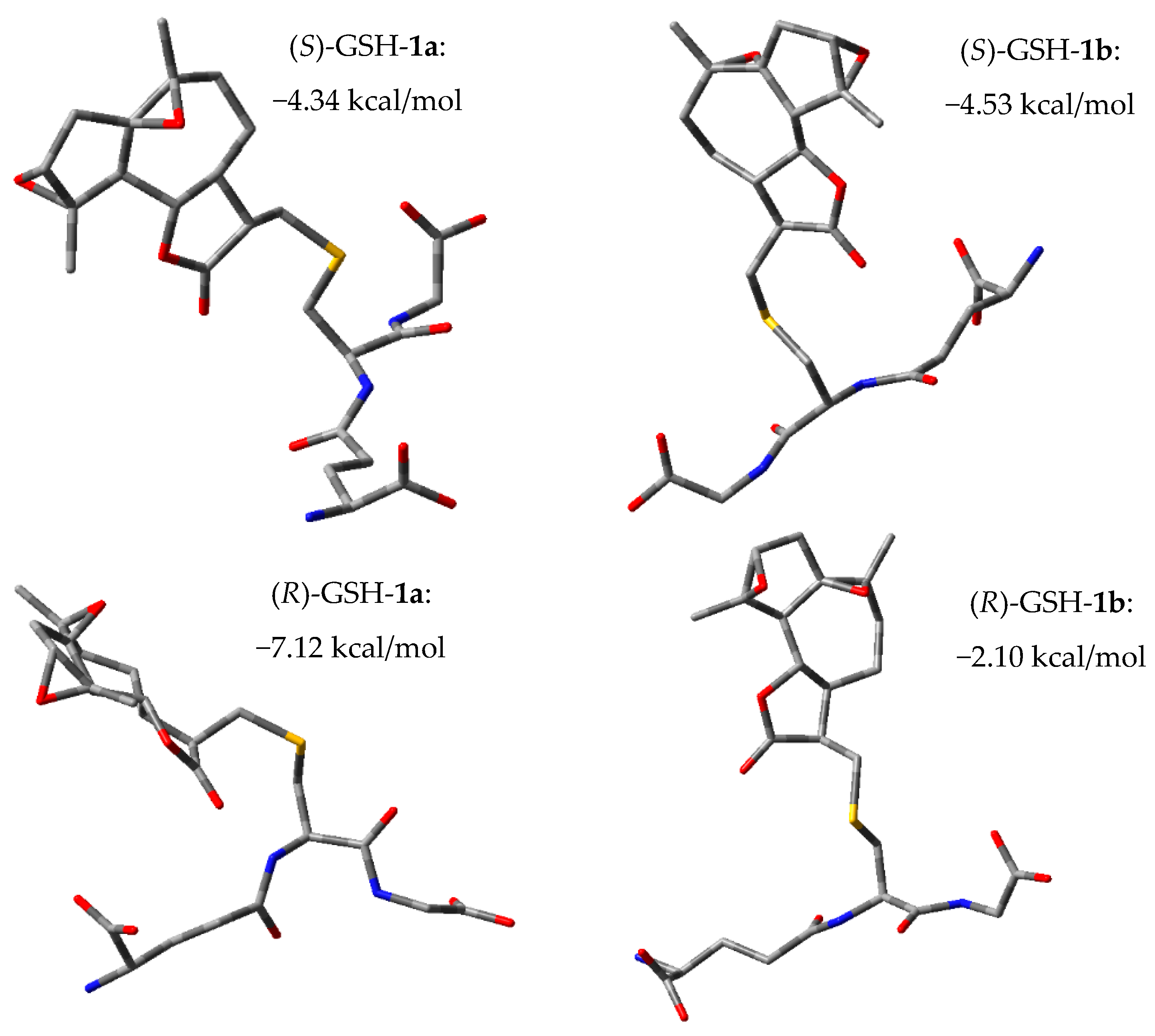
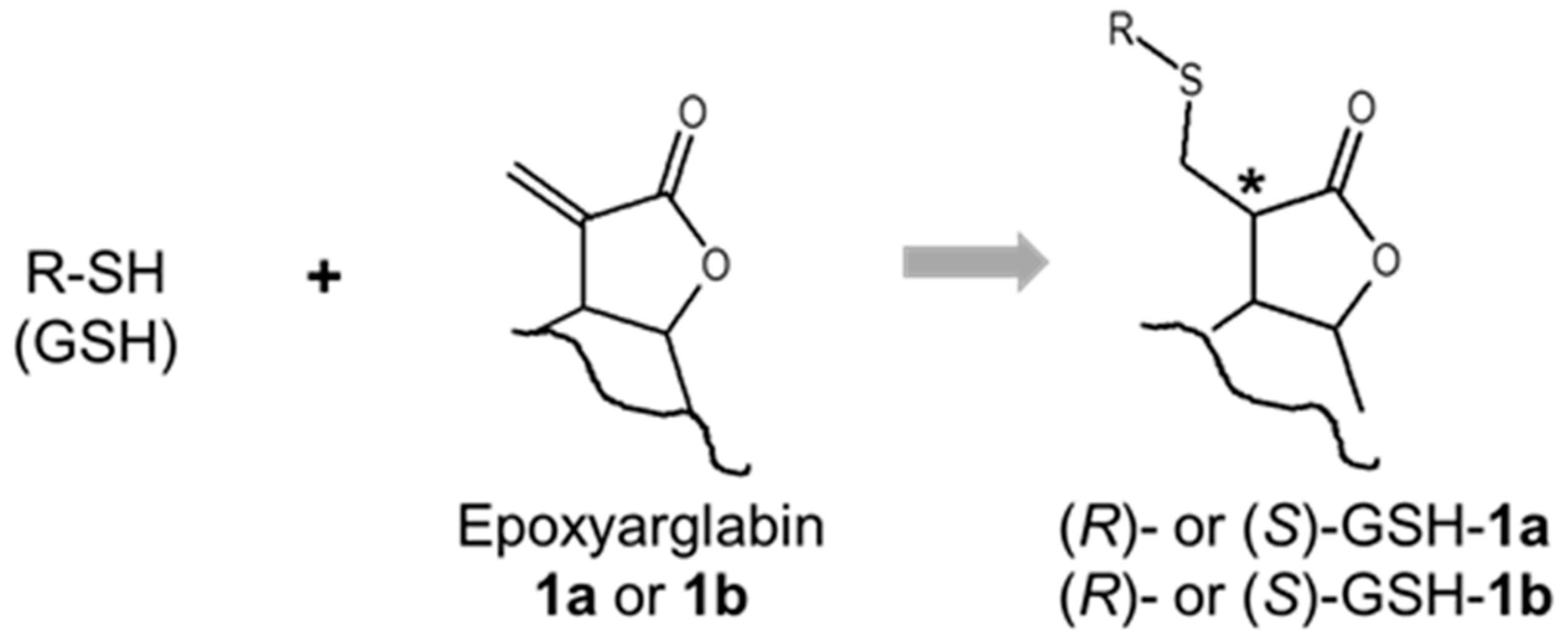
2.3. Glutathione (GSH) Reactivity of Semi-Synthetic SL Derivatives
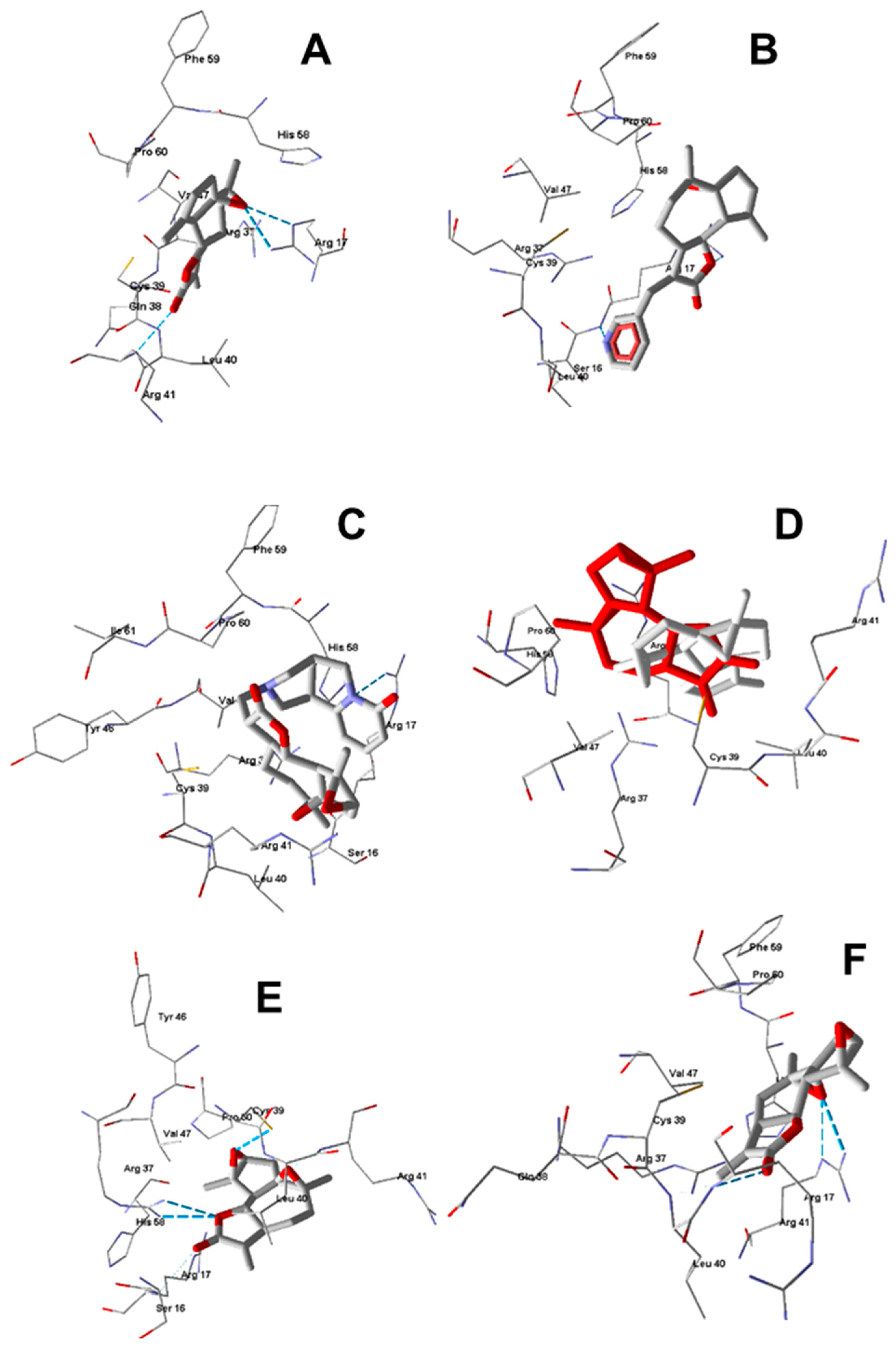
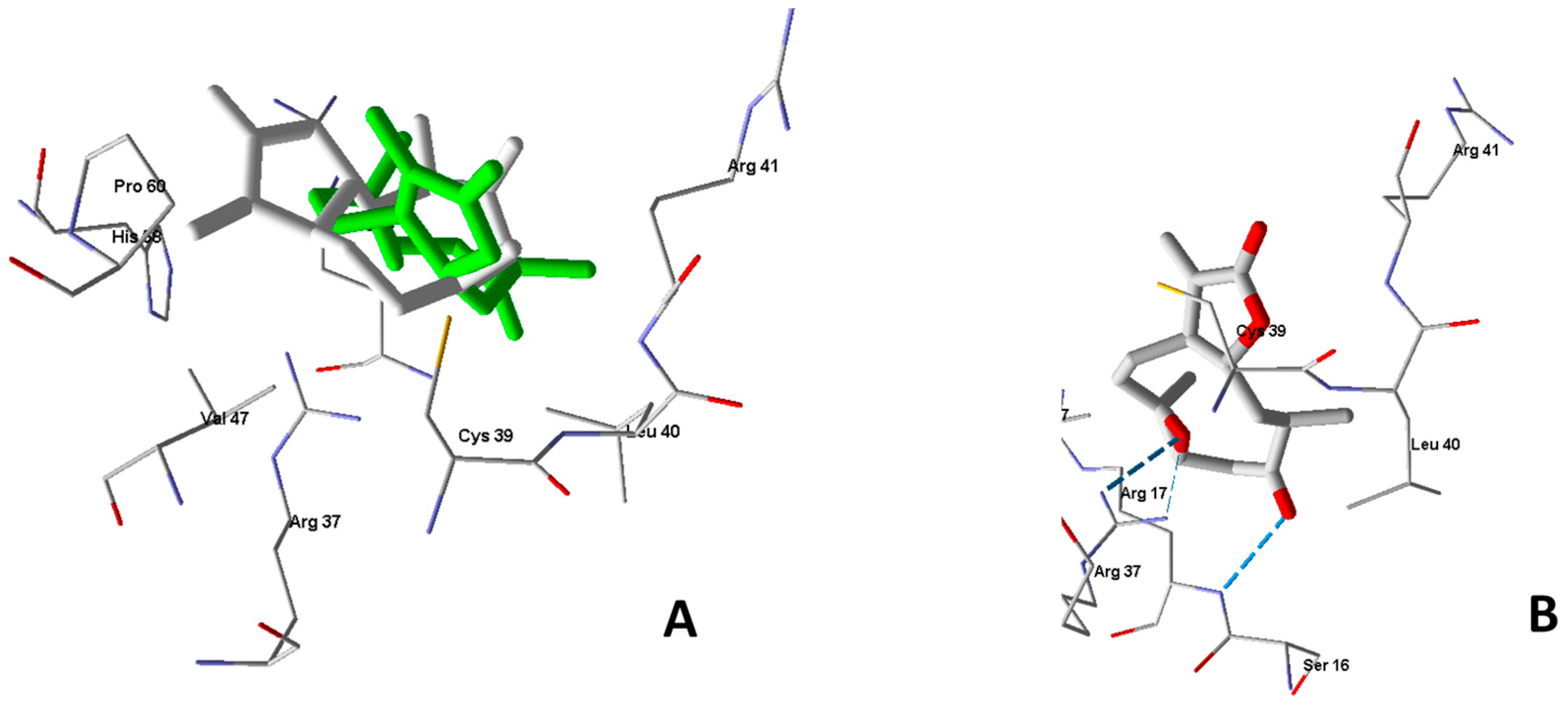
2.4. Molecular Modeling
3. Materials and Methods
3.1. Plant Material and Isolation of Natural SLs
3.2. Synthesis of Derivatives from Natural SLs
3.3. Materials for Biological Assays
3.4. Cell Culture
3.5. Isolation of Human Neutrophils
3.6. ERK1/2 Enzyme-Linked Immunosorbent Assay (ELISA)
3.7. Ca2+ Mobilization Assay
3.8. Assessment of Compound Cytotoxicity
3.9. Glutathione (GSH) Assay
3.10. DFT Calculations
3.11. Molecular Docking
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DMSO | dimethyl sulfoxide |
| FBS | fetal bovine serum |
| FPR | N-formyl peptide receptor |
| HBSS | Hanks’ balanced salt solution |
| ITAM | Immune-receptor tyrosine-based activation motif |
| TCR | T cell antigen receptor |
| ERK | extracellular signal-regulated kinase |
| MAPK | mitogen-activated protein kinases |
| GSH | glutathione |
| ELISA | enzyme-linked immunosorbent assay |
References
- Sauer, E.L.; Cloake, N.C.; Greer, J.M. Taming the TCR: Antigen-specific immunotherapeutic agents for autoimmune diseases. Int. Rev. Immunol. 2015, 34, 460–485. [Google Scholar] [CrossRef] [PubMed]
- Visperas, P.R.; Wilson, C.G.; Winger, J.A.; Yan, Q.; Lin, K.; Arkin, M.R.; Weiss, A.; Kuriyan, J. Identification of inhibitors of the association of ZAP-70 with the T cell receptor by high-throughput screen. SLAS Discov. 2017, 22, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Li, Y.; Ratnayake, R.; Luo, D.; Lo, J.; Reibenspies, J.H.; Xu, Z.S.; Clare-Salzler, M.J.; Ye, T.; Paul, V.J.; et al. Discovery, total synthesis and key structural elements for the immunosuppressive activity of cocosolide, a symmetrical glycosylated macrolide dimer from marine cyanobacteria. Chem. Eur. J. 2016, 22, 8158–8166. [Google Scholar] [CrossRef] [PubMed]
- Jelic, D.; Tatic, I.; Trzun, M.; Hrvacic, B.; Brajsa, K.; Verbanac, D.; Tomaskovic, M.; Culic, O.; Antolovic, R.; Glojnaric, I.; et al. Porphyrins as new endogenous anti-inflammatory agents. Eur. J. Pharmacol. 2012, 691, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Lee, Y.G.; Park, H.J.; Lee, J.A.; Kim, H.J.; Hwang, J.K.; Choi, J.M. Piceatannol inhibits effector T cell functions by suppressing tcr signaling. Int. Immunopharmacol. 2015, 25, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Kirpotina, L.N.; Mitchell, P.T.; Kishkentaeva, A.S.; Shaimerdenova, Z.R.; Atazhanova, G.A.; Adekenov, S.M.; Quinn, M.T. The natural natural sesquiterpene lactones arglabin, grosheimin, agracin, parthenolide, and estafiatin inhibit T cell receptor (TCR) activation. Phytochemistry 2018, 146, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, M.; Trewin, H.; Gawthrop, F.; Wagstaff, C. Sesquiterpenoids lactones: Benefits to plants and people. Int. J. Mol. Sci. 2013, 14, 12780–12805. [Google Scholar] [CrossRef]
- Hou, L.; Huang, H. Immune suppressive properties of artemisinin family drugs. Pharmacol. Ther. 2016, 166, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Yu, J.; Kinghorn, A.D. Development of anticancer agents from plant-derived sesquiterpene lactones. Curr. Med. Chem. 2016, 23, 2397–2420. [Google Scholar] [CrossRef]
- Adekenov, S.M. Chemical modification of arglabin and biological activity of its new derivatives. Fitoterapia 2016, 110, 196–205. [Google Scholar] [CrossRef]
- Guo, Z.R. The modification of natural products for medical use. Acta Pharm. Sin. B 2017, 7, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thome, R.; de Carvalho, A.C.; da Costa, T.A.; Ishikawa, L.L.W.; Fraga-Silva, T.F.D.; Sartori, A.; de Oliveira, A.L.R.; Verinaud, L. Artesunate ameliorates experimental autoimmune encephalomyelitis by inhibiting leukocyte migration to the central nervous system. CNS Neurosci. Ther. 2016, 22, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Nasim, S.; Crooks, P.A. Antileukemic activity of aminoparthenolide analogs. Bioorg. Med. Chem. Lett. 2008, 18, 3870–3873. [Google Scholar] [CrossRef] [PubMed]
- Neelakantan, S.; Nasim, S.; Guzman, M.L.; Jordan, C.T.; Crooks, P.A. Aminoparthenolides as novel anti-leukemic agents: Discovery of the NF-κB inhibitor, DMAPT (LC-1). Bioorg. Med. Chem. Lett. 2009, 19, 4346–4349. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Tang, J.J.; Zhang, C.C.; Tian, J.M.; Guo, J.T.; Zhang, Q.; Li, H.; Gao, J.M. Semisynthesis and in vitro cytotoxic evaluation of new analogues of 1-O-acetylbritannilactone, a sesquiterpene from Inula britannica. Eur. J. Med. Chem. 2014, 80, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Ding, Y.H.; Wang, P.P.; Zhang, Q.; Chen, Y. Protection-group-free semisyntheses of parthenolide and its cyclopropyl analogue. J. Org. Chem. 2013, 78, 10512–10518. [Google Scholar] [CrossRef] [PubMed]
- Arantes, F.F.P.; Barbosa, L.C.A.; Maltha, C.R.A.; Demuner, A.J.; da Costa, P.M.; Ferreira, J.R.O.; Costa-Lotufo, L.V.; Moraes, M.O.; Pessoa, C. Synthesis of novel α-santonin derivatives as potential cytotoxic agents. Eur. J. Med. Chem. 2010, 45, 6045–6051. [Google Scholar] [CrossRef]
- Han, C.H.; Barrios, F.J.; Riofski, M.V.; Colby, D.A. Semisynthetic derivatives of sesquiterpene lactones by palladium-catalyzed arylation of the α-methylene-γ-lactone substructure. J. Org. Chem. 2009, 74, 7176–7179. [Google Scholar] [CrossRef]
- Liu, S.X.; Liu, H.; Yan, W.Y.; Zhang, L.; Bai, N.S.; Ho, C.T. Studies on 1-o-acetylbritannilactone and its derivative, (2-O-butyloxime-3-phenyl)-propionyl-1-O-acetylbritannilactone ester. Bioorg. Med. Chem. Lett. 2004, 14, 1101–1104. [Google Scholar] [CrossRef]
- Tang, J.J.; He, Q.R.; Dong, S.; Guo, X.; Wang, Y.G.; Lei, B.L.; Tian, J.M.; Gao, J.M. Diversity modification and structure-activity relationships of two natural products 1 β-hydroxy alantolactone and ivangustin as potent cytotoxic agents. Sci. Rep. 2018, 8, 1722. [Google Scholar] [CrossRef]
- Torchia, M.L.G.; Dutta, D.; Mittelstadt, P.R.; Guha, J.; Gaida, M.M.; Fish, K.; Barr, V.A.; Akpan, I.O.; Samelson, L.E.; Tagad, H.D.; et al. Intensity and duration of tcr signaling is limited by p38 phosphorylation of ZAP-70T293 and destabilization of the signalosome. Proc. Natl. Acad. Sci. USA 2018, 115, 2174–2179. [Google Scholar] [CrossRef] [PubMed]
- Katz, Z.B.; Novotna, L.; Blount, A.; Lillemeier, B.F. A cycle of ZAP70 kinase activation and release from the TCR amplifies and disperses antigenic stimuli. Nat. Immunol. 2017, 18, 86–95. [Google Scholar] [CrossRef]
- Adekenov, S.M. Sesquiterpene lactones from endemic species of the family Asteraceae. Chem. Nat. Compd. 2013, 1, 158–162. [Google Scholar] [CrossRef]
- Adekenov, S.M.; Shaimerdenova, Z.R.; Gatilov, Y.V.; Atazhanova, G.A. Two new sesquiterpene lactones from Arthemisia halophile. Chem. Nat. Compd. 2017, 53, 241–245. [Google Scholar] [CrossRef]
- Adekenova, A.S.; Sakenova, P.Y.; Ivasenko, S.A.; Khabarov, I.A.; Adekenov, S.M.; Berthod, A. Gram-scale purification of two sesquiterpene lactones from Chartolepsis intermedia boiss. Chromatographia 2016, 79, 37–43. [Google Scholar] [CrossRef]
- Adekenov, S.M.; Aituganov, K.A.; Turdybekov, K.M.; Lindeman, S.V.; Struchkov, Y.T. Molecular and crystal-structure of argolide germacranolide from Artemisia glabella. Khim. Prir. Soedin. 1991, 5, 653–657. [Google Scholar] [CrossRef]
- Adekenov, S.M.; Aituganov, K.A.; Turdybekov, K.M.; Lindeman, S.V.; Struchkov, Y.T.; Bagryanskaya, I.Y.; Gatilov, Y.V. Reactions by binary bond and arglabin epoxy group. Khim. Prir. Soedin. 1991, 1, 33–42. [Google Scholar]
- Turdybekov, K.M.; Edilbaeva, T.T.; Raldugin, V.A.; Shakirov, M.M.; Kulyasov, A.T.; Adekenov, S.M. Molecular and crystalline structure of argolide epoxide. Khim. Prir. Soedin. 1998, 1, 60–64. [Google Scholar]
- Adekenov, S.M.; Kishkentaeva, A.S.; Shaimerdenova, Z.R.; Atazhanova, G.A. Bimolecular compounds based on natural metabolites. Chem. Nat. Compd. 2018, 54, 464–470. [Google Scholar] [CrossRef]
- Ishikawa, J.; Ohga, K.; Yoshino, T.; Takezawa, R.; Ichikawa, A.; Kubota, H.; Yamada, T. A pyrazole derivative, YM-58483, potently inhibits store-operated sustained Ca2+ influx and IL-2 production in T lymphocytes. J. Immunol. 2003, 170, 4441–4449. [Google Scholar] [CrossRef]
- Cerveira, J.; Begum, J.; Barros, R.D.; van der Veen, A.G.; Filby, A. An imaging flow cytometry-based approach to measuring the spatiotemporal calcium mobilisation in activated T cells. J. Immunol. Methods 2015, 423, 120–130. [Google Scholar] [CrossRef]
- Kim, J.-E.; White, F.M. Quantitative analysis of phosphotyrosine signaling networks triggered by CD3 and CD28 costimulation in Jurkat cells. J. Immunol. 2006, 176, 2833–2843. [Google Scholar] [CrossRef]
- Gringhuis, S.I.; Papendrecht-van der Voort, E.A.; Leow, A.; Nivine Levarht, E.W.; Breedveld, F.C.; Verweij, C.L. Effect of redox balance alterations on cellular localization of LAT and downstream T-cell receptor signaling pathways. Mol. Cell. Biol. 2002, 22, 400–411. [Google Scholar] [CrossRef]
- Itoh, T.; Ohguchi, K.; Nozawa, Y.; Akao, Y. Intracellular glutathione regulates sesquiterpene lactone-induced conversion of autophagy to apoptosis in human leukemia HL60 cells. Anticancer Res. 2009, 29, 1449–1457. [Google Scholar]
- Scarponi, C.; Butturini, E.; Sestito, R.; Madonna, S.; Cavani, A.; Mariotto, S.; Albanesi, C. Inhibition of inflammatory and proliferative responses of human keratinocytes exposed to the sesquiterpene lactones dehydrocostuslactone and costunolide. PLoS ONE 2014, 9, e107904. [Google Scholar] [CrossRef]
- Schmidt, T.J.; Lyss, G.; Pahl, H.L.; Merfort, I. Helenanolide type sesquiterpene lactones. Part 5: The role of glutathione addition under physiological conditions. Bioorg. Med. Chem. 1999, 7, 2849–2855. [Google Scholar] [CrossRef]
- Heilmann, J.; Wasescha, M.R.; Schmidt, T.J. The influence of glutathione and cysteine levels on the cytotoxicity of helenanolide type sesquiterpene lactones against KB cells. Bioorg. Med. Chem. 2001, 9, 2189–2194. [Google Scholar] [CrossRef]
- Mandal, P.K.; Shukla, D.; Govind, V.; Boulard, Y.; Ersland, L. Glutathione conformations and its implications for in vivo magnetic resonance spectroscopy. J. Alzheimers Dis. 2017, 59, 537–541. [Google Scholar] [CrossRef]
- Crestoni, M.E.; Chiavarino, B.; Scuderi, D.; Di Marzio, A.; Fornarini, S. Discrimination of 4-hydroxyproline diastereomers by vibrational spectroscopy of the gaseous protonated species. J. Phys. Chem. B 2012, 116, 8771–8779. [Google Scholar] [CrossRef]
- Zhuang, S.L.; Zhang, J.; Zhang, F.; Zhang, Z.S.; Wen, Y.Z.; Liu, W.P. Investigation of the diastereomerism of dihydrobenzoxathiin SERMS for ER alpha by molecular modeling. Bioorg. Med. Chem. Lett. 2011, 21, 7298–7305. [Google Scholar] [CrossRef]
- Yadav, A.; Mishra, P.C. Modeling the activity of glutathione as a hydroxyl radical scavenger considering its neutral non-zwitterionic form. J. Mol. Model. 2013, 19, 767–777. [Google Scholar] [CrossRef]
- Mak, T.W.; Grusdat, M.; Duncan, G.S.; Dostert, C.; Nonnenmacher, Y.; Cox, M.; Binsfeld, C.; Hao, Z.Y.; Brustle, A.; Itsumi, M.; et al. Glutathione primes T cell metabolism for inflammation. Immunity 2017, 46, 1089–1090. [Google Scholar] [CrossRef]
- Rungeler, P.; Castro, V.; Mora, G.; Goren, N.; Vichnewski, W.; Pahl, H.L.; Merfort, I.; Schmidt, T.J. Inhibition of transcription factor NF-κB by sesquiterpene lactones: A proposed molecular mechanism of action. Bioorg. Med. Chem. 1999, 7, 2343–2352. [Google Scholar] [CrossRef]
- Liu, L.; Hua, Y.P.; Wang, D.; Shan, L.; Zhang, Y.; Zhu, J.S.; Jin, H.Z.; Li, H.L.; Hu, Z.L.; Zhang, W.D. A sesquiterpene lactone from a medicinal herb inhibits proinflammatory activity of TNF-α by inhibiting ubiquitin-conjugating enzyme UBCH5. Chem. Biol. 2014, 21, 1415. [Google Scholar] [CrossRef]
- Garcia-Pineres, A.J.; Lindenmeyer, M.T.; Merfort, I. Role of cysteine residues of p65/ NF-κB on the inhibition by the sesquiterpene lactone parthenolide and N-ethyl maleimide, and on its transactivating potential. Life Sci. 2004, 75, 841–856. [Google Scholar] [CrossRef]
- Lovatt, M.; Filby, A.; Parravicini, V.; Werlen, G.; Palmer, E.; Zamoyska, R. Lck regulates the threshold of activation in primary T cells, while both Lck and Fyn contribute to the magnitude of the extracellular signal-related kinase response. Mol. Cell. Biol. 2006, 26, 8655–8665. [Google Scholar] [CrossRef]
- Visperas, P.R.; Winger, J.A.; Horton, T.M.; Shah, N.H.; Aum, D.J.; Tao, A.; Barros, T.; Yan, Q.R.; Wilson, C.G.; Arkin, M.R.; et al. Modification by covalent reaction or oxidation of cysteine residues in the tandem-SH2 domains of ZAP-70 and Syk can block phosphopeptide binding. Biochem. J. 2015, 465, 149–161. [Google Scholar] [CrossRef]
- Siriwan, D.; Naruse, T.; Tamura, H. Effect of epoxides and alpha-methylene-gamma-lactone skeleton of sesquiterpenes from yacon (Smallanthus sonchifolius) leaves on caspase-dependent apoptosis and NF-κB inhibition in human cercival cancer cells. Fitoterapia 2011, 82, 1093–1101. [Google Scholar] [CrossRef]
- Muller, S.; Murillo, R.; Castro, V.; Brecht, V.; Merfort, I. Sesquiterpene lactones from Montanoa hibiscifolia that inhibit the transcription factor NF-κB. J. Nat. Prod. 2004, 67, 622–630. [Google Scholar] [CrossRef]
- Ploger, M.; Sendker, J.; Langer, K.; Schmidt, T.J. Covalent modification of human serum albumin by the natural sesquiterpene lactone parthenolide. Molecules 2015, 20, 6211–6223. [Google Scholar] [CrossRef]
- Fu, G.; Chen, Y.; Yu, M.; Podd, A.; Schuman, J.; He, Y.; Di, L.; Yassai, M.; Haribhai, D.; North, P.E.; et al. Phospholipase Cγ1 is essential for T cell development, activation, and tolerance. J. Exp. Med. 2010, 207, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Sloan-Lancaster, J.; Kitchen, J.; Trible, R.P.; Samelson, L.E. Lat: The ZAP-70 tyrosine kinase substrate that links t cell receptor to cellular activation. Cell 1998, 92, 83–92. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.P.; Liu, X.; Tang, J.J.; Xiang, P.; Ma, X.M. Semisynthesis, an anti-inflammatory effect of derivatives of 1 β-hydroxy alantolactone from Inula britannica. Molecules 2017, 22, 1835. [Google Scholar] [CrossRef]
- Janganati, V.; Ponder, J.; Balasubramaniam, M.; Bhat-Nakshatri, P.; Bar, E.E.; Nakshatri, H.; Jordan, C.T.; Crooks, P.A. Mmb triazole analogs are potent NF-κB inhibitors and anti-cancer agents against both hematological and solid tumor cells. Eur. J. Med. Chem. 2018, 157, 562–581. [Google Scholar] [CrossRef]
- Akyev, B.; Kasymov, S.Z.; Sidyakin, G.P. Artesin—New sesquiterpenic lactone from Artemisia santolina. Khim. Prir. Soedin. 1972, 6, 733–735. [Google Scholar]
- Schepetkin, I.A.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. High-throughput screening for small-molecule activators of neutrophils: Identification of novel N-formyl peptide receptor agonists. Mol. Pharmacol. 2007, 71, 1061–1074. [Google Scholar] [CrossRef]
- Harling, J.D.; Deakin, A.M.; Campos, S.; Grimley, R.; Chaudry, L.; Nye, C.; Polyakova, O.; Bessant, C.M.; Barton, N.; Somers, D.; et al. Discovery of novel irreversible inhibitors of interleukin (IL)-2-inducible tyrosine kinase (ITK) by targeting cysteine 442 in the ATP pocket. J. Biol. Chem. 2013, 288, 28195–28206. [Google Scholar] [CrossRef]
- Glutathione. PubChem Identifier CID: 124886. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/glutathione (accessed on 15 January 2019).
- Stewart, J.J.P. Optimization of parameters for semiempirical methods v: Modification of nddo approximations and application to 70 elements. J. Mol. Model. 2007, 13, 1173–1213. [Google Scholar] [CrossRef]
- Raghavachari, K. Perspective on “Density functional thermochemistry. III. The role of exact exchange”. Theor. Chem. Acc. 2000, 103, 361–363. [Google Scholar] [CrossRef]
- Stephens, P.J.; Devlin, F.J.; Chabalowski, C.F.; Frisch, M.J. Ab-initio calculation of vibrational absorption and circular-dichroism spectra using density-functional force-fields. J. Phys. Chem. 1994, 98, 11623–11627. [Google Scholar] [CrossRef]
- Peverati, R.; Truhlar, D.G. Improving the accuracy of hybrid meta-GGA density functionals by range separation. J. Phys. Chem. Lett. 2011, 2, 2810–2817. [Google Scholar] [CrossRef]
Sample Availability: Samples of selected compounds are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Compound Number) | Jurkat T Cells | FPR2-HL60 | PMN | Jurkat T Cells | Cyto-Toxicity | |
|---|---|---|---|---|---|---|
| Ca2+ Flux | pERK1/2 | Ca2+ flux | [GSH]i | |||
| IC50 (μM) | ||||||
| Arglabin (1) * | 11.1 ± 2.7 | 28.7 ± 6.3 | N.A. | N.A. | 18.0 ± 1.8 | N.T. |
| α-Epoxyarglabin (1a) | 4.4 ± 1.8 | 32.1 ± 11.5 | N.A. | N.A. | (30%) | N.T. |
| β-Epoxyarglabin (1b) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Pyridinyl arglabin (1c) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Cytisinyl epoxyarglabin (1d) | 3.9 ± 1.2 | 15.4 ± 3.3 | N.A. | N.A. | 12.4 ± 3.5 | N.T. |
| Anabasinyl epoxyarglabin (1e) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Anabasinyl arglabin (1f) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Argolide (2) * | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| 1β,10α-Epoxyargolide (2a) | 14.0 ± 3.4 | 39.4 ± 8.5 | N.A. | N.A. | 36.1 ± 9.4 | N.T. |
| Cytisinyl argolide (2b) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Dihydroargolide (2c) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Grosheimin (3) * | 15.4 ± 4.3 | 43.0 ± 7.5 | N.A. | N.A. | 16.6 ± 4.8 | N.T. |
| Chloroacetate grosheimin (3a) | 6.0 ± 2.1 | 7.8 ± 2.9 | 13.2 ± 3.4 | 10.1 ± 2.8 | 3.8 ± 1.4 | 12.3 ± 3.6 |
| Cytisinyl grosheimin (3b) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| 3β-Hydroxyarhaline (4) * | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Epoxyarhaline (4a) | N.A. | N.A. | N.A. | N.A. | N.A. | N.T. |
| Compound | Name | d(C…S), Å | Docking Score (Cys39 Site) | Activity (Inhibition of TCR Response) | |
|---|---|---|---|---|---|
| Cys39 | Cys78 | ||||
| 1 | Arglabin | 3.55 | 6.74 | −67.6 | Active a |
| 1a | α-Epoxyarglabin | 3.77 | 5.36 | −72.5 | Active |
| 1b | β-Epoxyarglabin | 7.35 | 5.18 | −78.6 | N.A. |
| 1c | Pyridinyl arglabin | 6.15 | 6.85 | −60.9 | N.A. |
| 1d | Cytisinyl epoxyarglabin | No ML | No ML | −104.1 | Active |
| 1e | Anabasinyl epoxyarglabin | No ML | No ML | −85.0 | N.A. |
| 1f | Anabasinyl arglabin | No ML | No ML | −82.4 | N.A. |
| 2 | Argolide | 6.46 | 3.69 | −77.8 | N.A. a |
| 2a | 1β,10α-epoxyargolide | 4.97 | 10.02 | −70.5 | Active |
| 2b | Cytisinyl argolide | No ML | No ML | −85.2 | N.A. |
| 2c | Dihydroargolide | No ML | No ML | −70.7 | N.A. |
| 3 | Grosheimin | 5.25 | 7.45 | −68.8 | Active a |
| 3b | Cytisinyl grosheimin | No ML | No ML | −73.8 | N.A. |
| 4 | 3β-Hydroxyarhaline | No ML | No ML | −66.5 | N.A. a |
| 4a | Epoxyarhaline | No ML | No ML | −80.9 | N.A. |
| Argracin | 4.17 | 6.28 | −98.7 | Active a | |
| Estafiatine | 4.12 | 4.49 | −65.4 | Active a | |
| Parthenolide | 4.70 | 4.30 | −86.4 | Active a | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khlebnikov, A.I.; Schepetkin, I.A.; Kishkentaeva, A.S.; Shaimerdenova, Z.R.; Atazhanova, G.A.; Adekenov, S.M.; Kirpotina, L.N.; Quinn, M.T. Inhibition of T Cell Receptor Activation by Semi-Synthetic Sesquiterpene Lactone Derivatives and Molecular Modeling of Their Interaction with Glutathione and Tyrosine Kinase ZAP-70. Molecules 2019, 24, 350. https://doi.org/10.3390/molecules24020350
Khlebnikov AI, Schepetkin IA, Kishkentaeva AS, Shaimerdenova ZR, Atazhanova GA, Adekenov SM, Kirpotina LN, Quinn MT. Inhibition of T Cell Receptor Activation by Semi-Synthetic Sesquiterpene Lactone Derivatives and Molecular Modeling of Their Interaction with Glutathione and Tyrosine Kinase ZAP-70. Molecules. 2019; 24(2):350. https://doi.org/10.3390/molecules24020350
Chicago/Turabian StyleKhlebnikov, Andrei I., Igor A. Schepetkin, Anarkul S. Kishkentaeva, Zhanar R. Shaimerdenova, Gayane A. Atazhanova, Sergazy M. Adekenov, Liliya N. Kirpotina, and Mark T. Quinn. 2019. "Inhibition of T Cell Receptor Activation by Semi-Synthetic Sesquiterpene Lactone Derivatives and Molecular Modeling of Their Interaction with Glutathione and Tyrosine Kinase ZAP-70" Molecules 24, no. 2: 350. https://doi.org/10.3390/molecules24020350
APA StyleKhlebnikov, A. I., Schepetkin, I. A., Kishkentaeva, A. S., Shaimerdenova, Z. R., Atazhanova, G. A., Adekenov, S. M., Kirpotina, L. N., & Quinn, M. T. (2019). Inhibition of T Cell Receptor Activation by Semi-Synthetic Sesquiterpene Lactone Derivatives and Molecular Modeling of Their Interaction with Glutathione and Tyrosine Kinase ZAP-70. Molecules, 24(2), 350. https://doi.org/10.3390/molecules24020350